
外源γ-氨基丁酸延缓采后猕猴桃果实冷害及其与活性氧代谢的关系
2023-11-07 11:09夏明辉赵云峰陈发河林艺芬陈福泉
食品科学 2023年19期
夏明辉,张 珅,,赵云峰,汪 莹,何 凡,陈发河,林艺芬,陈福泉,
(1.集美大学海洋食品与生物工程学院,福建 厦门 361021;2.盐城工学院海洋与生物工程学院,江苏 盐城 224051;3.亚热带特色农产品采后生物学(福建农林大学)福建省高校重点实验室,福建 福州 350002;4.福建农林大学食品科学学院,福建 福州 350002)
猕猴桃为猕猴桃科(Actinidiaceae)猕猴桃属(Actinidia)藤本植物,原产于我国,在我国南北方地区均有大规模种植,果实产量居世界首位,是重要的大宗经济水果[1],并且具有很高的营养价值[2]。猕猴桃属于呼吸跃变型果实,其采后成熟过程的重要特征为接近完熟时果实迅速软化,导致果实贮运期受限,货架期和最佳食用期较短[3],因此在其采收和商品化处理、流通过程中广泛采用适当早采、单果内衬和箱式包装等方式[4],以提高果实耐贮运性,并结合冷藏、气调贮藏等手段以延缓后熟过程,延长其保鲜期[3]。作为冷敏型果实,猕猴桃在不适宜的低温条件下极易发生冷害,主要表现为果肉颗粒状木质化和水浸状腐败,严重降低果实的食用品质,导致贮藏、运输、流通和销售过程中产生巨大的经济损失[5]。
国内外研究人员采用许多化学和物理方法,如1-甲基环丙烯[9]、臭氧[10]、乙烯[11]、草酸[12]、褪黑素[13]以及气调[14]处理等,通过维持活性氧代谢酶系统较高的清除能力延缓果实冷害的发生。在诸多化学方法中,外部施用植物内源调节物质的方法具有较广的适用性,也更有助于深入发掘果蔬冷害发生与控制的内在调控点,可产生良好的冷害控制效果,加深对于相关机制的理论认知[15-17]。γ-氨基丁酸(γ-aminobutyric acid,GABA)是广泛存在于生物体内的天然氨基酸,具有天然无毒、极易获取、成本低等特点,并且可调节植物体内多种胁迫反应[18]。研究表明,GABA可以通过提高抗氧化酶活性[19]、缓解氧化应激反应[19]、维持能量水平[20]、减轻膜脂过氧化程度[21]等作用提高西葫芦[20]、番石榴[22]、印度油柑[23]等果蔬的组织抗冷性,减轻低温伤害。但是,目前鲜见将GABA应用于猕猴桃采后冷害控制的报道;GABA是否对冷藏猕猴桃果实活性氧清除系统,尤其是非酶清除系统具有调控效应,从而调节氧化还原平衡,增强组织抗冷性,也值得深入研究。因此,本实验采用GABA溶液对采后猕猴桃果实进行贮前处理,研究GABA处理对冷藏猕猴桃果实冷害发生和活性氧产生、活性氧清除酶系统和非酶系统的影响,旨在为采后猕猴桃果实冷害控制实践及调控机制的研究提供参考。
1 材料与方法
1.1 材料与试剂
猕猴桃果实采自福建省三明市建宁县聚晟源现代农业有限公司,果实采收后经单果套网袋和瓦楞纸箱包装,由厢式货车于5 h内运至集美大学农产品加工与贮藏实验室(厦门),选取约9 成熟且形状大小均一、无机械伤和病虫害的果实作为实验材料。
氢氧化钠、盐酸羟胺、对氨基苯磺酸、乙酸、三氯乙酸、硫代巴比妥酸、α-萘胺、亚硝酸钾、二甲酚橙、硫酸亚铁铵、乙二胺四乙酸、聚乙烯吡咯烷酮、H2O2、二硫苏代糖醇、蛋氨酸、氮蓝四唑 国药集团化学试剂有限公司;核黄素、二硫代硝基苯酸钾、GSH、氧化型谷胱甘肽、还原型辅酶II四钠盐、AsA、还原型辅酶I、抗坏血酸氧化酶、单脱氢抗坏血酸(monodehydroascorbic acid,MDHA)、磷酸二氢钠、磷酸氢二钠 阿拉丁(上海)试剂有限公司;以上试剂均为分析纯。
