
猫冠状病毒非结构蛋白3的多克隆抗体制备及亚细胞定位
2023-11-14 12:11:50王子仪金子安卢辰赫乔治王圣文颜焰周继勇郑肖娟
浙江大学学报(农业与生命科学版) 2023年5期
王子仪,金子安,2,卢辰赫,乔治,王圣文,颜焰,周继勇,郑肖娟*
(1.浙江大学动物医学中心,农业农村部动物病毒学重点实验室,浙江 杭州 310058;2.浙江大学海南研究院,海南 三亚 572025;3.浙江大学-帕特诺尔联合研发中心,浙江 杭州 310058)
冠状病毒可以感染各种哺乳动物和鸟类,临床症状大多与呼吸道、肠道和全身感染有关。猫冠状病毒(feline coronavirus, FCoV)主要感染家猫和野生猫科动物,临床上表现为2 种不同的生物型,一种生物型是通过侵害巨噬细胞引起严重的全身系统性感染的猫传染性腹膜炎(feline infectious peritonitis, FIP)[1],另一种生物型是通过侵害肠道上皮细胞引起轻微肠炎等症状。FCoV与猪传染性胃肠炎病毒(transmissible gastroenteritis virus, TGEV)、猪呼吸道冠状病毒(porcine respiratory coronavirus,PRCV)、犬冠状病毒(canine coronavirus, CCoV)和猪流行性腹泻病毒(porcine epidemic diarrhea virus,PEDV)同为冠状病毒科(Coronaviridae)α冠状病毒属(Alphacoronavirus)的成员[2],其病毒颗粒整体呈球形,囊膜表面有不规则的花瓣状凸起,在电子显微镜下呈皇冠状,直径为90~100 nm[3]。FCoV是不分节段的单股正链RNA 病毒,基因组全长约为29 kb,共有11 个可读框(open reading frame,ORF)。与冠状病毒家族的其他成员类似,FCoV基因组的整体结构依次为5´-leader-UTR-pp1abspike-Nsp3a-Nsp3b-Nsp3c-envelope-membranenucleocapsid-Nsp7a-Nsp7b-3´UTR-Poly(A)[4];FCoV中2 个部分重叠的ORF(ORF1a 和ORF1b)占据了基因组5´端的大部分区域(约20 kb),借助核糖体移码翻译产生的多聚蛋白前体(pp1ab)在病毒编码的木瓜样蛋白酶(papain-like protease, PLP)和3C样蛋白酶(3-chymotrypsin-like protease, 3CLP)的作用下水解生成16 个成熟的非结构蛋白[(nonstructural protein 1, Nsp1)~Nsp16],参与蛋白质降解、基因组复制和mRNA的合成过程[5-6]。
正链RNA 病毒的一个共同特征是它们可以组装与细胞质膜相关的复制/转录复合体(replication and transcription complex, RTC)。冠状病毒能维持庞大的基因组与其组装复制/转录复合体的特征有关[7],冠状病毒会将感染的宿主细胞膜重排成细胞器样复制结构,该结构由双膜囊泡(doublemembrane vesicle, DMV)和卷曲膜组成,参与RNA合成的非结构蛋白就定位于这些膜上,例如Nsp7和Nsp8 是RNA 聚合酶的辅助因子,Nsp9 是结合RNA 的磷酸酶,Nsp12 是RNA 依赖的RNA 聚合酶,Nsp13 是RNA 解旋酶,Nsp14 是拥有校对功能的3´→5´核糖核酸外切酶[8]。冠状病毒的非结构蛋白除了参与病毒的转录复制外,在病毒和宿主细胞的互作中也发挥重要作用,是重要的功能蛋白[8]。
