
151 份小麦品种(系)中主要品质基因的分子检测
2023-11-08 06:02钱禄巧金彦刚杨永乐夏中华
江汉大学学报(自然科学版) 2023年5期
张 红,王 歆*,钱禄巧,金彦刚,杨永乐,夏中华
(1.安徽科技学院 农学院,安徽 凤阳 233100;2.江苏瑞华农业科技有限公司/江苏省小麦商业化育种工程技术研究中心,江苏 宿迁 223800)
0 引言
小麦是禾本科(Gramineae)植物中最重要的栽培作物之一[1],也是全世界种植范围最广、面积最大、产量最高、贸易额最多的粮食作物之一,世界上约40%的人以小麦为主要食物[2]。小麦作为我国第三大粮食作物,常年播种面积和产量约占全国粮食作物的五分之一以上[3]。但小麦易受各种病害的侵染从而造成产量和品质下降。近些年来,分子标记辅助育种的方法已用于提高小麦品质和产量[4]。
小麦面粉的食用和加工品质受蛋白质含量或面筋强度、淀粉特性、面粉颜色和硬度等品质特征的影响。高分子量麦谷蛋白亚基(HWM-GS)是小麦面团流变学和面筋特性的主要因素[5-6]。它由位于小麦第一条同源染色体上Glu-A1、Glu-B1和Glu-D1基因座的基因编码,每个基因座包含两个紧密相连的基因[7]。它们编码具有较高分子量的X 型亚基和具有较低分子量的Y 型亚基[8]。虽然HWM-GS 只占小麦贮藏蛋白的10%,但是对面筋强度和面团黏弹性起决定性作用[9-11]。有研究[12]认为,含有Ax1、Ax2、Dx5、By8、Bx7、Dy10基因的小麦面粉制作的面包和面条品质会更好。在加工和储存过程中,很多面制食品会出现褐变现象,不仅会影响食物的外观,还会影响其营养价值[13-14]。多酚氧化酶(Polyphenol oxidase,PPO)是引起面粉制品颜色褐变的主要因素[15-16],其主效基因Ppo-A1和Ppo-D1位于染色体2AL 和2DL 上[17]。Sun 等[18]开发了定位于2AL 的PPO基因标记PPO18,该标记能有效区分控制PPO 活性高低的等位基因Ppo-A1a和Ppo-A1b。He 等[19]开发了定位于2DL 上的互补标记PPO16 和PPO29,能够有效区分Ppo-D1位点的等位基因Ppo-D1a(PPO 活性低)和Ppo-D1b(PPO 活性高)。因此,国内外致力于选育PPO 活性低的小麦品种。1B/1R异位系在世界各地应用十分广泛,在抗病、抗逆和丰产性等方面具有独特的优越性,曾在我国小麦育种和生产上大量利用。但该异位系加工品质普遍较差,面筋强度降低,烘烤品质差。非1B/1R异位系的黄色素含量更低,因此,低PPO 活性的非1B/1R异位系小麦品种,可用于改良小麦品种面粉色泽[20]。
本研究利用分子标记技术,对江苏省宿迁市瑞华农业科技有限公司151 份小麦品种(系)中的等位变异情况进行检测和分布规律分析,从中筛选出具有优质高分子量谷蛋白亚基、低PPO活性和非1B/1R异位系的小麦品种,为小麦的育种提供支持。
1 材料与方法
1.1 试验材料
选用江苏瑞华农业科技有限公司试验田2020 年主要推广的小麦品种和试验品系,每个品种(系)种植2 行,行长4 m,行距50 cm,株距20 cm。采用CTAB 法[21]提取2020 年12 月份田间采集的小麦叶片总DNA,并稀释至150 mg/μL,置于-20 ℃冰箱保存待用。每个品种(系)随机取样3 份。
1.2 小麦DNA 提取及分子标记检测
本文所用检测引物根据文献[22-25]进行选择,由生工生物工程(上海)股份有限公司合成,具体信息见表1。10 μL 2 × Taq Master Mix(北京普鲁顿生物科技有限公司),4SGelred 核酸染剂(生工生物工程(上海)股份有限公司)。PCR 扩增(基因扩增仪:TOUCH 960 型,杭州晶格科学仪器有限公司)反应体系为16 μL:DNA 2 μL,ddH2O 9.9 μL,10 × buffer(含Mg²+)1.5 μL,dNTP(10 mmol/L)1.5 μL,Primer F(10 pmol/μL)0.5 μL,Primer R(10 pmol/uL)0.5 μL,Taq 酶(DNA 聚合酶)0.1 μL。扩增产物使用1.5%琼脂糖凝胶在1 × TAE 缓冲液中进行电泳检测,使用凝胶成像系统(培清JS-680D 全自动凝胶成像分析仪)拍照保存。