1.2 仪器与设备
质构仪 厦门超技仪器设备有限公司;全自动分析天平 上海花潮实业有限公司;UV-8000紫外-可见分光光度计 上海元析仪器有限公司;SYG-1220数显恒温水浴锅 美国精骐有限公司;JM-05D-80超声波清洗机广东洁盟超声实业有限公司;PSX-330H智能恒温恒湿箱宁波莱福科技有限公司;DDS-11A电导率仪 上海大普仪器有限公司。
1.3 方法
1.3.1 果实采后处理及贮藏
前期预实验已筛选出2.0 mmol/L为较适宜的GABA处理浓度。将猕猴桃果实随机分为两组,每组300 个果实。一组果实用2.0 mmol/L GABA溶液浸泡20 min,另一组果实以蒸馏水代替GABA溶液浸泡作为对照组,处理后的果实在20 ℃下静置1 h,自然晾干,然后置于(4±1)℃、(85±5)%相对湿度的冷库中贮藏。贮藏期内每10 d随机取40 个猕猴桃果实样品进行相关指标的测定。
1.3.2 指标测定
1.3.2.1 细胞膜透性
参照Zhang Zhengke等[24]的方法,随机选取10 个猕猴桃果实,用打孔器(8 mm)在每个果实赤道面上取3 片2 mm厚的薄片,用电导率仪分别测定自然渗出电解质溶液和组织破坏后渗出电解质溶液的电导率,结果以相对电导率表示。
1.3.2.2 冷害指数
参考Li Ziying等[25]的方法,根据猕猴桃果实去皮后表面凹陷和水渍状部分占整果表面积比例,将冷害指数分为4 级:0级为无冷害症状;1级为冷害发生面积比例小于25%;2级为冷害发生面积比例大于25%且小于50%;3级为冷害面积比例大于50%且小于75%;4级为冷害发生面积比例大于75%。贮藏过程中及时剔除因微生物侵染而腐烂的果实,之后不再列入统计。冷害指数按式(1)计算。
1.3.2.3 丙二醛含量
参考Lin Yu zhao 等[26]的方法进行丙二醛(malondialdehyde,MDA)含量的测定,随机从20 个猕猴桃果实中取2.0 g果肉,采用硫代巴比妥酸比色法测定其在532 nm波长处的吸光度,分别在420 nm和600 nm波长处测定吸光度用于消除误差,结果以μmol/g为单位。
H2O2含量参考王俊文[28]的方法,随机从20 个猕猴桃果实中取2.0 g果肉,采用二甲酚橙显色法进行测定,以H2O2标准品作标准曲线,结果以μmol/g为单位。
1.3.2.5 可溶性蛋白含量
参考Lin Yifen等[27]的方法,采用考马斯亮蓝G-250比色法测定可溶性蛋白含量,以牛血清白蛋白作标准曲线,其含量用于后续酶活力计算。
1.3.2.6 活性氧清除酶活力
SOD活力的测定参考杨乾等[29]的方法,随机从20 个猕猴桃果实中取2.0 g果肉,以每分钟酶反应体系对氮蓝四唑光化学还原抑制50%为1 个活力单位(1 U)。CAT活力的测定参考Cakmak等[30]的方法,以每分钟反应溶液在240 nm波长处吸光度变化0.01为1 U。APX活力的测定参考Ren Yanfang等[31]的方法,以每分钟反应体系在290 nm波长处OD值变化0.01为1 U。上述酶活力结果均以U/mg pro为单位。
1.3.2.7 AsA和GSH含量
参考胡苗等[32]的方法,随机从20 个猕猴桃果实中取2.0 g果肉,采用比色法进行AsA含量的测定,对其还原的亚铁离子与红菲咯啉反应生成物进行比色;通过与5.5’-二硫双对硝基苯甲酸(5.5’-dithiobis(2-nitrobenzoic acid),DTNB)反应测定GSH含量,分别以AsA和GSH标准品作标准曲线,AsA和GSH含量分别以mg/100 g和μmol/g为单位。
1.3.2.8 AsA-GSH循环相关酶活力
