
新疆棉田蓟马的捕食性天敌优势种类筛选与控害功能评价
2023-10-24 05:41:28党雯雯刘冰褚栋陆宴辉
中国农业科学 2023年17期
党雯雯,刘冰,褚栋,陆宴辉,3
新疆棉田蓟马的捕食性天敌优势种类筛选与控害功能评价
党雯雯1,2,刘冰2,褚栋1,陆宴辉2,3
1青岛农业大学植物医学学院,山东青岛 266109;2中国农业科学院植物保护研究所植物病虫害综合治理全国重点实验室,北京 100193;3中国农业科学院西部农业研究中心,新疆昌吉 831100
【目的】蓟马是新疆棉田一类主要害虫,明确棉田蓟马的优势捕食性天敌种类及其控害功能,可为发展棉田蓟马的生物防治技术提供科学依据。【方法】于2021—2022年采用目测法,通过对田间不同小区进行五点取样,系统调查南疆库尔勒棉田蓟马及捕食性天敌的种群消长动态。同时对棉田不同生育期采集的捕食性天敌样本进行分子检测,先提取整头天敌的DNA,再利用烟蓟马()、花蓟马()特异性引物进行PCR扩增,获取各种天敌体内的蓟马物种信息,据此组建捕食性天敌-蓟马定量食物网,比较棉花不同生育期相关食物网结构的差异。并利用捕食者-猎物功能反应模型,针对不同天敌(包括成虫及幼体),分别设置50、100、200、300和400头/皿的花蓟马猎物密度,室内评价不同天敌对蓟马的捕食功能。【结果】棉花苗期和蕾期田间蓟马发生量偏低,花铃期蓟马密度较高,种群高峰期出现在7月下旬至8月初。从田间种群动态来看,多异瓢虫()在苗期、蕾期和花铃期均为棉田优势捕食性天敌,异须微刺盲蝽()在花铃期后期为主要天敌。分子检测表明,棉花各生育期多异瓢虫个体中蓟马DNA检出率均较高,苗期-蕾期十一星瓢虫()、花铃期后期异须微刺盲蝽中也有较高检出率。室内捕食功能反应表明,不同天敌对花蓟马的捕食能力均随猎物密度的增加而增加,多异瓢虫幼虫、异须微刺盲蝽若虫、普通草蛉()幼虫对花蓟马的捕食功能均符合Holling-Ⅱ模型,而多异瓢虫成虫、异须微刺盲蝽成虫对花蓟马的捕食功能符合Holling-Ⅲ模型,在高猎物密度(400头/皿)下,多异瓢虫成虫的捕食量显著高于其幼虫、异须微刺盲蝽成虫和若虫、以及普通草蛉幼虫。【结论】多异瓢虫在整个生育期均为棉田蓟马的优势捕食性天敌,异须微刺盲蝽为花铃期后期主要捕食性天敌。研究结果可为保育利用捕食性天敌防治新疆棉田蓟马提供科学依据。
