
遮阴对森林草莓分株生理特性的影响
2023-11-02 08:29侯佳聪邱安然常琳于玮玮王丽娟龙鸿
天津农学院学报 2023年4期
侯佳聪,邱安然,常琳,于玮玮,王丽娟,龙鸿
(天津农学院 园艺园林学院,天津 300392)
森林草莓(Fragaria vescaLinn.)是草莓属的一种野生植物,可以通过匍匐茎进行营养繁殖。在其繁殖季节,森林草莓当年生短缩新茎的叶腋处产生匍匐茎,节部向上生长产生叶和腋芽,向下生长产生不定根,进入土壤后,成为匍匐茎苗,与植株分离后,即可成为新的植株。这种初次产生匍匐茎的植株被称为“母株”,而母株通过匍匐茎产生的与其在基因型上相同的新个体分株被称作“子株”,母株和子株间发生着物质交换[1]。对于匍匐茎相连的植株而言,分株间处于不同的生长环境时为异质性生境,而处于相同生长环境时则为同质性生境。研究表明无性系繁殖的植物能够通过子母株间的物质交换来缓解处于资源短缺分株受到的胁迫,进而增强整个植物的存活和生长[2]。
在植物生长的过程中,光照、水分、温度以及各种矿物质等都是不可缺少的条件,这些条件在时空上常常呈异质性分布[3],非生物胁迫因素中,光是植物生长发育过程的一个重要环境因子,不同光强、光质和光照时间等均会影响植物的生长发育、形态变化和光合特性[4-5]。野生草莓一般分布在林缘或林下等环境中,匍匐茎相连的分株会跨越林缘、林下或者无高草的生境地[6],这样植株就会处于异质性光照的环境。研究表明无匍匐茎相连的植物对不均匀的环境资源分配常是“趋贫特化”[7],如在光照不足时,植株地上部分生长受限,而地下部分不受影响,植物优先将生物量分配给资源“贫乏”的光源捕获器官(地上茎、叶部等)[8];而有匍匐茎相连的植株在光照不足时,子株由于可从母株得到一定的物质补充,所以会将相对多的生物量投向资源“富足”的器官(根系),即子株发生了“趋富特化”[9]。对具匍匐茎的森林草莓进行遮阴和养分处理的试验表明,异质性生境,包括光照,更适合森林草莓的生长,其生物量和分株数明显增加[7]。然而,光照对森林草莓匍匐茎子株生长的影响主要是形态指标观察,对其生理特性报道较少[10]。本文以森林草莓为试验材料,分成半遮阴组(异质性光照)与全遮阴组(同质性光照),模拟森林草莓自然生长于林缘、林下等生境,对比不同遮阴处理子株光合特性及生理指标的变化,探讨森林草莓匍匐茎相连子株最适宜的光照条件,以及在异质性遮阴条件下,子母株间可能的物质交换和对胁迫条件的应答,以期为具有分株的森林草莓逆境生理特性提供试验依据。