参考王懿等[33]的方法,随机从20 个猕猴桃果实中取2.0 g果肉,谷胱甘肽还原酶(glutathione reductase,GR)活力以每分钟反应体系在340 nm波长处吸光度减少0.01为1 U;MDHA还原酶(monodehydroascorbate reductase,MDHAR)活力以消耗1 μmol NADH为1 U;脱氢抗坏血酸还原酶(dehydroascorbate reductase,DHAR)活力以生成1 μmol的AsA为1 U。结果均以U/mg pro为单位。
1.3.2.9 1,1-二苯基-2-三硝基苯肼自由基清除率和总还原力
参考Chen Chen等[34]的方法,随机从20 个猕猴桃果实中取2.0 g果肉,采用1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)自由基清除法和亚铁离子法分别测定DPPH自由基清除率及总还原力,分别在517 nm和700 nm波长处测定吸光度,DPPH自由基清除率采用公式(2)计算。总还原力以700 nm波长处吸光度表示。
1.4 数据统计与分析
每组实验进行3 次重复,使用Microsoft Excel 2019软件对实验数据进行处理并绘图。采用SPSS Statistics 26软件中单因素方差分析对结果进行差异显著性分析。采用Origin 2021软件对数据进行相关性分析和主成分分析(principal components analysis,PCA)并绘图。
2 结果与分析
2.1 GABA处理对冷藏猕猴桃果实冷害的影响
如图1A所示,冷藏猕猴桃果实的冷害指数随着贮藏时间的延长逐渐增加,与对照组果实相比,处理组果实的冷害指数在20~40 d和60 d时显著较低(P<0.05,P<0.01)。图1B显示,对照组猕猴桃果实的细胞膜透性在0~10 d迅速上升,随后缓慢升高。由于采后果实接近完熟时细胞膜透性会快速上升,可推断本研究供试果实冷藏条件下的快速后熟期为贮藏0~10 d。GABA处理组果实细胞膜透性的变化趋势与对照组一致,且在整个贮藏期内显著低于对照组(P<0.05,P<0.01)。由图1C可知,猕猴桃果实MDA含量随贮藏时间的延长逐渐增加;与对照组相比,经GABA处理的果实MDA含量在贮藏0~30 d缓慢升高,其中在10~30 d和60 d时显著较低(P<0.05,P<0.01)。


图1 GABA处理对冷藏猕猴桃果实冷害指数(A)、细胞膜透性(B)和MDA含量(C)的影响Fig.1 Effect of GABA treatment on chilling injury index (A),cell membrane permeability (B),and MDA content (C) in kiwifruit during cold storage
2.2 GABA处理对冷藏猕猴桃果实产生速率和H2O2含量的影响
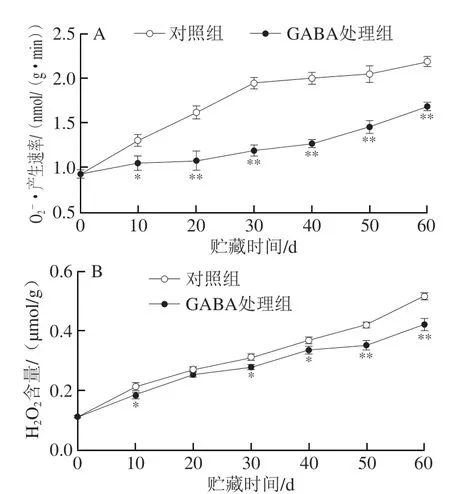
图2 GABA处理对冷藏猕猴桃果实产生速率(A)和H2O2含量(B)的影响Fig.2 Effect of GABA treatment on superoxide anion radical production rate (A) and H2O2 content (B) in kiwifruit during cold storage
2.3 GABA处理对冷藏猕猴桃果实活性氧清除酶活力的影响