棉花;蓟马;多异瓢虫;异须微刺盲蝽;捕食作用;食物网
0 引言
【研究意义】蓟马类害虫通常食性复杂,繁殖能力强,寄主范围广;吸食植物汁液,造成叶片萎蔫,影响植株生长,导致作物产量与品质下降;部分种类还会传播多种植物病毒,造成更加严重的间接危害。蓟马是新疆棉田的一类重要害虫,近年来发生危害逐渐加重。天敌介导的生物控害是农作物害虫综合治理的重要措施[1],明确不同天敌对害虫的控制作用将有助于优化天敌的保育对策,创新发展害虫生物防治技术。因此,了解棉田蓟马及其天敌种群动态,明确不同生育期优势天敌的物种组成,阐明棉田蓟马及其捕食性天敌的食物网结构,评估不同捕食性天敌的生物控害能力,有助于系统解析棉田自然天敌对蓟马的控害服务功能,提升棉田蓟马的综合治理水平。【前人研究进展】国内外研究报道捕食性天敌对一些种类蓟马具有明显控制作用[2],但大多数集中在蔬菜和园林作物上,其中天敌以捕食螨[3-5]和小花蝽[6-8]居多。如美国应用黄瓜新小绥螨()和斯氏钝绥螨()防治辣椒上的茶黄蓟马()[9];在欧洲利用巴氏新小绥螨()和黄瓜新小绥螨防治烟蓟马()和西花蓟马()[10];禹云超等[11]报道了斯氏钝绥螨对西花蓟马和豆大蓟马()的捕食功能符合Holling-Ⅱ模型;王嘉阳等[12]评价了双尾新小绥螨()对花蓟马()的室内捕食功能反应及其在棉田的控害效果,发现双尾新小绥螨对花蓟马的1龄若虫捕食量显著高于2龄若虫,日捕食量最高可达6.1头。捕食性蝽也是重要的天敌昆虫,被广泛应用于蓟马的生物防治[13-15]。如孙英等[16]报道南方小花蝽()对蚕豆上的黄蓟马()成虫和若虫的捕食功能反应符合Holling-Ⅱ模型,日最大捕食量分别为29.3和34.3头,南方小花蝽的发生与黄蓟马呈现一定的跟随关系;胡昌雄等[17-18]研究了辣椒田蓟马及主要捕食性天敌种类,西花蓟马是10种蓟马中优势度最高的物种,而南方小黑花蝽()在捕食性天敌中的优势度最高,捕食性天敌活动高峰时间明显滞后于蓟马;王杰等[19]报道释放东亚小花蝽()对温室辣椒和茄子花内的西花蓟马具有良好的控制作用。部分其他类型的捕食性天敌对蓟马也有较好的控制作用。如六斑月瓢虫()可以捕食黄胸蓟马()若虫,捕食功能反应符合Holling-Ⅱ模型[20]。两种通草蛉和对温室番茄上的西花蓟马有较好防治效果[21]。