1 材料与方法
1.1 试验材料
森林草莓种植于智能光照培养箱(MMM Climacell)中,种植条件:温度25 ℃/18 ℃(白天16 h/晚上8 h),光照强度100 μmol/(m2·s),挑选长势良好的森林草莓一级分株(子株)压入基质(V蛭石∶V草炭∶V珍珠盐=1∶2∶1)中(图1),并去除其他分株,正常光照和水分培育,待子株长出根且长势大小和母株一致时,将材料移出光照培养箱,在自然光照下缓苗2 周。
图1 野生森林草莓相连分株
1.2 试验方法
选择株高一致,至少5 片功能叶的森林草莓为供试材料,利用不同密度的黑色遮阴网设置3个遮阴梯度:30%、50%、70%,每个梯度下分为全遮阴组和半遮阴组,0%遮阴率为对照(CK),全遮阴为子株和母株均处于遮阴下,半遮阴为仅子株处于遮阴下,母株不遮阴(图2),每个处理设置3 个重复。试验期间施充足水分,在自然光照下持续培养约一个月后测定各处理子株的生理指标和光合参数等。
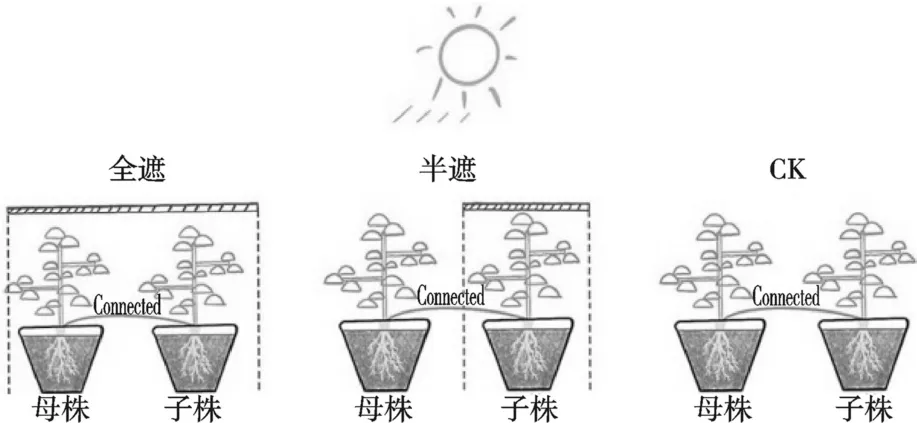
图2 试验设计图解
1.3 测量和数据分析
以各遮阴处理子株的叶片为材料,取0.2 g 叶片,加入石英砂与5 mL 提取液,放入冰浴快速研磨至浆状,并于4 ℃,12 000 r/min 下高速离心20 min,取上清液1.5 mL 为酶粗提取液,放入离心管中并于4 ℃冰箱保存备用,测定SOD 活性和POD 活性参考王奎学[11]方法。取剪碎叶片0.5 g,加入10 mL 的10%三氯乙酸(TCA)及石英砂研磨至匀浆,4 000 r/min 离心10 min,取上清液为提取液,MDA 含量测定参考陈建勋等方法[12]。参考李合生的方法用G-250 考马斯亮蓝测定可溶性蛋白质含量,乙醇提取法测定叶绿素的含量[13]。使用便携式光合测定仪GFS-3000(德国WALZ 公司)按照徐苏婷等的方法测定光合参数[14]。试验数据用Excel 2016 进行数据分析及绘图,采用SPSS 20.0 统计软件进行单因素方差分析,在P值为0.05 的差异显著水平下进行比较。
2 结果与分析
2.1 遮阴对森林草莓子株光合特性的影响
2.1.1 光合色素