Nsp3 是冠状病毒基因组中分子量最大(约170 kDa)的非结构蛋白,在冠状病毒的不同属中都具有保守的8 个结构域和2 个跨膜区[9]。这些结构域包括泛素样结构域1(ubiquitin-like domain 1,Ubl1)、酸性残基(acidic residues, Ac)结构域、二磷酸腺苷核糖磷酸酶(adenosine diphosphate-ribose phosphatase, ADRP)结构域、泛素样结构域2(ubiquitin-like domain 2, Ubl2)、PLP结构域、核酸结合(nucleic acid-binding, NAB)结构域、胞外域(ectodomain)以及未知功能的Y 结构域,在病毒感染中发挥着不同的作用[10]。Nsp3从pp1ab中被释放后,先与其他非结构蛋白相互作用后,再与病毒RNA 结合形成RTC[11]。Nsp3 在冠状病毒生命周期内可以作为支架蛋白与其他病毒蛋白和宿主蛋白结合,Nsp3 的Ubl1 和Ac 结构域与病毒的核衣壳(nucleocapsid, N)蛋白之间存在互作,Nsp3 还会通过胞外域的氮端糖基化与宿主蛋白(如RCHY1)互作以促进病毒的存活[12]。Nsp3 可能与宿主的免疫反应相关,其与Nsp4 和Nsp6 共表达时可以在严重急性呼吸综合征冠状病毒(severe acute respiratory syndrome coronavirus, SARS-CoV)感染的细胞中诱导双膜囊泡的形成[13]。然而,关于FCoV Nsp3 的许多功能有待进一步研究。感染FCoV的猫会对非结构蛋白产生早期应答反应[9,14],但是Nsp3 蛋白氮端的免疫原性较低,这可能是因为Ac结构域中存在高丰度的甘氨酸[15-16]。
Nsp3 作为切割多聚蛋白前体的酶在病毒感染中发挥关键作用,被认为是广谱抗冠状病毒药物开发的潜在靶点[17]。目前针对FCoV非结构蛋白的研究进展缓慢,只能借助其他研究较充分的冠状病毒推测其功能,此外,FCoV Nsp3抗体工具的缺乏限制了Nsp3的蛋白质功能研究,给研究FCoV Nsp3与其他病毒蛋白或宿主蛋白之间的互作关系带来了困难[6]。因此,本研究在表达和纯化Nsp3 重组蛋白的基础上制备了FCoV Nsp3 的特异性多克隆抗血清(多抗血清),并通过激光共聚焦显微镜观察了该蛋白在感染细胞中的亚细胞定位,以期为后续开展Nsp3的生物学功能研究奠定基础。
1 材料与方法
1.1 毒株、细胞与实验动物
FCoV WSU 79-1683毒株和猫肾细胞(Crandell-Rees feline kidney, CRFK)获赠于复旦大学基础医学院叶荣老师。载体质粒pCMV-N-Flag、pEGFPC3、pCOLD-TF 由本实验室保存;膜结构定位质粒pEYFP-Golgi(高尔基体)、pEGFP-Mem(细胞膜)、pDsRed-Mito(线粒体)和pDsRed-ER(内质网),以及微丝骨架定位质粒pEGFP-Act 均购自美国Clontech 公司;大肠埃希菌感受态细胞DH5α和BL21(DE3)由本实验室制备并保存;N 蛋白兔多抗血清由本实验室制备并保存;6~8周龄SPF级雌性BALB/c小白鼠购自浙江中医药大学动物实验研究中心。本研究动物试验参照《实验动物 福利伦理审查指南》(GB/T 35892—2018)进行。
1.2 实验试剂与材料
TRIzol 试剂盒购于上海普飞生物技术有限公司;反转录试剂盒、DNA 分子量分子标志物购于美国Thermo Fisher Scientific 公司;KOD One 高保真DNA 聚合酶购于杭州硕盟生物科技有限公司;超级感受态细胞制备试剂盒购于生工生物工程(上海)股份有限公司;聚合酶链反应(polymerase chain reaction, PCR)清洁回收试剂盒、凝胶回收试剂盒购自南京诺唯赞生物科技股份有限公司;质粒抽提试剂盒购自天根生化科技(北京)有限公司;限制性内切酶购自日本TaKaRa 公司;引物合成由生工生物工程(上海)股份有限公司完成;菌液测序由浙江尚亚生物技术有限公司完成;Flag/绿色荧光蛋白(green