
香叶树优树自由授粉家系幼林生长性状遗传变异与初选
2023-11-08 11:55胥清利
武夷学院学报 2023年9期
胥清利
(福建省顺昌埔上国有林场,福建 顺昌 353205)
林木生长性状变异不仅受到外界环境的影响,同时受一定程度的遗传特性控制,适度的遗传变异可改变遗传改良的能力,其变异广泛存在于区组与家系间[1]。家系优良选育是推进林木遗传育种和改良进程的关键途径,探究家系生长性状的变异程度及规律是科学规划育种措施的前提,也是估算亲本家系遗传参数和遗传价值的重要环节[2],前向选择可区分母树的优劣[3],而后向选择可对优质家系资源进行筛选,提高遗传改良潜力[4]。同时自由授粉可确保亲本获得大量种子,增强林木可育性以及扩大子代变异系数[5],其技术已广泛应用于麻栎[1]和木荷[6]等树种的优树家系选择。因此深入研究香叶树自由授粉家系生长性状和遗传特性的差异性,揭示遗传控制机理,进行优良家系早期选育,对乡土珍贵树种遗传改良、优质选育和高效培育具有重要意义。生长量和保存率是阔叶树种最主要的表型性状,其中地径、胸径、树高和材积是表征林木生长状况的关键指标,生存率则侧重林木的存活状况,结合生长性状和存活率指标,可实现综合改良的目标[7]。
香叶树(Lindera communis hemsl)属樟科(Lauraceae)山胡椒属(Lindera)乔木植物,亚热带常绿树种,喜光和温暖气候,抗逆性强,耐干旱瘠薄,主要分布在海拔300~2000 m 间。香叶树是我国西南地区优质用材树种,因其材质细致、生长速度快、果实与种子含油率高、培土以及涵养水源能力强,在建筑用材和园林绿化等方面均具有重要作用[8]。目前有关香叶树的研究多集中在种子休眠、油脂和群落特征等方面[9],鲜有分析优良家系选育的报道;同时香叶树各家系间生长性状间的差异性和遗传变异机制揭示不足均限制了其目标家系的选择以及育种造林的推进。鉴于此,选取3 年生香叶树优树自由授粉家系测定林为材料,分析其生长性状的遗传变异程度与性状间的相关性,在此基础上结合主成分分析法和聚类分析法综合选取具有代表性的优良家系,以期为香叶树的优质家系选育、生产实践以及后期推广示范提供科学依据。
1 材料与方法
1.1 试验地概况与试验材料
试验地位于福建省顺昌埔上国有林场(117°43′56″E,26°57′37″N)。林场境内山地多属武夷山脉的低山丘陵地带,平均海拨300~700 m,年平均气温为18.6 ℃,最高气温40.3 ℃,最低气温零下6.8 ℃,年平均降水量1300~2100 mm,日照时数1526 h,无霜期可达305 天,属中亚热带海洋性季风气候,林地多为山地红壤,土层深厚,土壤肥沃、湿润,适合香叶树栽培生长。供试苗木材料均来自于福建省林科院,共29个家系。2019 年春,选取29 个家系的香叶树轻基质容器苗,采用完全随机分组设计,3 次重复,10 株单列小区,共870 株,株行距为2.0~3.0 m,各区组的周界种植1~2 列杉木作为隔离行,造林前每穴施复合肥0.1 kg,及时加强幼抚、早期除萌和修枝,按一般生产经营进行幼林抚育。