在半遮阴处理时,森林草莓子株叶片中的叶绿素a、b 含量随着遮阴率的增高呈现先降低后升高再降低的趋势,在50%的遮阴时,叶绿素a 含量在半遮阴处理组最高,叶绿素b 含量显著高于其他处理(P<0.05),叶绿素a 含量分别是遮阴率为0%、30%、70%的1.04、1.14、1.09 倍,叶绿素b 含量分别是1.62、1.75、1.49 倍。类胡萝卜素含量在不同遮阴率时变化趋势不显著(P>0.05)。在全遮阴处理中,子株叶片中的叶绿素a、b 含量随遮阴率的增高呈逐渐上升趋势,类胡萝卜素含量在50%处理出现谷值,但与其他处理差异不显著(P>0.05),在70%处理时,叶绿素a、b 含量显著高于其他处理(P<0.05)。叶绿素a/b 在50%半遮阴时显著降低,其他处理均无显著差异(P>0.05),全遮阴时叶绿素a 含量和叶绿素a/b 值均高于半遮阴(表1)。
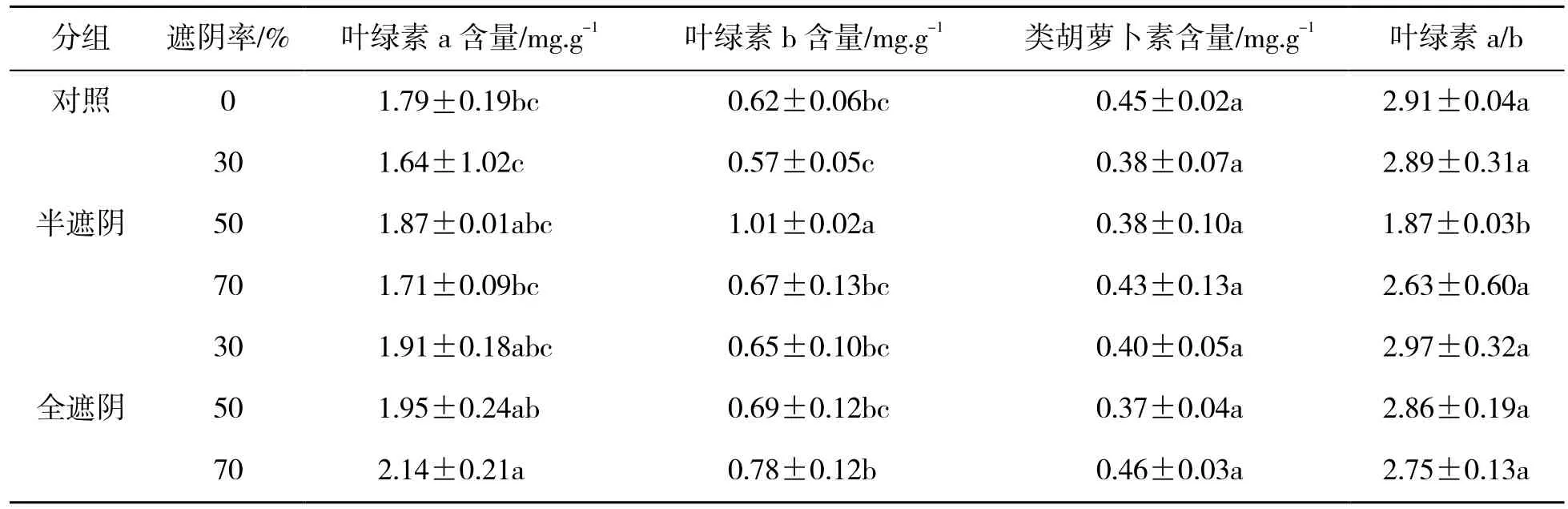
表1 遮阴处理对森林草莓子株光合色素含量的影响
2.1.2 净光合速率
净光合速率(Pn)的日变化主要有三种类型:单峰型、双峰型、三峰型。在半遮阴时,30%遮阴的子株叶片Pn表现为双峰曲线与对照一致,均在12:00 时出现第一个峰值,此时对照和30%半遮阴时的Pn分别为6.79 和5.97 μmol/(m2·s)。随着温度的升高植物气孔关闭,使得森林草莓子株出现光合“午休”现象,在14:00 出现谷值,在下午16:00 时Pn出现第二个峰值,对照与30%处理的值分别为7.69 和4.11 μmol/(m2·s),50%和70%的遮阴处理时均为单峰型,均在10:00 出现不明显的单峰,未出现“午休”现象,对比不同遮阴处理时净光合速率日变化,CK>30%>50%>70%。在全遮阴时,各遮阴处理的子株叶片Pn均表现为单峰型,其中70%处理较其他处理较早出现峰值,在12:00 时叶片Pn达到最高为5.10 μmol/(m2·s),30%和50%处理时Pn在16:00 出现峰值数值分别为9.41 和6.68 μmol/(m2·s),不同遮阴处理的Pn日变化,表现为30%>CK>50%>70%(图3)。30%和50%全遮阴子株叶片Pn均大于同遮阴率的半遮阴处理,而70%全遮阴在10:00 前的Pn低于70%半遮阴处理,但在12:00 进行了超跃。30%全遮阴处理时Pn大于对照,表明当子株和母株处在轻度遮阴30%的同质性光照时,其子株积累同化物的能力高于子、母株处于异质性光照,且高于未遮阴。