【本研究切入点】新疆棉田蓟马主要是烟蓟马与花蓟马,棉花苗期以烟蓟马为主[22],花蓟马主要在花铃期危害[23]。新疆是我国最大的产棉区,2022年产量已占全国的90%以上,随着棉花种植规模的不断扩大,蓟马已由偶发性害虫上升为主要害虫。目前,棉田蓟马防治以使用化学农药为主,其优势天敌种类及其控害功能尚未明确,亟需开展系统研究。【拟解决的关键问题】于2021—2022年系统调查南疆库尔勒棉田蓟马及其捕食性天敌的种群消长动态,对棉田捕食性天敌样本进行分子检测,构建捕食性天敌-蓟马食物网进而解析食物关系,通过室内捕食功能反应评价不同天敌对蓟马的捕食能力,系统阐明不同天敌对蓟马的控制作用,为创新发展新疆棉田蓟马的生物防治技术提供科学依据。
1 材料与方法
1.1 棉田蓟马及捕食性天敌种群动态调查
棉田蓟马及天敌种群调查于2021和2022年在中国农业科学院植物保护研究所新疆库尔勒试验基地(85.81°E,41.75°N)开展。供试棉花品种为‘中49’,于每年4月中旬播种,覆膜栽培,单膜宽1.25 m,1膜4行,株距10 cm,行距15 cm,以膜下滴灌方式灌溉,整个生育期不施用任何化学农药。试验共设6个棉花小区,作为6个重复,每个小区面积为700 m2(50 m×14 m),相邻小区之间设置1 m间隔。采用五点取样法,每点选10株棉花,用小吊牌定点定株。每年从6月初棉花3—5叶期开始,至8月底,每7 d 1次采用目测法系统调查各小区所标记棉株上蓟马成/若虫数量、主要捕食性天敌的种类及数量。
1.2 棉田捕食性天敌-蓟马食物网结构分析
在进行棉田蓟马及捕食性天敌种群动态调查的同时,每14 d采集1次各种捕食性天敌昆虫。对于种群密度高的多异瓢虫(),每次采集10—30头左右,而其他发生量少的天敌每种每次采集10头以下。采集后,立即将天敌单头放入装有100%酒精的1.5 ml离心管中,用铅笔写好标签,装入自封袋中,在-20 ℃冰柜中保存。
根据DNA提取试剂盒(天根生化科技(北京)有限公司)说明书步骤提取蓟马和棉田常见昆虫的DNA,用于制作蓟马特异性引物检测模板DNA。烟蓟马特异性SS-COI引物(Ti-5F:5′-ATTATCAACAT TCTATCATTCGGGTA-3′,Ti-5R:5′-AGGATCAAAA AATGAGGTA-3′)参考游中华等[24]。