1.2 数据调查与处理
2022 年3 月16 日进行全林调查,调查指标包含各家系保存率 (preservation rate,PR)、地径(ground path,GP)、胸径(diameter at breast height,D) 和树高(height,H)。单株材积(volume,V)参照施恭明[10]的方法计算:
式中:V 为单株材积(m3),D 为胸径(cm),H 为树高(m)。
采用Excel 2020 进行数据整理,使用R 语言程序包“Halfsib MS”计算遗传力和变异系数。
式中:σf 为家系遗传标准差,σP 为表型标准差且σp=σf+σe(σe 为环境标准差),X 为群体平均值。
使用SPSS 25.0 进行方差分析、相关性分析、主成分分析以及聚类分析。
2 结果与分析
2.1 香叶树不同家系间生长指标比较
由表1 可知,各家系地径、树高、胸径、单株材积和保存率的分布范围分别为:1.02~7.25 cm、1.50~4.39 m、0.2~4.3 cm、0.000004~0.004168 m3和10 %~100 %。平均地径生长较高的前5 个家系依次为2 号、1 号、4号、5 号和3 号,其值处于3.72~5.45 cm 间,均比总体平均地径(2.38 cm)增加56.30%以上,其中2 号(5.45 cm)增加1.2 倍以上,显著大于其余家系;平均树高生长较高的前5 个家系依次为2 号、1 号、27 号、5 号和3 号,其值处于3.21~3.80 cm 间,均比整体平均树高(2.12 m)增加51.42%以上;平均胸径生长较好的前5 个家系依次为2 号、5 号、4 号、27 号和1 号,其值处于2.70~3.35 cm 间,其中2 号家系平均胸径显著大于其余各家系,比总体平均胸径(1.29 cm)增加1.5 倍以上;平均材积较高的前5 个家系依次为2 号、27 号、5 号、4号和1 号,其值处于0.001391~0.002431 m3间,均比平均材积(0.0004445 m3)增加2 倍以上,其中2 号家系比平均材积增加4 倍以上;保存率为100 %的家系有4 个,分别为6 号、4 号、2 号和1 号,除22 号和13号(53.33%)保存率小于60 %外,剩余家系保存率维持在60%~96.67%之间,无显著差异。

表1 香叶树各家系生长性状指标Tab.1 Growth traits of each family of L.communis