fluorescence protein, GFP)标签抗体购自杭州华安生物技术有限公司;辣根过氧化物酶(horseradish peroxidase, HRP)、异硫氰酸荧光素(fluorescein isothiocyanate, FITC)、A546标记的山羊抗鼠/兔IgG均购自美国Sigma公司;胰酶和Dulbecco’s modified Eagle’s medium(DMEM)细胞培养基购自美国Gibco公司;胎牛血清购自上海吉泰依科赛生物科技有限公司;异丙基硫代-β-D-半乳糖苷(isopropylthioβ-D-galactoside, IPTG)、Ni-TED 镍亲和纯化试剂盒购于德国Qiagen公司;Quick Antibody-Mouse5W佐剂购自苏州博特龙免疫技术有限公司;jetPRIME转染试剂购自法国Polyplus公司。
1.3 Nsp3 蛋白的生物信息学分析
从NCBI网站(https://www.ncbi.nlm.nih.gov/)搜索并下载不同物种的冠状病毒代表毒株序列,共51条(统一命名为GenBank 登录号/病毒缩写/毒株名);现有犬猫冠状病毒常见毒株序列,共83 条(统一命名为GenBank 登录号/分离时间/国家名/毒株名/宿主名)。经Mafft 软件对齐后,用MEGA7 软件基于临接法构建进化树[利用自展法(bootstrap)检验1 000 次]。使用iTOL 软件美化进化树,使用Jalview软件进行序列比对。
1.4 重组原核表达质粒的构建与鉴定
利用DNAstar 软件对WSU 79-1683 毒株(GenBank登录号:JN634064.1)的Nsp3 序列进行抗原表位和跨膜区预测,随后截取第50—550 位氨基酸并利用Primer Premier 5.0 软件设计引物,上游引物P1为5´-CGGGATCCACGACTTGGGATGAATT TGAAG-3´(下划线处为BamHⅠ酶切位点),下游引物P2为5´-GCGTCGACTTAGACTTTGAAAAT ACCAACACTAATAAG-3´(下划线处为SalⅠ酶切位点)。采用TRIzol法提取病毒基因组RNA并进行反转录,以合成的cDNA 为模板,用KOD One 高保真DNA 聚合酶对截短型Nsp3基因进行扩增,PCR程序中退火温度设置为58 ℃,延伸时间设置为30 s,循环35 次。对清洁回收的PCR 产物进行BamHⅠ和SalⅠ双酶切,同时对pCOLD-TF载体进行双酶切后割胶回收,于16 ℃条件下连接过夜。将连接产物转化DH5α感受态细胞并涂布于含氨苄抗性的Luria-Bertani(LB)培养基平板,培养14~20 h后,挑取单克隆接种到含氨苄抗性的LB 液体培养基中,于37 ℃下振荡培养2 h 后,对菌液进行PCR扩增。将PCR 鉴定结果呈阳性的菌液送至浙江尚亚生物技术有限公司进行测序,将上述过程中构建成功的重组质粒命名为pCOLD-FCOV-Nsp3。
1.5 重组蛋白的表达与纯化
将pCOLD-FCOV-Nsp3质粒转化BL21感受态细胞并涂布于含氨苄抗体的LB 培养基平板,培养14 h 后挑取单克隆,接种到2 mL 含氨苄抗性的LB液体培养基中,于37 ℃条件下振荡培养12~16 h进行活化,将菌液按1∶100 的比例扩繁培养至吸光度值D(600 nm)=0.5 时,吸取1 mL 菌液置于干净的Eppendorf(EP)管中,作为诱导前蛋白样品。将剩余菌液即刻转移至4 ℃条件下降温30 min,再加入IPTG使其终浓度为1 mmol/L。在低温(16 ℃)条件下诱导过夜以表达目的蛋白。取1 mL 诱导后的菌液并离心,收集菌体沉淀,经细胞裂解液处理后制备成蛋白样品,进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis, SDS-PAGE)和考马斯亮蓝染色(Coomassie blue staining, CBS)。