如图3A所示,对照组和GABA处理组果实的SOD活力均在0~10 d上升,之后随着贮藏时间的延长不断下降;与对照组相比,GABA处理组果实的SOD活力在10~30 d和50 d时显著较高(P<0.05,P<0.01)。由图3B可知,对照组果实CAT活力在整个贮藏期内逐渐下降,而GABA处理组果实CAT活力在0~10 d升高,10~20 d迅速下降,随后呈缓慢降低趋势,并且在整个贮藏期内与对照组相比显著较高(P<0.05,P<0.01)。图3C显示,在整个贮藏期间,对照组和GABA处理组果实APX活力均在贮藏0~20 d上升并到达高峰,20~60 d逐渐下降;相较于对照组,GABA处理组果实APX活力在20 d和40~60 d时显著较高(P<0.05,P<0.01)。

图3 GABA处理对冷藏猕猴桃果实SOD(A)、CAT(B)和APX(C)活力的影响Fig.3 Effect of GABA treatment on SOD (A),CAT (B) and APX (C)activities in kiwifruit during cold storage
2.4 GABA处理对冷藏猕猴桃果实AsA和GSH含量的影响
如图4A所示,对照组果实AsA含量在0~10 d快速升高,10~30 d缓慢上升,30~60 d不断下降;与对照组相比,GABA处理组果实AsA含量在0~10 d升高较快,10~30 d维持在较高水平,随后逐渐下降,且在贮藏10~60 d极显著较高(P<0.01)。由图4B可知,对照组和GABA处理组果实的GSH含量均在0~40 d逐渐上升,40~60 d迅速下降,且与对照组相比GABA处理组果实GSH含量在10~40 d和60 d时显著较高(P<0.05,P<0.01)。
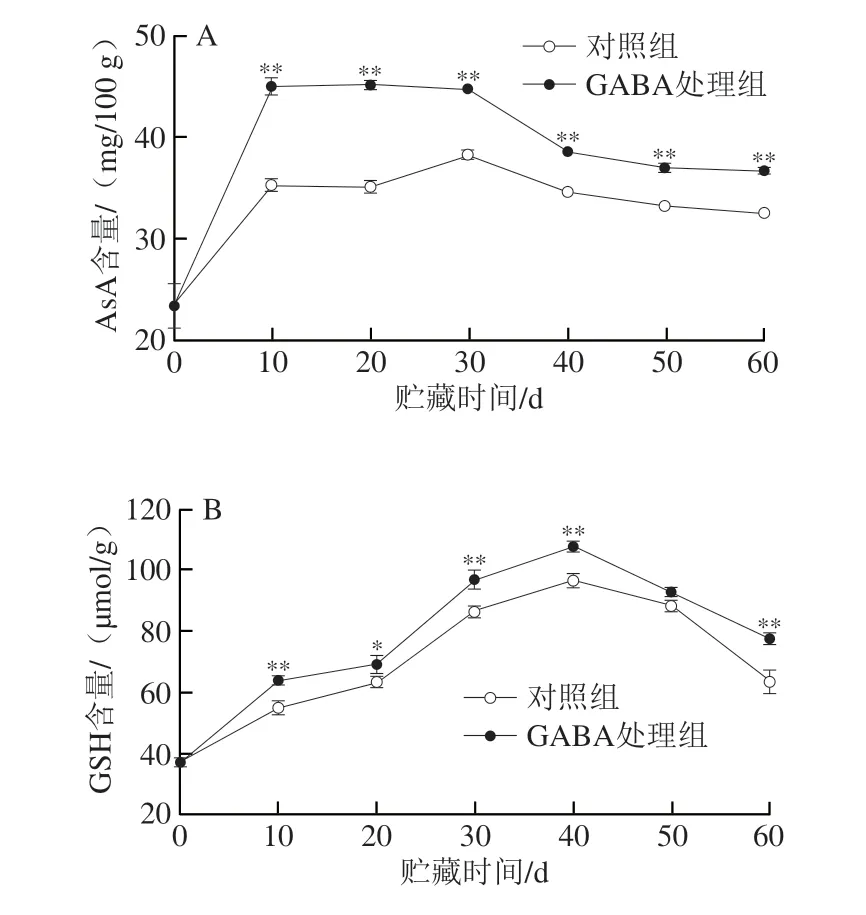
图4 GABA处理对冷藏猕猴桃果实AsA(A)和GSH(B)含量的影响Fig.4 Effect of GABA treatment on the contents of AsA (A) and GSH (B)in kiwifruit during cold storage
2.5 GABA处理对冷藏猕猴桃果实AsA-GSH循环酶活力的影响
如图5A所示,对照组果实GR活力在贮藏0~40 d缓慢升高,40~60 d快速下降;而GABA处理组果实的GR活力在0~30 d迅速上升,30~60 d迅速下降。与对照组相比,GABA处理组果实的GR活力在30~40 d极显著较高(P<0.01)。图5B显示,对照组果实MDHAR活力在0~30 d上升并达到高峰,30~50 d迅速下降后趋于稳定;GABA处理组果实的MDHAR活力变化趋势与对照组一致,并在20~30 d显著较高(P<0.05,P<0.01)。由图5C可知,对照组果实DHAR活力在贮藏期内变化较平稳,0~40 d缓慢升高,随后有所降低;与对照组相比,GABA处理组果实DHAR活力在0~30 d快速上升并达到峰值,30~40 d迅速下降,随后较为稳定,且在整个贮藏期间均极显著较高(P<0.01)。

图5 GABA处理对冷藏猕猴桃GR(A)、MDHAR(B)和DHAR(C)活力的影响Fig.5 Effect of GABA treatment on the activities of GR (A),MDHAR (B) and DHAR (C) in kiwifruit during cold storage
2.6 GABA处理对冷藏猕猴桃果实DPPH自由基清除率和总还原力的影响
如图6所示,对照组和GABA处理组果实的DPPH自由基清除率和总还原力均随着贮藏时间的延长而下降,表明果实抗氧化性逐渐降低。而在相同贮藏期内,与对照组相比,GABA处理组果实具有显著较高的抗氧化性(P<0.05,P<0.01)。