对香叶树29 个家系指标进行方差分析(表2),各家系生长指标在家系间均达到极显著水平,而区组间仅地径和单株材积达到极显著水平,说明相较于区组,香叶树4 个指标在家系间存在更广泛变异,即各家系生长方差来源主要以家系为主。同时各家系生长性状指标变异系数均在40%以上,其中单株材积变异系数最高,为85.53%,其次为胸径,75.02%。地径和树高两者变异系数接近,分别为48.74%和43.37%。这表明香叶树各形态指标存在丰富变异,表型分化明显,改良潜力大,为优良家系的选择奠定基础。
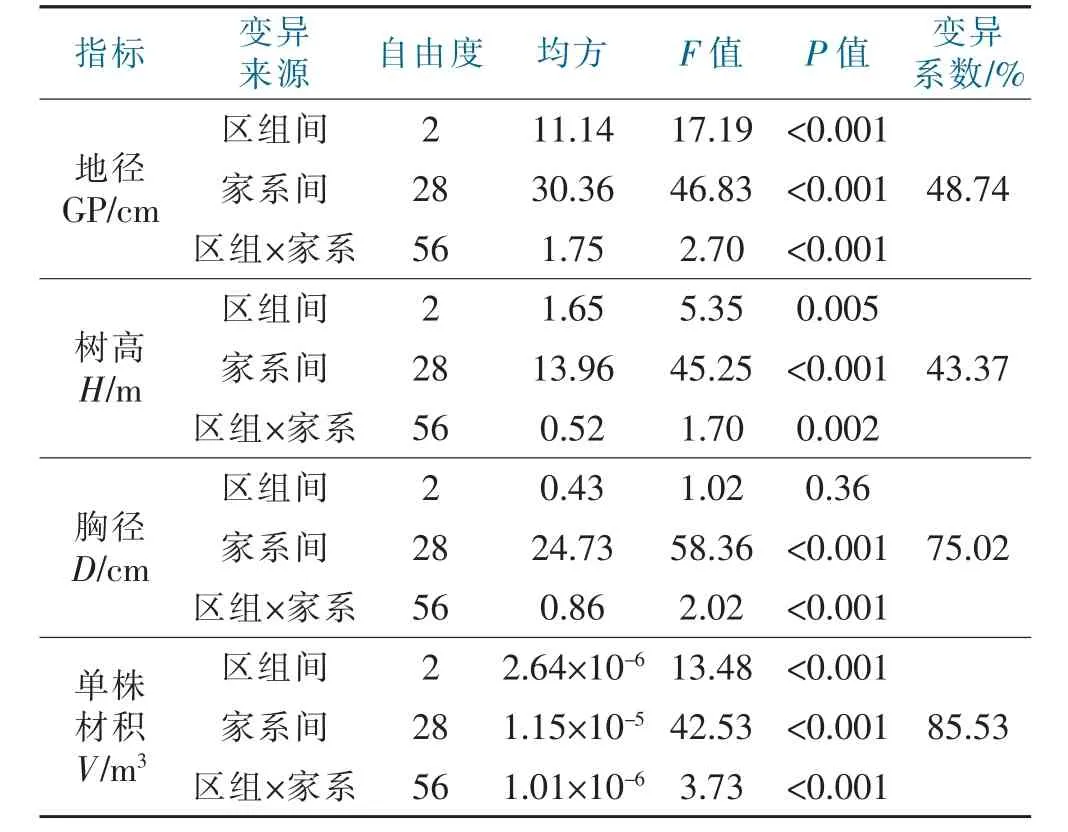
表2 家系间各指标方差分析F 值与变异系数Tab.2 Family variance analysis F value,coefficient of variation,and indicators
2.2 香叶树不同家系生长指标遗传参数
遗传参数可反映家系各生长性状的总体遗传特征,对其进行估算有助于育种方法的确定以及后续制定相匹配的选育策略(表3)。由表3 可知,各性状遗传变异系数和表型变异系数均表现为:单株材积>胸径>地径>树高,遗传变异系数范围为0.37~1.39,表型变异系数范围为0.62~2.34,两种变异系数均在0.3 以上,表明香叶树各家系整体遗传和表型变异丰富,具有优秀的选育基础,可为后续家系选育提供理论依据。遗传力方面,各性状家系遗传力大小表现为:胸径=树高>地径>单株材积,其遗传力值均处以0.9 以上,属于高度遗传范围,说明香叶树各生长性状受遗传因素影响较大,优良性状均可随着选育家系稳定遗传下去。
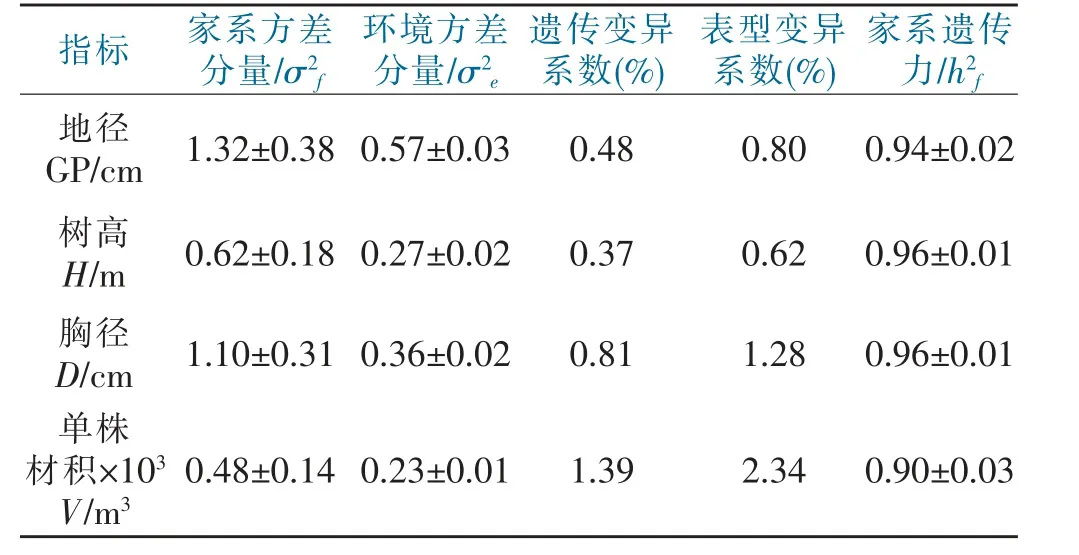
表3 香叶树各家系遗传参数估算Tab.3 The estimation of genetic parameters in L.communis
2.4 香叶树不同家系间生长指标主成分分析与选择
对香叶树29 个家系各生长指标进行主成分分析,由表5 可知第1 成分特征值为4.305,贡献率达86.099%,第2 成分特征值为0.488,贡献率为9.758%,两主成分累积贡献率高达95.857%,涵盖香叶树5 个生长指标的绝大多数信息,可作为评价不同家系香叶树的指标。第1 主成分权重占比较高的为:地径、树高、胸径、材积和保存率,可体现树木的整体生长状况,而第2 主成分保存率的权重相对较高,侧重反映树木的存活状况。
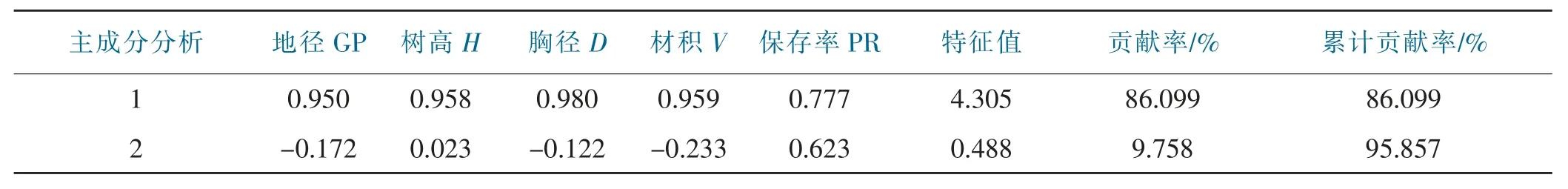
表5 香叶树各家系生长性状指标主成分分析Tab.5 Principal component analysis of growth traits of each family of L.communis