采用蛋白质印迹法(Western blotting, WB)对表达的蛋白进行鉴定,以稀释1 000 倍的His 鼠源抗体作为一抗,以稀释4 000 倍的HRP 标记的山羊抗鼠IgG 作为二抗进行孵育,当WB分析和条带均符合预期时,确认目标重组蛋白进行了表达。采用镍柱亲和层析法对目的蛋白进行纯化,方法如下:收集剩余菌体沉淀,用磷酸盐缓冲液(phosphate buffered saline, PBS)清洗3次并重悬,加入溶菌酶后低温处理30 min,超声破碎,在12 000 r/min 下离心20 min,收集上清液并用0.45 μm 滤膜过滤,然后与2 mL Ni-TED 镍填料于4 ℃条件下充分结合过夜,最后将结合后的混悬液转移至蛋白纯化柱,依次用不同浓度的咪唑缓冲液洗脱并收集纯化后的重组蛋白。
1.6 多克隆抗体的制备及反应性鉴定
将成功表达和纯化的Nsp3 重组蛋白与Quick Antibody-Mouse5W 佐剂等比例混合后,采用小腿肌肉注射方式免疫6~8周龄的BALB/c雌鼠,每只小鼠注射100 μL(所需Nsp 重组蛋白约为25 μg),初次免疫后21 d 时按同样的方法注射小鼠以加强免疫。初次免疫后35 d 于小鼠尾部采血,于37 ℃条件下静置30 min,再转移至4 ℃条件下析出血清,以1 000g离心10 min 后小心吸取位于上层的血清,用间接酶联免疫吸附测定(enzyme-linked immunosorbent assay, ELISA)检测抗体效价后冻存,备用。
利用WSU 79-1683 毒株感染CRFK 细胞24 h后收集细胞,用WB 法鉴定Nsp3 多抗血清对感染细胞样品的反应性,以稀释1 000倍的Nsp3 多抗血清作为一抗,以稀释4 000 倍的HRP 标记的山羊抗鼠IgG 作为二抗进行孵育。将CRFK 细胞铺至96孔板中,待细胞生长至密度为80%时,以病毒感染复数(multiplicity of infection, MOI)为0.1 接种WSU 79-1683 病毒,待观察到合胞体等细胞病变(需要约24 h)时,用4%甲醛固定液于室温下固定15 min,加入含0.1% Triton X-100 的PBS 溶液透膜处理10 min,加入5%脱脂奶于37 ℃条件下封闭1 h。加入稀释1 000 倍的Nsp3 多抗血清作为一抗,在37 ℃摇床上孵育1 h,用含0.05% Tween-20的PBS(PBS with Tween 20, PBST)清洗5 遍;加入稀释400 倍的FITC 标记的山羊抗鼠IgG 作为二抗,于37 ℃条件下避光孵育1 h,用PBST 清洗5遍;用4´,6-二脒基-2-苯基吲哚(4´, 6-diamidino-2-phenylindole, DAPI)对细胞核染色后,每孔加入50 μL PBST,于荧光显微镜下避光观察并拍照记录。
1.7 FCoV感染细胞中Nsp3蛋白的亚细胞定位分析
将CRFK 细胞铺至共聚焦皿中,待细胞生长至密度为60%~70%时,用转染试剂jetPRIME对膜结构定位质粒pEYFP-Golgi、pEGFP-Mem、pDsRed-Mito、pDsRed-ER和微丝骨架定位质粒pEGFP-Act进行转染。在200 μL jetPRIME 转染缓冲液中加入2 μg 质粒,混匀后再加入4 μL jetPRIME 转染试剂,室温下静置15 min,再将质粒和转染试剂混合物轻轻吹打混匀后加到共聚焦皿中,将上述培养皿放回37 ℃细胞培养箱中继续培养4 h 后,用含2%血清的培养基换液。完成转染12 h 后,以MOI 为0.1 接种WSU 79-1683 病毒,感染16 h 后固定细胞,基于免疫荧光双标法同时对Nsp3 和N 蛋白进行标记。以稀释1 000 倍的Nsp3 蛋白鼠多抗血清和N 蛋白兔多抗血清作为一抗,以稀释400 倍的FITC/A546标记羊抗兔(N 蛋白)和A647 标记羊抗鼠(Nsp3 蛋白)作为二抗同时孵育,用DAPI 对细胞核染色,置于Zeiss LSM-880 激光共聚焦显微镜下观察并拍照,记录感染细胞中结构蛋白N、非结构蛋白Nsp3与亚细胞结构的定位关系和结构变化情况。