花蓟马引物序列(Fi63-4F:5′-CGCTAAGTATTTTAATTCGG-3′;Fi63- 4R:5′-AGTGGCACTAGTCAGTTTCC-3′)参考李海强等[25]。以对应蓟马物种DNA为模板,对2对引物进行温度梯度PCR扩增试验,得到各引物对的最佳退火温度。以烟蓟马、花蓟马、多异瓢虫、异须微刺盲蝽(s)、黑食蚜盲蝽()、十一星瓢虫()、异色瓢虫()、大草蛉()、普通草蛉()、棉蚜()、绿盲蝽()、三点盲蝽()的DNA作为模板,检测引物特异性。
从贮存离心管中取出田间采集的天敌样本,待酒精挥发后,对整头天敌进行DNA提取,利用烟蓟马、花蓟马特异性引物进行PCR扩增。PCR反应采用25 µL反应体系,其中含2 µL DNA模板,2.5 µL 10×buffer,0.5 µL 10 mmol·L-1dNTP混合液,引物(10 µmol·L-1)各1 µL,TaqDNA聚合酶(2.5 U·µL-1,NEB)0.5 µL。反应条件:94 ℃预变性5 min;30个循环:94 ℃30 s,48 ℃40 s,72 ℃1 min;72 ℃后延伸10 min。将扩增后的所有PCR样品点入6×RNA/DNA Loading Buffer(北京博迈德生物技术有限公司),在琼脂糖凝胶(1.5%)(北京酷来搏科技有限公司)上进行点样;PCR样品每孔点入5.9 µL,以BM2000 Marker(北京博迈德生物技术有限公司)作为参照物点入量为3 µL,电泳缓冲液为1×TAE,设定电压为200 V、电流400 mA进行电泳检测扩增产物。根据凝胶电泳结果,判断天敌是否取食蓟马,获得天敌体内的蓟马物种信息,构建捕食性天敌-蓟马食物网。
1.3 天敌对蓟马的室内捕食功能评价
以花蓟马为代表开展天敌对蓟马的捕食功能评价,测试的捕食性天敌包括多异瓢虫3龄幼虫和2—6日龄成虫、异须微刺盲蝽3龄若虫和2—6日龄成虫、普通草蛉3龄幼虫。花蓟马和各种捕食性天敌均于2022年6月下旬至7月初采自中国农业科学院植物保护研究所新疆库尔勒试验基地棉田。所有天敌昆虫试验前均饥饿24 h。猎物为花蓟马成虫,设置5个密度,分别为50、100、200、300和400头/皿。在直径9 cm塑料培养皿中放置1片大小相近的幼嫩棉花叶,用脱脂棉蘸少许清水包住叶柄保湿,用小毛刷小心接入不同密度的花蓟马成虫和1头已饥饿处理24 h的天敌昆虫,置于(25±1)℃,45%±5% RH室内环境中,24 h后记录各培养皿中花蓟马剩余数量,统计被捕食猎物数量,每个密度均进行5次重复。