依据主成分分析,对不同家系各成分贡献率进行得分排序(表6)。X1、X2、X3、X4和X5分别表示标准化后的地径、树高、胸径、材积和保存率值。第1 主成分F1得分表达式为:F1=0.458X1+0.462X2+0.472X3+0.462X4+0.374X5;第2 主成分F2得分表达式为:F2=0.033X2-0.246X1-0.175X3-0.333X4+0.891X5;综合得分F 表达式为:F=0.861F1+0.098F2。按第1 主成分得分对18 个家系进行排序,得分为正数的有7 个,分别为:2号、1 号、4 号、5 号、27 号、6 号,其分值均处于2.49 分以上;按第2 主成分得分进行排序,得分为正数的有15 个,前6 名分别为:21 号、24 号、26 号、7 号、20 号和15 号,其分值均处于0.35 分以上;综合得分大于0的有8 个,排前6 名的分别为:2 号、1 号、4 号、5 号、6号和27 号。
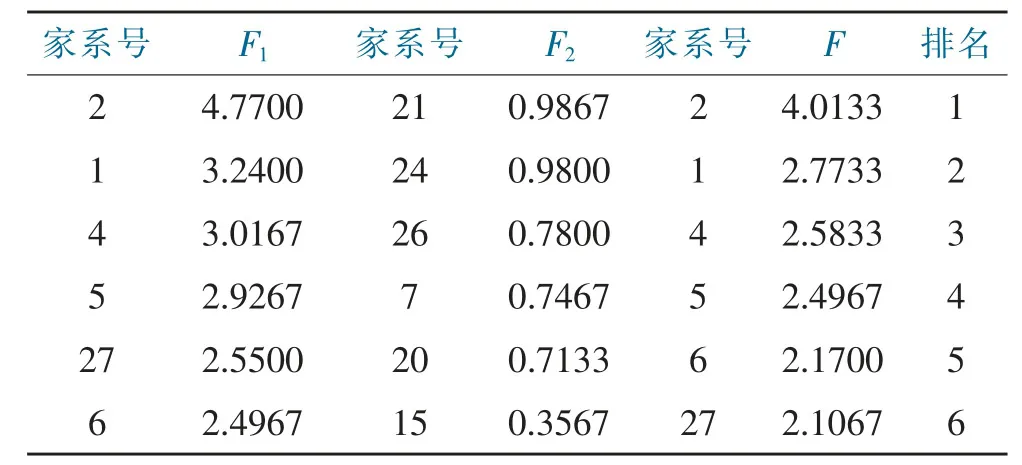
表6 香叶树各家系主成分得分及排名Tab.6 The scores and rankings of principal components of each family of L.communis
2.5 香叶树家系间生长指标聚类分析与选择
依据香叶树不同生长指标的综合表现,采用组间欧式距离对29 个家系进行聚类分析(图1)。以欧氏距离10 为阈值,将香叶树29 个家系分成3 类:第1 类仅有22 号家系,性状表现最差,各主成分得分均为最低;第2 类包含7 种家系,分别为1 号、2 号、3 号、4号、5 号、6 号和27 号家系;第3 类为剩余的21 个家系,性状表现中等,除20 号家系主成分综合得分略大于0 外,其余家系得分均为负数。
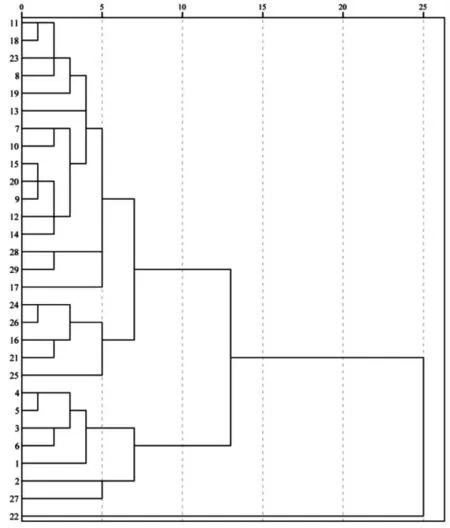
图1 香叶树29 个各家系聚类分析Fig.1 Cluster analysis of 29 provenances of L.communis
2.6 香叶树家系早期选择
以各家系具体生长指标数值为基础上,利用主成分分析和聚类分析结果,对各家系进行综合评价,采用20%的优良家系入选率[3]开展福建香叶树早期优良家系选择,选取2 号、1 号、5 号、4 号、27 号和6 号为最优家系。其中,入选家系地径均值为4.19 cm,树高均值为3.35 m、胸径均值为2.85 cm、单株材积均值为0.001568 m3和保存率均值为96.11%,比群体平均值分别增加了76.20%、58.32%、118.53%、252.55%和23.69%。
3 讨论
3.1 香叶树各家系生长量与保存率分析