2 结果与分析
2.1 冠状病毒Nsp3 蛋白的遗传进化分析
从GenBank数据库下载不同物种冠状病毒代表毒株的51条Nsp3蛋白序列进行进化树分析,结果显示:FCoV与CCoV、TGEV、PRCV的亲缘关系较近,而与同属其他冠状病毒的亲缘关系较远(附图1A,http://www.zjujournals.com/agr/CN/10.3785/j.issn.1008-9209.2022.08.011),这印证了JAIMES 等[18]基于S基因提出的α冠状病毒属进化模式,即Ⅰ型FCoV 与Ⅰ型CCoV 拥有共同祖先,并通过重组逐渐演化出Ⅱ型FCoV、Ⅱ型CCoV、TGEV 和PRCV。进一步从GenBank 数据库下载83 条犬猫冠状病毒Nsp3蛋白序列进行进化树分析,结果显示:进化树主要分为3 个分支,除FCoV 与CCoV 分支外,从英国分离到的FCoV 毒株与其他毒株亲缘关系较远;以不同颜色标记毒株的分离地点(国家),发现进化树分支的聚集现象与毒株分离地点(国家)存在一定程度的相关性(附图1B,http://www.zjujournals.com/agr/CN/10.3785/j.issn.1008-9209.2022.08.011)。在此基础上,将其中56 条FCoV Nsp3 蛋白的核苷酸序列进行比对,发现多个不保守区域,例如400~580、1 300~1 354、2 745~2 796 bp 区域,且序列相似度(统一着色)从95%降到5%条件下依然存在的空白区域为少数毒株独有的插入突变(图1A)。同时,以WSU 79-1146为参照截取Nsp蛋白的核苷酸序列比对结果发现,多数毒株蛋白的核苷酸序列在400~580 bp区域存在不同程度的插入或缺失突变(图1B)。

图1 Nsp3蛋白的序列比对分析Fig.1 Sequence alignment analysis of Nsp3 protein
2.2 Nsp3 蛋白的跨膜区、信号肽和抗原表位预测
采用TMHMM 2.0 软件对Nsp3 蛋白进行跨膜区预测,结果显示:WSU 79-1683 毒株的Nsp3 蛋白存在4 个跨膜结构域,分别位于第944—962、1 005—1 027、1 085—1 107、1 127—1 149 位氨基酸(图2A)。采用SignalP 5.0 软件对Nsp3 蛋白进行信号肽预测,结果显示WSU 79-1683 毒株的Nsp3 蛋白无信号肽序列(图2B)。进一步使用DNAstar 软件预测并分析其抗原表位和亲和力,结果如图2C 所示。由于Nsp3 蛋白翻译产物分子量较大且存在4 个跨膜结构域,因此挑选抗原性高的截短区域(第50—550 位氨基酸)用于后续的原核表达。

图2 WSU 79-1683 Nsp3蛋白的跨膜区、信号肽和抗原表位预测Fig.2 Transmembrane region, signal peptide, and epitope prediction of WSU 79-1683 Nsp3 protein
2.3 Nsp3 蛋白的原核表达与纯化