1.4 统计分析
通过独立样本T-test检验法分别比较棉花苗期-蕾期(每年5月下旬至7月上旬)、花铃期(7月中旬以后)之间蓟马、捕食性天敌种群密度差异。因两组数据均满足正态性和方差齐性,因此直接用原始数据进行T检验。
针对分子检测数据,绘制棉田捕食性天敌-蓟马食物网结构图,对多异瓢虫、异须微刺盲蝽等天敌体内烟蓟马与花蓟马的检出数量进行卡方检验,分别比较不同时期这两种蓟马检出率的差异。
根据室内所测定不同天敌对花蓟马的捕食情况,绘制捕食量与花蓟马密度关系曲线,利用Holling圆盘方程进行捕食功能反应分析[26]。Holling-Ⅱ型方程式为Na=a'TN(/1+a'ThN),式中,Na为1头捕食者实际消耗的猎物数量,a'为捕食者对猎物的瞬时攻击率,N为设定的猎物密度,Th为捕食者处理单头猎物的时间,试验处理时间T设置为1 d(T=1)。此外,根据捕食反应曲线,对多异瓢虫成虫及异须微刺盲蝽成虫捕食量进行Holling-Ⅲ方程拟合,方程式为Na=a*EXP(-b/N),N为猎物密度,Na为相应密度下的实际捕食量,a为理论最大捕食量,b为捕食者的最佳寻找密度。利用T检验法比较相同猎物密度下捕食性天敌成虫和幼(若)虫对花蓟马捕食量的差异。
所有数据分析均通过R 4.2.1软件完成[27]。
2 结果
2.1 棉田蓟马及捕食性天敌种群动态
由图1-A、1-B可知,两年调查棉田蓟马发生趋势基本一致,从6月初开始,蓟马种群密度逐渐增加,至7月底和8月初棉花花铃期达到高峰期,2021和2022年高峰期蓟马平均密度分别为617和1 287头/50株,随后种群开始消退,8月底至9月初随着花期结束田间种群基本消退。统计分析表明,花铃期蓟马种群密度显著高于苗期-蕾期(2021:t=16.5,df=10,<0.001;2022:t=9.4,df=10,<0.001)。
由图1-C、1-D可知,多异瓢虫和异须微刺盲蝽的种群密度在两年各调查时期的占比均超过棉田捕食性天敌调查总量的97%,表明这两种天敌为棉田优势捕食性天敌。从种群数量上看,多异瓢虫从棉花苗期开始,至花铃期均为棉田优势捕食性天敌,发生高峰期主要集中在6月下旬和7月上中旬。