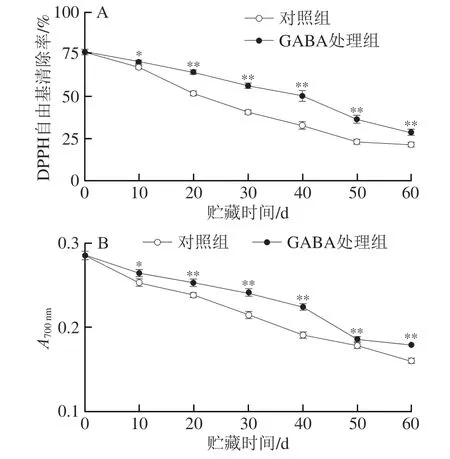
图6 GABA处理对冷藏猕猴桃果实DPPH自由基清除能力(A)和总还原力(B)的影响Fig.6 Effect of GABA treatment on DPPH radical scavenging capacity (A)and total reducing power (B) in kiwifruit during cold storage
2.7 相关性分析及PCA
如图7A所示,在冷藏期间,对照组猕猴桃果实冷害指数与细胞膜透性呈显著正相关(r=0.83,P<0.05)。细胞膜透性和MDA含量呈显著正相关(r=0.87,P<0.05),MDA含量与产生速率(r=0.94)、H2O2含量(r=0.97)均呈显著正相关(P<0.05)。产生速率与SOD活力呈显著负相关(r=-0.86,P<0.05)。H2O2含量与CAT活力呈显著负相关(r=-0.93,P<0.05)。MDHAR和DHAR活力均与AsA含量呈正相关,GR活力与GSH含量呈显著正相关(r=0.94,P<0.05)。冷害指数(r=-0.96)和细胞膜透性(r=-0.93 )均与总还原力呈显著负相关(P<0.05)。总还原力与产生速率(r=-0.95)和H2O2含量(r=-0.99)均呈显著负相关(P<0.05),并与SOD活力(r=0.84)和CAT活力(r=0.91)呈显著正相关(P<0.05)。采用PCA法分析了对照组和GABA处理组猕猴桃果实冷害相关指标和活性氧代谢相关酶的活力。如图7B所示,通过分析协方差矩阵的特征值,发现两个主成分(PC)占数据集总方差的86.6%,表明降维后的数据能够很好地表征原始数据。PC1涵盖了数据集中53.6%的方差,主要体现果实冷害、活性氧水平和活性氧清除酶活力的特征值变化。随着贮藏时间的延长,对照组和GABA处理组的负荷值在PC1上均不断增大,表明活性氧清除酶活力逐渐下降,活性氧不断积累,冷害指数升高。与对照组相比,GABA处理组果实能够维持较低的负荷值,说明GABA处理能够较好地维持冷藏猕猴桃果实活性氧清除酶系统的清除能力,延缓活性氧积累,减缓冷害的发生。PC2涵盖了数据集中33.0%的方差,主要体现活性氧清除非酶系统(AsA-GSH循环)与冷害和活性氧水平之间的关系。随着贮藏时间的延长,对照组和GABA处理组的负荷值在PC2上均先增大后减小,AsA-GSH循环中相应物质含量和酶活力在贮藏中期均达到高峰。与对照组相比,GABA处理组果实能够使以上指标维持较高的负荷值,说明GABA处理能够提高AsAGSH循环的活力,延缓活性氧的积累及冷害的发生。

图7 冷藏猕猴桃果实冷害和活性氧代谢相关指标的相关性分析(A)和PCA(B)Fig.7 Correlation analysis (A) and principal components analysis plot(B) of chilling injury related indexes versus ROS of postharvest kiwifruit during cold storage
3 讨论