以FCoV(WSU 79-1683 毒株)的cDNA 为模板,采用PCR 成功扩增Nsp3 截短片段(第50—550位氨基酸)。为获得可溶性的His-Nsp3 重组蛋白,将Nsp3 截短片段(第50—550 位氨基酸)亚克隆至pCOLD-TF 表达载体上,成功构建pCOLD-TFNsp3 重组原核表达质粒。进一步将pCOLD-TFNsp3 质粒转化BL21(DE3)感受态细胞后,于37 ℃条件下振荡培养过夜,隔日按照1∶200 的比例扩繁培养至菌液吸光度值D(600 nm)约为0.5 时,冰水浴30 min 后加入IPTG 至终浓度为1 mmol/L。在16 ℃条件下振荡诱导16~20 h,收集诱导前后的菌体蛋白进行SDS-PAGE 分析和考马斯亮蓝染色。结果显示,诱导后His-Nsp3 重组蛋白大量表达,已知载体标签触发因子大小约为50 kDa,截短的Nsp3约为56 kDa,预测获得目的蛋白的条带大小约为106 kDa,诱导后的重组蛋白在100~130 kDa 范围内大量表达,其大小与预期结果相符。利用镍柱亲和层析在非变性条件下对His-Nsp3 重组蛋白进行纯化,用含有40、50、60 mmol/L 咪唑的洗脱溶液洗脱后获得了纯化后浓度较高的目的蛋白,其最高质量浓度为0.48 mg/mL(图3)。采用His 标签抗体对纯化蛋白进行WB 检测,结果显示纯化蛋白在130 kDa 附近呈单一条带,小心裁去130 kDa 处曝光条带后,其对应下方其他染色条带的位置也出现微弱条带,推测为Nsp3 的降解产物(图3)。上述结果说明成功实现了对FCoV Nsp3 的原核表达与纯化。

图3 Nsp3重组蛋白的诱导表达、亲和纯化与鉴定Fig.3 Induced expression, affinity purification, and identification of Nsp3 recombinant protein
2.4 Nsp3 多抗血清与FCoV 感染细胞的反应性鉴定
收集FCoV 感染和未感染的CRFK 细胞样品,采用Nsp3 多抗血清进行WB 检测(内参使用β-actin),结果显示,稀释1 000倍的Nsp3多抗血清可与FCoV 感染的CRFK 细胞样品发生特异性反应,在100 kDa 以上位置出现了拖尾条带,而未感染的CRFK 细胞样品没有出现类似条带(图4A)。免疫荧光实验显示,Nsp3多抗血清可与FCoV感染的CRFK 细胞发生特异性的免疫荧光反应,且呈现特征性的颗粒状或团块状信号,而在未感染的CRFK 细胞中则没有观察到类似的荧光信号(图4B)。这些结果表明,制备的Nsp3 多抗血清与FCoV 感染的CRFK 细胞之间具有特异的WB 和间接免疫荧光试验(indirect immunofluorescence assay,IFA)反应性。
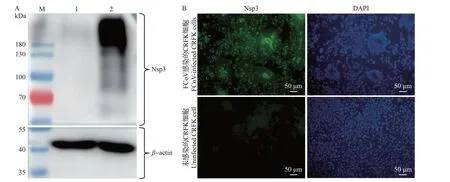
图4 Nsp3多抗血清的蛋白质印迹法(A)和间接免疫荧光试验(B)反应性结果Fig.4 Reactivity results of Nsp3 polyclonal antiserum by WB (A) and IFA (B)
2.5 Nsp3 蛋白在FCoV 感染的宿主细胞内的定位观察
跨膜区预测显示Nsp3 蛋白具有4 个跨膜结构域。为进一步分析Nsp3 蛋白与细胞内多种膜结构或其他细胞器间的定位关系,将膜结构定位质粒pEYFP-Golgi、pEGFP-Mem、pDsRed-Mito 和pDsRed-ER以及微丝骨架定位质粒pEGFP-Act分别转染至CRFK 细胞,感染FCoV 后固定细胞,并分别用N 蛋白兔多抗血清、Nsp3 蛋白鼠多抗血清对其进行免疫荧光双标染色,通过激光共聚焦显微镜对FCoV 感染细胞中病毒蛋白的亚细胞定位情况进行观察。结果如图5 所示:在感染细胞中,无论是否形成合胞体,FCoV 的N 蛋白都均匀分布在细胞质内;而Nsp3 蛋白在感染细胞的胞浆内或合胞体的中央区域聚集成颗粒状或团块状,与线粒体、高尔基体、微丝和细胞膜均没有明显的信号重叠,但与内质网的信号重叠率高,且经ImageJ 软件计算,其皮尔逊相关系数为0.82,提示Nsp3 与内质网之间存在明显的共定位现象。

图5 Nsp3在FCoV感染CRFK细胞中的亚细胞定位Fig.5 Subcellular localization of Nsp3 in FCoV-infected CRFK cells