其中,2021年多异瓢虫种群密度至7月中旬盛花期达到最高峰,峰值密度为97头/50株,随后种群密度开始下降,至8月底田间种群基本消退,苗期-蕾期虫口密度显著低于花铃期(t=-4.3,df=10,<0.001)。2022年棉田多异瓢虫发生早于2021年,6月底种群密度达到高峰,峰值密度为283头/50株,该年度苗期-蕾期多异瓢虫种群密度显著高于花铃期(t=21.5,df=10,<0.001)。此后,随着棉花进入结铃期,多异瓢虫种群数量快速消退,至8月底几乎全部消失。花铃期后期,异须微刺盲蝽种群密度快速增加,逐渐成为棉田主要捕食性天敌,2021和2022年异须微刺盲蝽高峰期虫量分别为68和91头/50株,其种群密度显著高于苗期-蕾期(2021:t=16.1,df=10,<0.001;2022:t=13.6,df=10,<0.001)。
从种群密度可知,多异瓢虫在整个生育期均为棉田优势捕食性天敌,异须微刺盲蝽为花铃期后期主要捕食性天敌。
2.2 棉田捕食性天敌-蓟马食物网结构
两年结果表明,在棉花各生长期田间所采集到的捕食性天敌中以多异瓢虫样本最多,其他几种天敌的发生及采集数量偏低(表1)。在棉花苗期-蕾期,2021和2022年分别采集多异瓢虫146和360头,此外还采集到少量十一星瓢虫。2021年DNA检测发现多异瓢虫体内烟蓟马和花蓟马的检出率分别为75.3%和92.5%,对应两种蓟马的组成比例分别为44.9%和55.1%,二者无显著差异(2=2.55,df=1,=0.110)(图2-A)。2022年多异瓢虫体内两种蓟马的检出率低于2021年,分别为31.9%和26.9%,对应两种蓟马的组成比例分别为54.2%和45.8%,二者也无显著差异(2=1.53,df=1,=0.216)(图2-B)。在花铃期,多异瓢虫两年采集数量分别为446和232头,其体内烟蓟马和花蓟马的检出率分别为41.0%和83.6%(2021年)、23.3%和51.7%(2022年),对应两种蓟马组成比例分别为32.9%和67.1%(2021年)、31.0%和69.0%(2022年),花蓟马所占比例约为烟蓟马的2倍,二者之间差异显著(2021:2=64.93,df=1,<0.001;2022:2=25.03,df=1,<0.001)(图2-C、2-D)。苗期-蕾期与花铃期相比,多异瓢虫体内烟蓟马和花蓟马的物种组成有显著差异(2021:2=10.02,df=1,=0.002;2022:2=19.99,df=1,<0.001)。此外,2022年苗期-蕾期共采集到27头十一星瓢虫,体内检出烟蓟马和花蓟马的概率分别为77.8%和66.7%,对应这两种蓟马组成比例分别为53.8%和46.2%,二者无显著差异(2=0.23,df=1,0.631)(图2-B)。