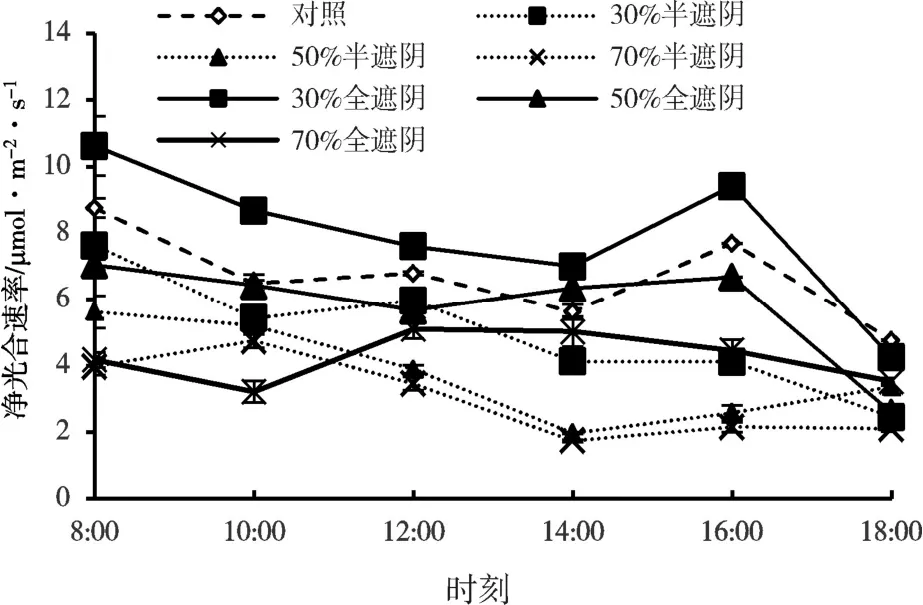
图3 森林草莓半遮阴与全遮阴处理下子株净光合速率的日变化
2.1.3 气孔导度
在半遮阴时,森林草莓的气孔导度(Gs)在30%处理时表现为双峰曲线,与对照趋势一致,出现两个峰值的时间分别是10:00 和16:00 点,在14:00 时出现谷值。50%和70%处理均表现为单峰型。对比不同遮阴处理时叶片Gs日变化表现为CK>30%>50%>70%。在全遮阴时,30%、50%、70%遮阴处理的叶片Gs变化均为单峰型,30%处理在16:00 出现峰值,叶片Gs为129.84 mol/(m2·s),50%和70%处理在14:00 出现峰值,叶片Gs分别为96.20 和62.51 mol/(m2·s)(图4),遮阴处理的Gs均为先降低再升高再降低,而对照有一个升高的过程。对比不同遮阴处理时气孔导度日变化,表现为30%>CK>50%>70%,总体来看,全遮组Gs峰值出现时间比半遮组有所延迟。
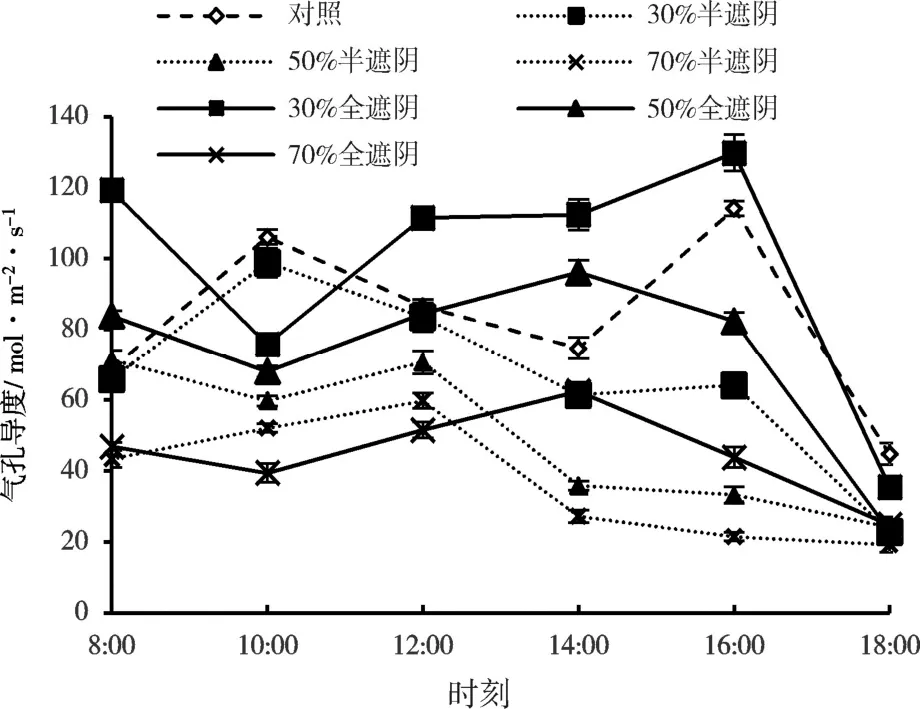
图4 森林草莓半遮阴与全遮阴处理下子株气孔导度的日变化
2.1.4 蒸腾速率
在半遮阴时,各处理子株叶片蒸腾速率(Tr)在一天中均低于对照,且各遮阴处理的Tr日变化规律呈现单峰型,均在10:00 达到最高值,30%、50%和70%处理时叶片Tr分别为4.23、3.55、2.59 mmol/(m2·s)。对照表现为双峰型,在12:00 和16:00 分别出现两次峰值,16:00 时蒸腾速率最高,达到5.35 mmol/(m2·s)。在全遮阴时,各遮阴处理Tr日变化仍呈单峰型,30%与对照一致,在16:00 达到Tr最大值且高于对照,此时30%处理时子株叶片Tr为7.01 mmol/(m2·s)。50%和70%处理在14:00 出现峰值,早于30%处理出现峰值时间,对比不同遮阴处理时叶片Tr日变化为30%>CK>50%>70%(图5)。总体来看,全遮组子株Tr出现峰值时间相比半遮组均有推迟,除全遮组中的30%处理的Tr高于对照外,其余处理均低于对照,说明在子株、母株处于同质性30%遮阴处理时,植株表现出较好的适应性。