细胞膜是果实细胞内外生理生化状态变化的重要反映者,是维持细胞稳态的关键屏障,而果蔬冷害发生的初级与次级反应均与细胞膜的相态、结构和功能等变化有关[35]。作为植物低温胁迫下的主要次级反应,活性氧的大量产生与积累会直接影响细胞膜系统和生物大分子,造成细胞结构和功能损伤,导致膜透性升高[12]。MDA作为活性氧攻击细胞膜脂的产物,也具有较强的氧化性,会进一步加重细胞膜的损伤,从而加剧冷害症状。研究表明,外源GABA处理能够延缓樱桃[19]、番石榴[22]和桃[36]等果实冷藏期间冷害指数、细胞膜透性和MDA含量的上升,从而延缓果实冷害。Huang Hua等[37]研究发现,采用苹果酸处理可以延缓香蕉果实产生速率和H2O2含量的上升,从而减缓冷害进程。本研究结果表明,猕猴桃果实冷害指数、细胞膜透性和MDA含量在低温贮藏过程中都随着时间的延长而升高(图1),同时产生速率和H2O2含量不断上升(图2)。相关性分析结果表明,细胞膜透性与冷害指数和MDA含量在整个贮藏期间均呈显著正相关,而MDA含量与产生速率和H2O2含量在整个贮藏期内均呈显著正相关(图7A)。由PCA结果可知,冷害指数、细胞膜透性、MDA含量、产生速率和H2O2含量都在PC1中具有较高的正载荷值,且GABA处理组果实的正载荷值低于对照组果实(图7B)。由此推断,猕猴桃果实冷害的发生与冷藏过程中活性氧大量积累从而加剧膜脂过氧化,进而破坏细胞膜结构有关。GABA处理显著抑制了冷藏猕猴桃果实冷害指数、细胞膜透性和MDA含量的上升(图1),同时延缓了产生速率和H2O2含量的上升(图2),说明GABA处理可以通过延缓活性氧的积累,减少果实组织中MDA的产生,减轻细胞膜脂过氧化反应,保持细胞膜较好的完整性,减轻果实在低温胁迫下的功能失调。
低温胁迫条件下,植物体内活性氧产生与清除的不平衡会引发哈密瓜[8]、甜瓜[29]和杏[38]等果实体内产生氧化胁迫,加剧其生理失调与冷害症状。许多研究表明,减轻氧化胁迫对维持冷敏型果实细胞膜完整性、提高其抗冷性、延缓低温冷害有重要作用,其内在主导因素为酶和非酶系统组成的活性氧清除系统[29]。活性氧清除酶系统是主要的活性氧清除方式,其中SOD专一歧化形成H2O2,CAT和APX可将H2O2还原为H2O和O2[39],从而减轻氧化损伤。何欢等[38]研究发现,褪黑素处理能够维持杏果实较高的SOD、CAT和APX活性,延缓产生速率和H2O2含量的上升,减轻细胞膜损伤,延缓果实冷害。Vali等[19]报道,采用GABA处理延缓了樱桃果实中和H2O2的累积,并且伴随着SOD、CAT和APX活性的增加,表明GABA可以通过调节活性氧清除酶系统活性来延缓樱桃果实的冷害。本研究中,对照组果实在贮藏期内不断升高的产生速率与SOD活力呈显著负相关,同样持续增加的H2O2含量与不断下降的CAT活力呈显著负相关(图7A)。PCA结果表明,在PC1中产生速率和H2O2含量具有较高的正载荷值,SOD和CAT活力具有较高的负载荷值,并且GABA处理组上述各点的载荷值均低于对照组(图7B)。由此可知,冷藏猕猴桃果实活性氧清除酶(SOD、CAT和APX)活力在果实快速成熟的0~10 d期间均升高,可能是由于接近完熟时相关调控基因的表达与蛋白合成增加,以应对较高的活性氧产生速率,维持氧化还原稳态,减轻细胞损伤;随后SOD和CAT活力随着贮藏时间的延长迅速降低,APX活力在20 d后快速下降,表明活性氧清除酶系统活力下降,活性氧大量积累,细胞氧化损伤加剧,果实冷害进程加快。与对照组相比,GABA处理延缓了果实产生速率和H2O2含量的上升(图2),并且增大了SOD、CAT和APX的活力(图3)。由此可推断,GABA处理可以通过增强冷藏猕猴桃果实活性氧清除酶系统的能力延缓活性氧积累,从而减轻猕猴桃果实的冷害症状。