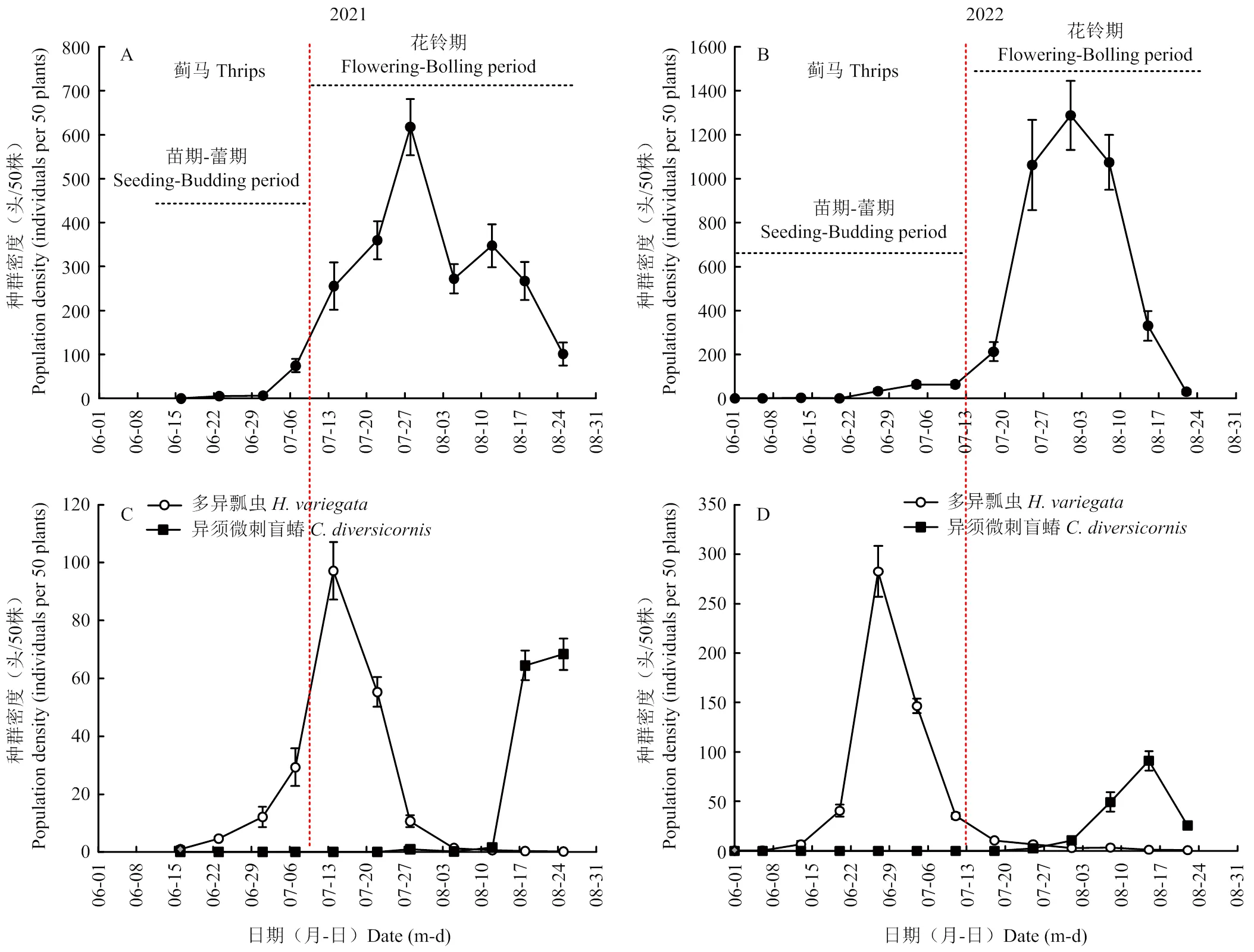
A、B:蓟马种群动态the population dynamic of thrips;C、D:捕食性天敌种群动态The population dynamic of predators
另外,在花铃期,2021年棉田共采集异须微刺盲蝽119头,其体内没有检出烟蓟马,但有47.9%的个体检出了花蓟马(表1、图2-C),这两种蓟马检出比例差异显著(2=57.00,df=1,<0.001)。2022年共采集到51头异须微刺盲蝽,烟蓟马和花蓟马的检出率分别为23.5%和54.9%,对应蓟马组成比例分别为30.0%和70.0%,花蓟马检出率超过烟蓟马的2倍(2=6.40,df=1,=0.011)(图2-D)。2022年在花铃期还采集到极少量的普通草蛉和黑食蚜盲蝽,体内均检测出了花蓟马的DNA,但捕食比例要低于多异瓢虫和异须微刺盲蝽(图2-D)。
在上述结果的基础上,构建了捕食性天敌-蓟马食物网结构图(图2),全面展示了苗期-蕾期、花铃期不同捕食性天敌的种群密度及对烟蓟马、花蓟马的捕食检测比率。从猎物检出率可知,从棉花苗期-蕾期至花铃期,多异瓢虫均为棉田蓟马的重要捕食性天敌,棉花苗期-蕾期十一星瓢虫、花铃期后期异须微刺盲蝽也为重要天敌。
2.3 天敌对蓟马的室内捕食功能反应
图3显示了不同天敌昆虫对花蓟马的捕食量与猎物密度之间关系,根据捕食量-猎物密度曲线,发现天敌对花蓟马的捕食量均随猎物密度增加而增加。对多异瓢虫幼虫、异须微刺盲蝽若虫、普通草蛉幼虫用Holling-Ⅱ圆盘方程进行模拟,对多异瓢虫成虫及异须微刺盲蝽成虫用Holling-Ⅲ方程进行模拟,拟合结果均较好。说明这3种捕食性天敌对花蓟马均具有较好的控制作用。Holling-Ⅱ模型拟合结果表明,多异瓢虫幼虫对花蓟马的瞬时攻击率最高,为1.2069(图3-A),其次为普通草蛉幼虫和异须微刺盲蝽若虫,对花蓟马的瞬时攻击率分别为0.7655(图3-C)和0.4093(图3-B)。Holling-Ⅲ模型拟合结果表明多异瓢虫成虫对花蓟马的理论日最大捕食量为436.78头(图3-A),高于异须微刺盲蝽成虫(其理论日最大捕食量为239.28头,图3-B)。多异瓢虫成虫和异须微刺盲蝽成虫对花蓟马的最佳寻找密度分别为262.45和212.29头。