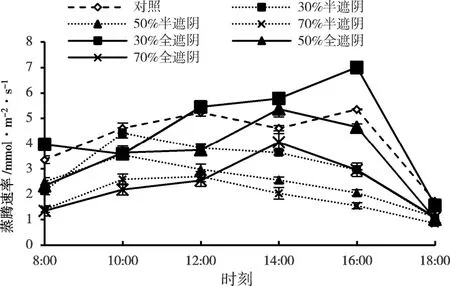
图5 森林草莓半遮阴与全遮阴处理下子株叶片蒸腾速率的日变化
2.2 遮阴对森林草莓子株抗氧化酶的影响
2.2.1 超氧化物歧化酶
随着遮阴率的增加半遮组与全遮组子株的超氧化物歧化酶(SOD)活性均呈先降低后上升的趋势(图6A),均在70%遮阴时的SOD 活性显著高于其他处理(P<0.05),在半遮阴时分别是对照、30%、50%处理的1.15、1.25、1.16 倍,在全遮阴时分别是1.23、1.38、1.09 倍,而70%的全遮阴和半遮阴两个处理的SOD 活性差异并不显著(P>0.05)。在所有处理中,30%的全遮阴和半遮阴两组处理的SOD 活性最低,虽30%全遮阴组稍低于30%半遮阴组,但二者差异并不显著。
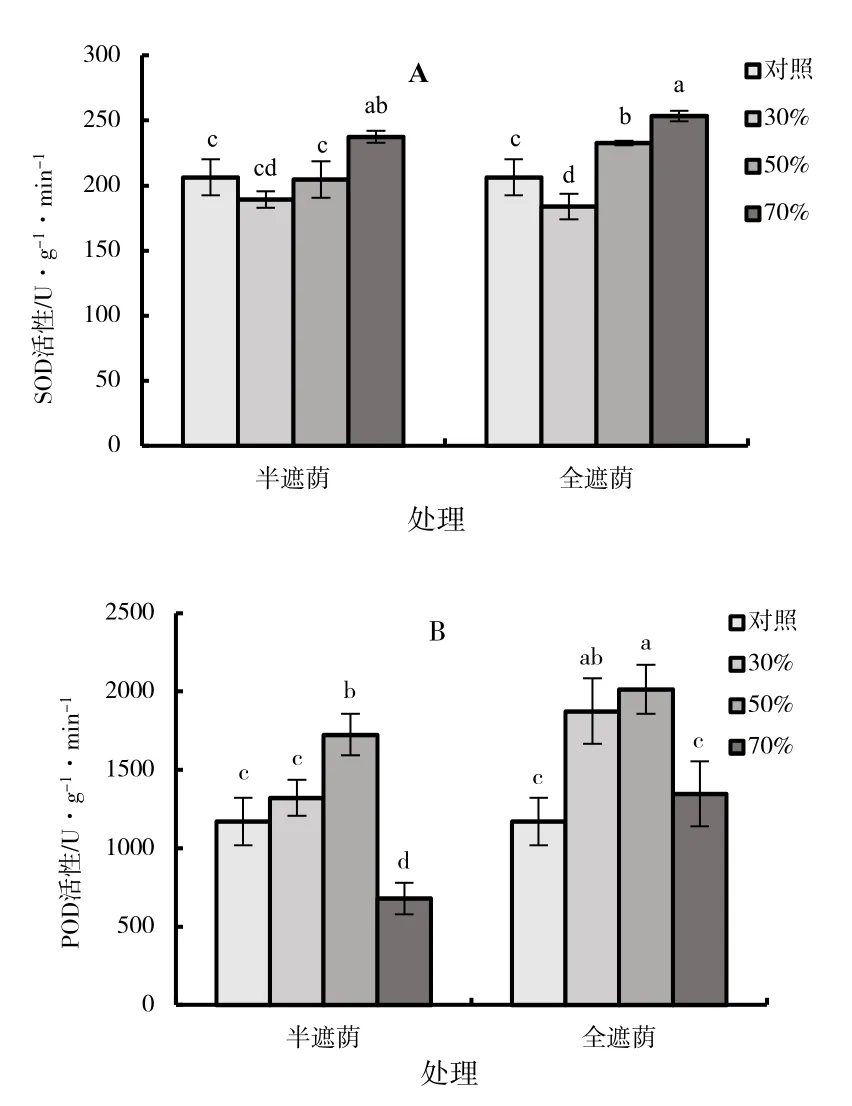
图6 森林草莓半遮阴与全遮阴处理下子株叶片抗氧化酶活性指标
2.2.2 过氧化物酶
过氧化物酶(POD)能够清除植物细胞内过多活性氧,保护植物免受伤害。随着遮阴率的升高,两个处理子株的POD 活性变化趋势一致,均呈先升高后降低(图6B),遮阴率为30%时半遮阴子株的POD 活性与对照差异不显著,而全遮阴则显著升高(P<0.05),在50%遮阴时两个处理组的POD 活性均显著高于各组其他处理(P<0.05),半遮阴组相对于对照增加了47.3%,全遮阴组则增加了72%,在各遮阴处理中全遮阴组的子株POD 活性均高于半遮阴组。
2.3 遮阴对森林草莓子株可溶性蛋白含量的影响
随着遮阴率的增高,全遮阴组与半遮阴组中的子株叶片可溶性蛋白含量呈先升高后降低的趋势(图7),在半遮阴组中,50%处理时可溶性蛋白含量显著高于其他处理(P<0.05),分别是对照、30%、70%处理的1.14、1.16、2.22 倍。在全遮阴组中,30%处理时可溶性蛋白含量显著高于其他处理(P<0.05),分别是对照、50%、70%处理的1.33、1.87、6.17 倍。在70%遮阴处理时,半遮阴和全遮阴子株中叶片可溶性蛋白含量相对于对照分别减少了49.83%和78.40%,全遮阴70%处理的可溶性蛋白含量极显著低于对照及其他处理(P<0.05)。
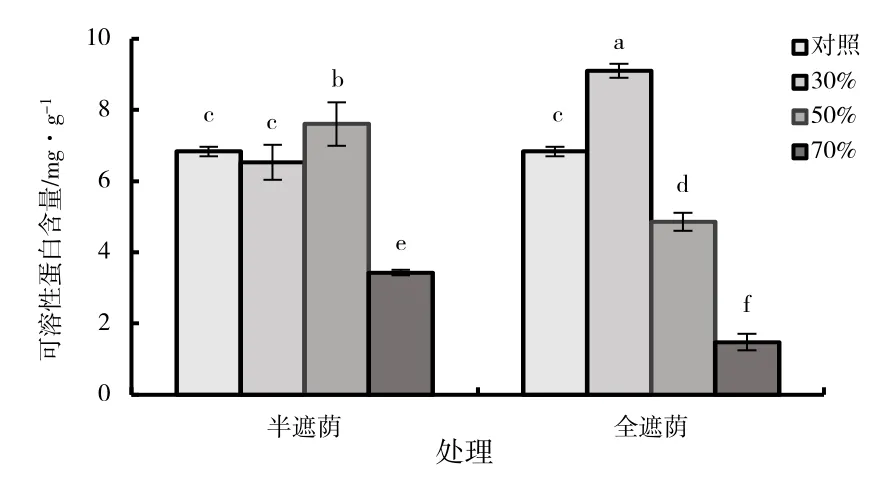