AsA和GSH作为采后果实中的重要抗氧化活性物质,在非酶促活性氧清除系统中起着重要的作用[40]。AsA可通过自身还原性或作为电子供体在APX催化下清除H2O2[33,41],氧化生成MDHA。MDHA可由MDHAR以NADH为电子供体催化还原,重新形成AsA[33],或进一步氧化成脱氢AsA,而后者通过DHAR作用亦可还原为AsA,使AsA再生;DHAR也可将GSH氧化成氧化型谷胱甘肽(oxidized glutathione,GSSG),GSSG可由GR催化还原为GSH,构成AsA-GSH循环[42]。此外,GSH自身也可清除活性氧,生成GSSG[29]。采用甘氨酸甜菜碱或NO处理均能通过提高冷藏桃果实MDHAR、DHAR和GR活性来维持AsA和GSH水平,减少活性氧积累,延缓冷害进程[33,43]。本研究发现,对照组猕猴桃果实AsA含量在后熟期不断增加(图4A),在0~10 d快速升高,并于10~30 d维持在较高水平,APX、MDHAR、DHAR活力分别在0~20 d、0~30 d和0~40 d逐渐增加达到高峰,随后逐渐下降(图3C及图5B、C);而GSH含量和GR活力均在0~40 d快速增加,随后下降(图4 B、5 A)。相关性分析表明,对照组果实的MDHAR(r=0.60)和DHAR活力(r=0.61)在贮藏期内与AsA含量呈正相关,DHAR活力和GSH含量呈显著正相关(r=0.86,P<0.05),GR活力和GSH含量呈显著正相关(P<0.05)(图7A)。PCA结果表明,在PC2中AsA和GSH含量与MDHAR、DHAR和GR活力都具有较高的正载荷值,其中对照组和GABA处理组果实的上述载荷值在贮藏30~40 d时最高,且GABA处理组的正载荷值均高于对照组(图7B)。由此可推断,在猕猴桃果实快速后熟期(0~10 d),MDHAR、DHAR和GR活力的快速上升促进了AsA和GSH积累,这可能与冷藏初期果实对低温胁迫的响应有关;随着冷藏的进行,MDHAR、DHAR和GR活力在贮藏中期果实完熟时(30~40 d)达到高峰,此时AsA-GSH循环的活性氧清除能力最高,以抵御低温导致的活性氧爆发;而后随着贮藏时间延长,AsA和GSH含量以及MDHAR、DHAR和GR活力下降,即AsA-GSH循环活性氧清除能力降低,致使活性氧加速积累,细胞膜损伤加剧。GABA处理组果实相较于对照组保持了较高的AsA和GSH水平(图4)以及MDHAR、DHAR和GR活力(图5),说明GABA处理能够促进AsAGSH循环运行,提升活性氧非酶清除系统的活性,从而减轻冷害。
果实组织的DPPH自由基清除率和总还原力可用来衡量其抗氧化能力[26]。本研究中,在贮藏期内不断降低的DPPH自由基清除率和总还原力与冷害指数、细胞膜透性、产生速率和H2O2含量呈显著负相关(图7A),与活性氧代谢酶(SOD和CAT)活力呈显著正相关(图7A)。推测猕猴桃果实冷害进程中,果实内活性氧大量积累,活性氧清除能力降低,导致了组织抗氧化能力减弱。GABA处理能够使枇杷果实在冷藏期间维持较高的DPPH自由基清除率和总还原力(图6),说明GABA处理维持了果实的抗氧化能力,从而减轻氧化胁迫,延缓细胞膜损伤,延迟果实冷害进程。
猜你喜欢
河北果树(2020年4期)2020-11-26
中国果树(2020年2期)2020-07-25
中国果树(2020年2期)2020-07-25
延边大学学报(自然科学版)(2019年2期)2019-10-08
广西林业科学(2016年2期)2016-03-20
山东医药(2015年38期)2015-12-07
生物加工过程(2015年2期)2015-11-11
食品工业科技(2014年13期)2014-03-11
食品工业科技(2014年7期)2014-03-11
食品科学(2013年24期)2013-03-11