表1 小麦基因标记引物及扩增片段大小Tab.1 Primers and amplified fragments of wheat gene markers
2 结果与分析
2.1 151 份小麦材料面筋强度相关基因分子检测
在Glu-A1、Glu-D1和Glu-B1位点共检测出4 种类型蛋白亚基基因(见表1)。在Glu-A1位点为稀有亚基基因Ax2,利用标记Ax2可以扩增出1 319 bp 目的片段,在151 份小麦材料中有2 份材料含有Ax2 基因,频率为1.32%(部分结果见图1、表2);在Glu-D1位点为亚基基因Dx5,利用标记Dx5可以扩增出450 bp 目的片段,有26 份材料含有Dx5基因,频率为17.22%(部分结果见图2、表2);在Glu-B1位点检测出By8和Bx7基因,利用标记By8和Bx7可以扩增出527 bp 和447 bp 的目的片段,含有By8基因有106 份,含有Bx7基因有135 份,频率分别为70.20%和89.40%(部分结果见图3 和图4、表2)。其中,在Glu-B1位点上亚基Bx7基因出现频率最高。

图1 Ax2 标记扩增部分品种琼脂糖凝胶电泳Fig.1 Agarose gel electrophoresis of some varieties amplified by Ax2 markers

图2 Dx5 标记扩增部分品种琼脂糖凝胶电泳Fig.2 Agarose gel electrophoresis of some varieties amplified by Dx5 markers
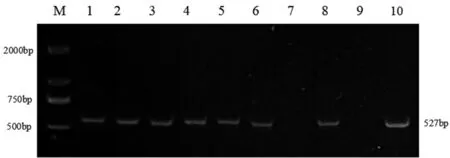
图3 By8 标记扩增部分品种琼脂糖凝胶电泳Fig.3 Agarose gel electrophoresis of some varieties amplified by By8 markers
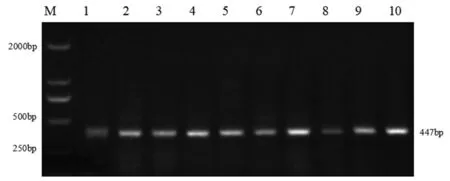
图4 Bx7 标记扩增部分品种琼脂糖凝胶电泳Fig.4 Agarose gel electrophoresis of some varieties amplified by Bx7 markers

表2 151 份小麦材料品质基因型表现Tab.2 Genotypic performance of quality traits of 151 wheat materials
2.2 151 份小麦材料面粉色泽相关基因分子检测
在Ppo-A1基因位点,利用共显性标记引物PPO18 可检测低PPO 活性基因Ppo-A1b和高PPO 活性基因Ppo-A1a,可以扩增出685 bp 和876 bp 的目的片段。在151 份小麦材料中有77 份材料是高PPO 活性的Ppo-A1a基因型,频率为50.99%;有69 份材料是低PPO 活性的Ppo-A1b基因型,频率为45.70%(部分结果见图5、表2);最终还有5 份模糊条带,其结果不明,所占比例为3.31%。在Ppo-2D基因位点,利用互补显性标记引物PPO16 和PPO29,检测出低PPO 活性基因Ppo-D1a和高PPO 活性基因Ppo-D1b,分别扩增出713 bp 和490 bp 的目的片段。有112 份含有Ppo-D1b基因,其频率为74.17%;有39 份材料含有Ppo-D1a基因,其频率为25.83%(部分结果见图6、表2)。同时含有Ppo-A1b和Ppo-D1a基因有13 份材料,频率为8.61%。

图5 PPO18 标记扩增部分品种琼脂糖凝胶电泳Fig.5 Agarose gel electrophoresis of some varieties amplified by PPO18 markers

图6 PPO29 和PPO16 标记扩增部分品种琼脂糖凝胶电泳Fig.6 Agarose gel electrophoresis of some varieties amplified by PPO29 and PPO16 markers
2.3 151 份小麦材料1B/1R 异位系的标记检测
在Glu-B3j基因位点为1B/1R异位系基因,利用1B/1R标记扩增出1 598 bp 的目的片段,在151 份材料中共检测出131 份含有1B/1R异位系,其频率为86.75%;非1B/1R异位系对小麦面粉的白度具有很好的正向作用,共有20 份,其频率为13.25%(部分结果见图7、表2)。