图7 森林草莓半遮阴与全遮阴处理下子株叶片可溶性蛋白含量指标
2.4 遮阴对森林草莓子株丙二醛含量的影响
丙二醛(MDA)的含量是衡量膜脂过氧化损伤的重要指标,半遮阴组与全遮阴组子株的MDA含量随遮阴率的增高呈先升高后下降再升高的趋势(图8)。在30%半遮阴处理的子株MDA 的含量显著高于其他处理(P<0.05),其含量相比对照增加了40.28%。50%半遮阴处理的MDA 含量显著低于对照及其他处理(P<0.05)。在全遮阴组中,30%的全遮阴处理MDA 含量最高,在所有处理仅次于30%半遮阴处理,二者差异显著(P<0.05),但与对照差异不显著(P>0.05);50%全遮阴处理时MDA 含量显著低于对照及全遮阴时的其他处理(P<0.05),半遮阴子株比全遮阴对不同光照反应更敏感,30%半遮阴时MDA 含量显著高于全遮阴处理(P<0.05),而50%和70%半遮阴的子株叶片MDA 含量均显著低于全遮阴(P<0.05)。
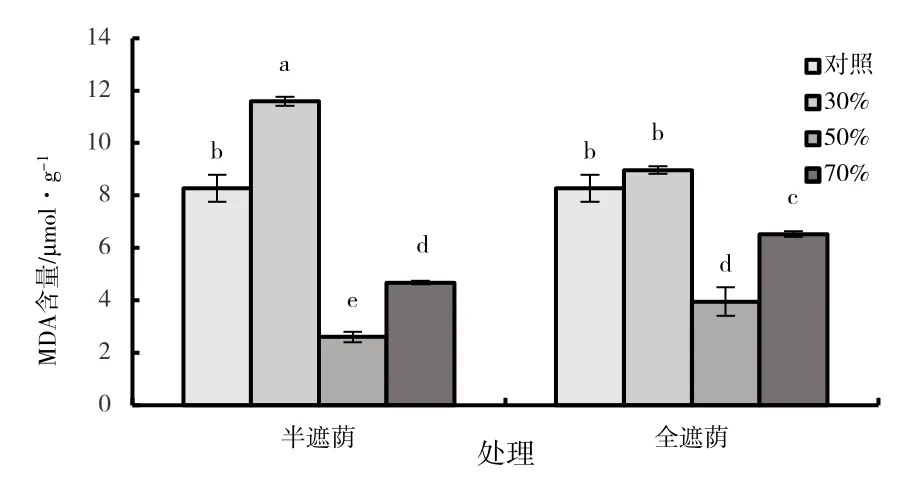
图8 森林草莓半遮阴与全遮阴处理下子株叶片丙二醛含量指标
3 讨论与结论
光照在植物形态建成中具有重要作用,光合作用是植物最基本的生命活动,光可以通过光合作用为植物生长提供能量,也可以作为环境信号因子来调控植物自身的生长发育进程。遮阴是植物自然生境下面临的一种光照强度变化,也可以是农业促成栽培生产实践中调整光照强度的技术手段。研究表明,过强的光照会引起叶绿素的光氧化和分解,遮阴可以提高青钱柳、朱顶红等叶绿素含量[15-16]。本研究结果表明,森林草莓全遮阴子株的光合色素总量高于半遮阴的子株,随遮阴率的增加光合色素总量呈增加趋势,在70%遮阴处理时各指标含量最高,可能是叶片为适应弱光环境的一种反应[17]。此外,遮阴对其他光合作用的生理指标也产生影响。两个草莓品种‘宝交早生’和‘硕丰’遮阴试验表明,经过遮阴处理后,两种草莓在不同光强的光合速率都有所下降[18],咖啡叶片的Pn日变化为双峰曲线,下午的Pn出现峰值仍低于上午[19],本研究中,各处理中的Pn日变化出现的峰值低于上午8:00,与前人结果一致,可能是下午低温、低湿的原因。