对香叶树各自由授粉家系筛选评价旨在推广示范及提升当地林分精准质量,而生长量与保存率受自身遗传特性以及外界环境的影响,能较大程度反映对当地的适应情况,表达各家系整体生长状况[11]。本研究主要对象为3 年生香叶树,生长期早,平均生长量相对较低,但总体保存率高达77.7%,其中部分家系保存率达到100%,且相应生长量显著大于平均生长量,通过综合选育可获得适合福建本土的优良特色香叶树家系。29 个家系中保存率最高的有1 号、2 号等家系(100%),保存率最低的为22 号家系(23.33%),不同家系保存率相差较大可能原因为:各家系抗风、抗寒能力参差不齐;初期采种时种子成熟度存在差异[3];受外界环境影响,如白蚁的局部侵蚀,确凿原因有待后续进一步探究。
3.2 香叶树各家系生长性状差异性与遗传力分析
遗传与变异是林木遗传改良与选择的关键因素,其潜力取决于遗传变异的发掘与利用[12]。相关研究表明[13],具有类似遗传特性树种的遗传变异不仅存在于林分、种间及种内,且由于环境和栽培措施的差异性,各种因素间存在一定的交互作用。表型为本身遗传特性以及外界环境因素共同影响所形成,苑海静[1]对麻栎优树家系研究发现,地上单株生物量存在家系与区组间显著互作效应,且区组效应为主要因素。本研究表明,香叶树各家系地径、胸径、树高和材积在家系、区组×家系均表现为极显著差异,且家系效应远大于区组与区组×家系互作效应,与王云鹏[15]对10 年生木荷(Schima superba)家系木材基本密度受家系效应影响最大结果相同,表明各家系生长性状同时受到自身遗传特征以及环境影响,但主要由遗传基因型控制,选择潜力大,具有丰富的家系变异基础,有利于优良家系的综合选育。
遗传参数估算是林木选育的必要途径,可反映物种的遗传多样性,而性状遗传力是关键遗传参数之一,体现基因型的优劣程度。王云鹏[6]研究发现10 年生木荷胸径和树高均受到中度以上的家系控制;肖德卿[16]研究表明栽培模式可改变木荷家系遗传特性。本研究发现香叶树地径、树高、胸径和材积的家系遗传力均处于0.9 以上,属于强遗传控制,与彭洋[17]对棕榈[Trachycarpus fortunei(Hook.)H.Wendl.]半同胞家系评价结果相似,表明香叶树各生长性状主要受家系遗传特性控制,家系亲本遗传资源丰富。变异系数可衡量群体的分散程度,控制选择范围,是决定生长指标遗传潜力的重要指标。方差分析显示香叶树各性状变异系数处于43%以上,其中材积变异系数高达85.53%,表明香叶树各性状分化明显,材积较其余性状变异相对丰富。遗传参数估算结果显示,各性状表型变异系数均大于遗传变异系数,表明由于外部环境的影响,香叶树自身遗传性状特征会小于所对应的表型性状。材积的遗传和表型变异系数均大于胸径、地径和树高,与欧阳天林[18]对木荷种子园的结果相似,这表明在香叶树各生长性状中,材积的选择空间及潜力最大。其原因可能是林木对同质资源的竞争规律,即优势林木在早期生长时为了争夺光质资源与生存空间,优先选择树高生长策略,而胸径间存在差异性,从而由胸径和树高公式计算而来的材积差异性则更为显著[19]。