3 讨论与结论
冠状病毒是一类广泛存在于自然界的RNA 病毒,其宿主遍布哺乳动物甚至鸟类,严重威胁着畜牧业生产和公共安全[19]。更严重的是,冠状病毒容易发生重组和突变,也能逐渐改变细胞嗜性和毒力[20],不断促使自身适应新的宿主,导致新的冠状病毒毒株甚至新的冠状病毒物种不断出现,给冠状病毒防控工作带来了巨大的挑战[21-22]。虽然目前还没有FCoV感染人的病例出现,但全基因组测序证明,在急性呼吸道疾病患者中,有1.50%~3.54%感染了犬猫冠状病毒的重组毒株(canine-feline-porcinelike, CFPL)CoVs[23]。2020 年,众多猫被饲主传染SARS-CoV-2 的实例已被报道[24],尽管没有大流行病暴发,也应防止宠物猫被人传染SARS-CoV-2而造成病毒重组风险。在宠物猫与人和其他宠物共居的客观环境下,应该定期对猫砂盆进行清洁和消毒处理,以减少冠状病毒潜在的种间重组风险,进而降低未来新的冠状病毒再次暴发的可能性。
20 世纪60 年代前后FCoV 从美国传播到世界各地,如今在某些多猫环境中,FCoV检出率已经超过90%,再加上近年来猫作为伴侣动物的受欢迎程度大大提高,而且城市化进程加快也使得野生或半野生(社区)猫的数量进一步增长,导致多猫聚集环境下FCoV传播风险加大[25-26]。此外,肠道病毒有着通过粪便持续排毒的特点,因此FCoV 通过粪口传播的概率也随着猫的密集程度上升而显著上升[4]。
抗体依赖增强(antibody dependent enhancement, ADE)作用导致疫苗开发遭遇瓶颈,而由于特效药的缺乏,一般采取支持疗法帮助患猫延缓疾病进展[27]。虽然已有应用抗病毒化合物GS-441524成功治疗患猫的案例,但出于安全性和实用性的考量,目前抗病毒化合物GS-441524还未获得上市批准[28]。在宿主的天然免疫反应抑制病毒复制的同时,病毒也会进化出一系列策略以躲避宿主的天然免疫反应。Nsp3 蛋白在冠状病毒的生命周期中发挥关键作用,同时,基于其序列的高度保守性,Nsp3被认为是广谱抗冠状病毒药物开发的潜在靶点[29]。
本研究将截短的Nsp3基因克隆到原核表达载体上,成功表达了可溶性FCoV Nsp3截短蛋白并制备了多抗血清,WB 和IFA 结果显示Nsp3 蛋白对病毒感染细胞具有特异性反应。我们还研究了Nsp3蛋白在FCoV 感染细胞中的分布情况,并观察到Nsp3 与内质网存在明显的共定位现象,推测该行为有利于Nsp3 蛋白发挥多种生物学功能(如切割多聚蛋白前体、组装复制/转录复合体、诱导双膜囊泡形成和调控宿主天然免疫反应等[27,30])。同时,本研究填补了猫冠状病毒非结构蛋白在细胞中定位相关研究的空白,也为深入探究FCoV 的复制机制奠定了基础。
致谢感谢浙江大学农生环测试中心李云琴老师在激光共聚焦实验中提供的技术支持。
猜你喜欢
科学大观园(2022年2期)2022-01-23 11:05:15
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
食品科学(2018年10期)2018-05-23 01:27:28
天津科技大学学报(2016年1期)2016-02-28 16:59:45
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
动物医学进展(2015年10期)2015-12-07 05:46:18
西南医科大学学报(2015年1期)2015-08-22 13:01:46
中国当代医药(2015年9期)2015-03-01 02:01:59
现代检验医学杂志(2015年2期)2015-02-06 02:01:01
西南军医(2015年6期)2015-01-23 01:25:50