随着遮阴率增加,森林草莓的Pn、Gs、Tr在同一时间半遮阴时均呈现CK>30%>50%>70%,全遮阴为30%>CK>50%>70%,半遮阴的趋势与前人研究结果一致,而在全遮阴的30%处理最高,这可能是试验材料受到了自然强光的影响,全遮阴30%更适宜子株利用光资源。在相同遮阴率时,全遮阴森林草莓子株叶片的Pn、Gs、Tr在下午均超过了半遮阴,全遮阴的各光合参数日变化出现峰值的时间均比半遮阴组有所推迟,说明母株通过匍匐茎对子株的光合作用产生了影响。
植物体受到逆境胁迫时会产生应激生理反应,产生过量自由基(ROS),导致膜脂过氧化程度增加,其最终产物MDA 含量与植物受伤害的程度呈正比,抗氧化酶和渗透调节物质都会发生变化,SOD 是生物体内重要的抗氧化酶,也是第一道防线,可以清除细胞中多余的O2-、OH-发生歧化反应形成O2和H2O2,POD 为清除H2O2的关键酶[20]。遮阴处理模拟了植物生长的弱光胁迫,研究表明,不同品种的甜瓜叶片遮阴处理后MDA、POD 活性均有所增加[21];此外,随着遮阴程度的增加,辣椒SOD、POD 和CAT 等保护酶活性均呈下降趋势,适度遮阴降低了MDA 的含量[22]。牛轭草及铺地锦竹草随遮阴程度加强,POD 活性先升高后降低,SOD 活性先降低后升,反映出抗氧化酶间的协同作用[23]。本试验中,两处理组的SOD 活性均呈先降低后上升趋势,POD 活性先升后降,与前人结果一致[23],对照和30%遮阴处理的子株受强光照胁迫,保护酶保持较高活性,遮阴后期SOD 活性升高,POD 活性降低,可能是发生弱光胁迫,使得植物体内活性氧上升,植物主要靠激发SOD 活性来增加抗逆能力。张婷等认为,匍枝萎陵菜遮阴处理SOD 活性与MDA 含量存在负相关[24]。本试验中,MDA 的变化在两个处理组中均呈先升高后降低再升高的趋势。在自然光下,CK 和30%遮阴时,强光胁迫对植物体影响占据主导,在30%半遮阴处理时MDA 含量比对照显著增加,此时处于异质性高光照下的母株受到了强光胁迫,而30%遮阴不足缓解强光对子株的影响,母株通过匍匐茎对子株产生了负影响,导致半遮阴子株的MDA含量升高;当母株同处于30%遮阴时,较前者相对缓解了自然强光对整个植株带来的影响,但同CK 依旧受到了自然强光的负影响,因此全遮阴子株的MDA 含量与CK 差异不显著。在50%遮阴处理时,两个处理组MDA 含量均达到最低值,在70%遮阴时两个处理组的POD 活性骤降,WSP 含量低于正常条件,弱光处理(50%~70%)时半遮阴子株的MDA 含量显著低于全遮阴,表明此时处于异质性生境的母株对子株有显著支持。