3.3 香叶树优良家系早期综合筛选与评价
主成分分析法是基于保存绝大多数优良基因资源前提下,将多个相关性指标压缩转换成少数个独立主成分综合指标的多元分析方法,简化了评价结构流程,避免信息重叠和主观性,使其结果更具科学合理性,广泛应用于各林木家系选育分析中[19]。本研究将地径、树高、胸径、材积和保存率5 个生长性状指标转换成2 个主成分,两者累计贡献率高达95.857%,可表述绝大多数指标信息。通过各性状指标权重计算家系综合得分,并按照20%的入选率筛选得分排序前6 的家系,其品质综合表现最佳,可为种质选育提供实践参考。
聚类分析方法对各家系进行分析归类,保证同类具有相似的遗传信息表达,可客观简明阐述各家系整体优劣性,梳理家系间亲远度,利于亲本选择以及优良种质筛选[20]。聚类分析将香叶树29 个家系划分为3个类群,分别为最优家系、中等家系以及最劣家系,与生长指标及得分密切相关,为后续优良家系选育奠定基础。
分析林木表型生长性状,探究遗传变异特性与规律,明晰其变异特点与丰富度,对香叶树优良家系选择和开发利用起到至关重要的作用。于志民[20]研究发现,在主成分分析基础上采用聚类分析方法可使亲本遗传差异性增大,产生更为广泛的变异,有利于筛选出适应性较强的家系。本研究在具体生长性状值和将多个指标提炼成个数指标的主成分分析法基础上,以聚类分析进行整体分析,按照20%入选率,选育出2号、1 号、5 号、4 号、27 号和6 号为最优家系。这些家系可作为后续香叶树优良种质选育和促进林木生长的备选材料,其研究可为香叶树优树自由授粉家系遗传改良、种质推广、优质林分的营造,生态修复及景观提升,发挥良好的示范带动作用。
4 结论
3 年生香叶树优树29 个自由授粉家系间存在丰富的遗传变异,具有较高的改良潜力,地径、胸径、树高和单株材积在家系间呈极显著差异,且遗传力均处于0.9 以上,表现为强度遗传控制。主成分分析选育出的2 号、1 号、5 号、4 号、27 号和6 号共6 个家系最优家系,在聚类分析中属于同一类,且得分均处于前列,各生长性状显著大于群体平均水平,可作为后续香叶树优良种质选育和促进乡土林木生长的备选材料。
猜你喜欢
少年文艺·我爱写作文(2023年2期)2023-02-24
世界热带农业信息(2022年11期)2022-10-18
吉林林业科技(2020年6期)2020-12-16
南方文学(2017年6期)2018-01-07
作文周刊·小学一年级版(2016年20期)2017-06-03
长城(2017年1期)2017-02-17
西南农业学报(2016年6期)2016-04-16
武夷学院学报(2015年3期)2015-07-18
长春工程学院学报(自然科学版)(2013年4期)2013-12-06
食品与机械(2012年2期)2012-12-28