图7 1B/1R 异位系标记扩增部分品种琼脂糖凝胶电泳Fig.7 Agarose gel electrophoresis of some varieties amplified by 1B/1R translocation line markers
2.4 聚合优异品质基因材料的筛选
在151 份小麦材料中共检测8 个品质相关优质基因,优质基因组合共11 个(见表3)。在品质基因中,小麦籽粒中的高分子量麦谷蛋白亚基是决定小麦加工品质、面团流变特性和面团微观结构的重要因素。小麦基因的组合和相对表达显著都会影响面粉的最终用途,在HWM-GS 中优质基因组合是Ax2+Dx5、Ax2+By8、Ax2+Bx7,这3 种组合的频率都是0.66%,含有这3 种优质基因组合的材料是西农235;Dx5+By8、Dx5+Bx7、By8+Bx7优质基因组合的频率分别是7.28%、14.57%、62.25%;Ax2+Dx5+Bx7、Dx5+By8+Bx7优质基因组合的频率分别是0.66%、4.64%。小麦低PPO 活性可降低面粉在加工和贮存过程中的褐变程度,所以双低Ppo-A1b/Ppo-D1a为优质基因组合,共有13 份材料,其频率为8.60%;非1B/1R异位系材料的黄色素含量更低,同时携带低PPO 活性,且为非1B/1R异位系的材料有2 份,为优异等位基因组合,频率为1.32%。

表3 151 份小麦材料品质性状优质基因组合Tab.3 Excellent gene combinations of quality traits of 151 wheat materials
3 讨论
近年来,品质育种在小麦育种研究中越来越受欢迎和关注。小麦的品质性状是由基因和外在环境共同决定的,如基因型、外界环境、土地酸碱度、小麦热敏感性等多种因素影响[26]。本研究利用分子标记对宿迁市江苏瑞华农业科技有限公司试验田种植的151 份小麦品种(系)中8 个品质基因进行检测,研究结果可为今后小麦品质育种亲本选择和品种布局提供依据。
Dhaka 等[27]研究发现,HWM-GS 对改良小麦面筋强度具有非常重要的作用,按照作用程度可分为Glu-D1>Glu-B1>Glu-A1。具有Glu-A1位点上的基因的小麦品种有较好的面包烘烤品质,具有Glu-B1位点上的基因的小麦品种对烘烤品质有正向效应,具有Glu-D1位点上的基因的小麦品种与面包品质呈正相关[13]。本试验研究结果发现,在Glu-A1位点上,优质基因Ax2的频率为1.32%,Ax2亚基是稀有优质亚基,在我国普通小麦品种中分布很少[6];在Glu-B1位点上,Bx7、By8分别出现的频率为89.4%、70.2%,由于基因间存在互作Bx7+By8的形式,其频率为61.58%,蛋白亚基Bx7 可明显提高HWM-GS 和不溶性谷蛋白大聚体的含量,使面团强度显著增强,可大大提高小麦品质[28]。在Glu-D1位点上,Dx5+Dy10是优质亚基,可改善面筋质量,本研究中含有Dx5优质基因的材料占17.22%。在全国育成的品种中,含有Dx5基因的平均频率为15.7%~ 18.6%,本试验研究结果与前人研究结果基本一致,在151 份小麦材料中,同时含有Dx5、Bx7+By8的频率为4.64%。本试验筛选出的优质品质基因可供育种时参考。
小麦籽粒中多酚氧化酶的活性与加工面食制品的颜色变化密切相关,直接影响面粉和面食制品的品质和商业价值[29-30]。低PPO 活性减少了面粉在加工和储存过程中的褐变,因此,可以通过遗传育种途径改良面粉颜色的褐变[31]。经检测发现,在151 份小麦材料中,Ppo-A1a的出现频率为51%(高PPO 活性),Ppo-A1b的出现频率为45.7%(低PPO 活性),已有研究[24]发现Ppo-A1b的分布频率为43.8%~ 53.8%,表明本文研究结果与已有研究基本吻合;Ppo-D1a的分布频率为25.8%(低PPO 活性),Ppo-D1b的分布频率为74.2%(高PPO 活性),低PPO 活性等位变异组合Ppo-A1b/Ppo-D1a在本研究中有13 份材料同时含有这两个基因,频率为8.60%;1B/1R异位系数量较多,其频率为86.75%;含有低PPO 活性优异等位组合且为非1B/1R异位系的材料只有2 份,份额较少,频率为1.32%,可用于高白度面粉小麦品种的选育。
由此可见,近几年来江苏瑞华农业科技有限公司品质育种相对单一,今后应与分子标记辅助选择紧密结合,选择具有优质基因的品种作为亲本,进行聚合杂交,提高小麦品质和产量。
猜你喜欢
上海金属(2021年6期)2021-12-02
昆明医科大学学报(2021年3期)2021-07-22
世界最新医学信息文摘(2020年68期)2020-12-25
生物学通报(2019年3期)2019-02-17
临床医药文献杂志(电子版)(2017年11期)2017-05-17
国外医药(抗生素分册)(2016年1期)2016-07-10
吉林大学学报(医学版)(2015年4期)2015-12-17
郑州大学学报(医学版)(2015年1期)2015-02-27
中国中医药现代远程教育(2014年11期)2014-08-08
赤峰学院学报·自然科学版(2013年4期)2013-07-31