具有匍匐茎的植物由于形成了子母株,且子母株之间保持着物质和信息的交流。研究发现当智利草莓[25]、芦苇[26]、空心莲子草[27]等带有相连分株的无性系植物生长于异质性生境时,在高养分的分株可以给低养分分株显著支持,实现资源共享。森林草莓和过路黄分株的同质性和异质性光照及养分试验表明,资源异质性条件更适宜克隆片段生物量积累[28];其他匍匐茎植物光照、水分、养分交互斑块性环境试验结果也显示类似生物量分配格局[29]。5 种地被竹的母株或子株遮阴试验也表明子母株资源传递均为双向传递,有效缓解了环境胁迫[30]。本试验结果表明,夏季自然高光胁迫在CK 和30%遮阴占据主导,30%半遮阴子株MDA 含量高于CK,可能是受到了处于光抑制中母株的负影响;随遮阴率增加(>30%),半遮阴子株的MDA 含量显著低于全遮阴组,表明当弱光胁迫占据主导时,处于异质性生境的母株对子株有了显著支持。在30 天的遮阴处理中,同质性30%遮阴处理森林草莓各光合参数均达到了峰值,此时利于森林草莓子株对光资源的利用,50%遮阴时MDA 含量在两个处理组均达到谷值,表明自然光照下适度遮阴适宜森林草莓的生存,可能与森林草莓的耐荫性有关,而重度遮阴(>50%)破坏了森林草莓的生理平衡,影响其正常生长。在自然生境中,子母株各司其职获取资源并相互影响着,任何一方所处环境对于整个植株不利,便会对相连分株产生影响,母株可以正向保护受胁迫的子株,同时在自身受到胁迫时也会反向获取资源,这种双向传递有效缓解了环境胁迫,可能就是匍匐茎分株具有较强生态适应性的原因。
猜你喜欢
中国机械工程(2022年8期)2022-05-09
中国机械工程(2021年8期)2021-05-07
西藏农业科技(2019年2期)2019-07-10
音乐教育与创作(2019年8期)2019-05-16
江苏农业科学(2017年3期)2017-05-02
果树实用技术与信息(2016年8期)2016-09-21
江苏农业科学(2015年8期)2015-09-10
生态学报(2015年4期)2015-03-10
生态学报(2015年23期)2015-02-11
河南科技(2014年4期)2014-02-27