表1 棉花各生育期用于DNA分析的捕食性天敌样本数量及两种蓟马的检出率
“—”:未采集到该种天敌No predator had been sampled
各种天敌的捕食量均随猎物密度增加而增加,但在相同猎物密度下,多异瓢虫成虫与幼虫对花蓟马的捕食量之间有明显变化(图3-A)。在较低花蓟马密度下,如50和100头/皿,多异瓢虫成虫捕食量略低于幼虫,但无显著差异(50头/皿:t=-1.46,df=8,=0.183;100头/皿:t=-0.54,df=8,=0.605);随着猎物密度的增加,成虫对花蓟马的捕食量快速增加,花蓟马密度超过200头/皿时,成虫捕食量增速逐渐超过幼虫,但差异仍不显著(200头/皿:t=0.10,df=8,=0.925;300头/皿:t=1.75,df=8,=0.118);而400头/皿密度下,成虫捕食量显著高于幼虫(t=16.90,df=8,<0.001)。
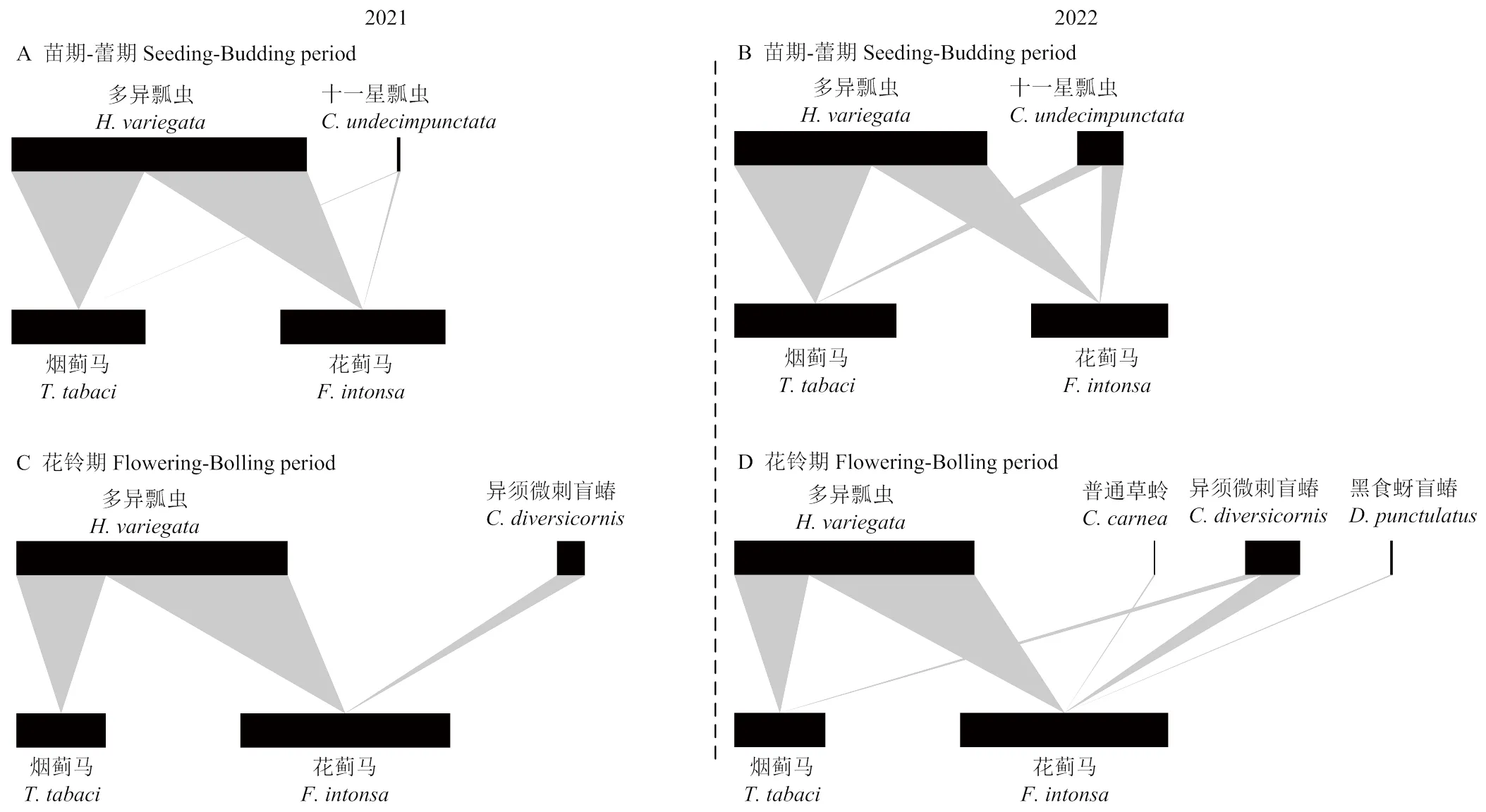
A—B、C—D分别代表2021和2022年棉花苗期-蕾期、花铃期捕食性天敌-蓟马食物网。上层为捕食者,底层为两种蓟马,连接上下两营养层间的灰色三角形区域宽度反映每种捕食者体内检测出的蓟马比例差异。上层黑色条带长度反映各时期采集到的不同捕食者数量差异
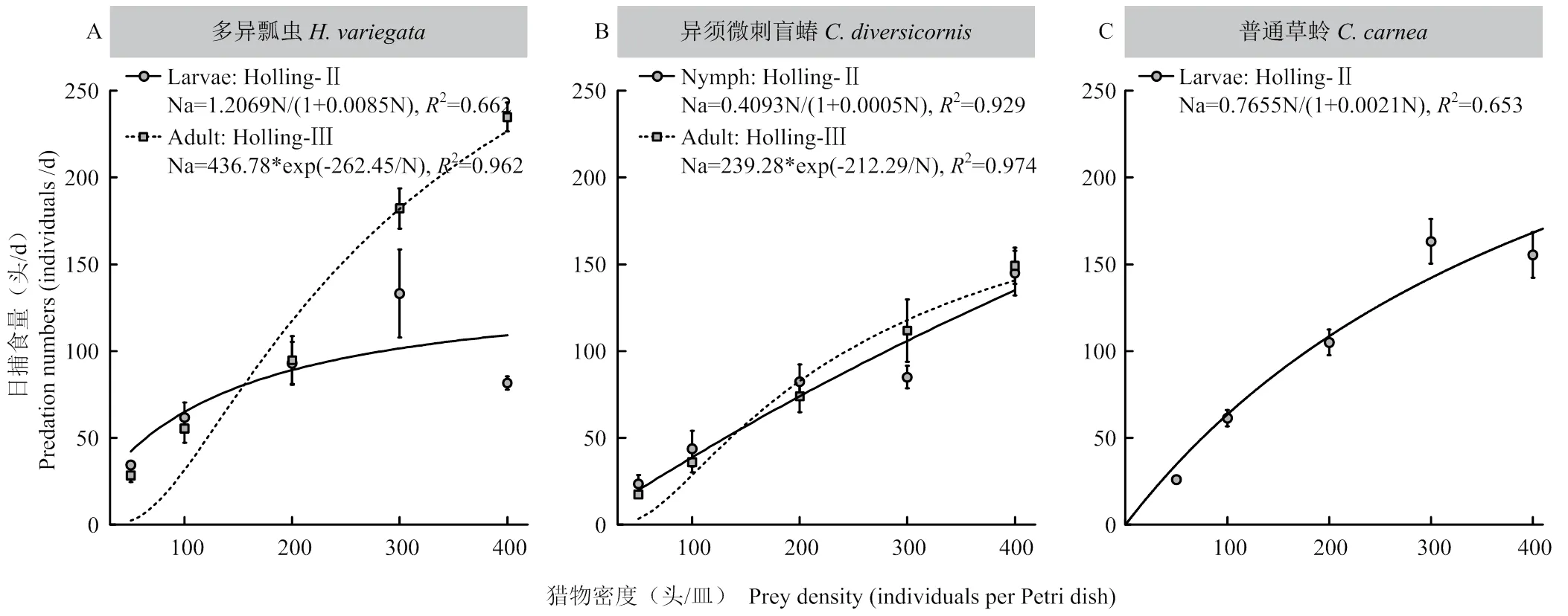
图中实线和虚线分别代表Holling-Ⅱ和Holling-Ⅲ模型拟合曲线,实线代表幼虫(若虫),虚线代表成虫;数据点代表该猎物密度下的捕食量(平均值±标准误)
随着花蓟马密度的增加,异须微刺盲蝽对其捕食量也随之增加。在相同猎物密度下,异须微刺盲蝽成虫和若虫的捕食量无显著差异(50头/皿:t=-1.17,df=8,=0.276;100头/皿:t=-0.66,df=8,=0.527;200头/皿:t=-0.63,df=8,=0.549;300头/皿:t=1.41,df=8,=0.196;400头/皿:t=0.255,df=8,=0.806)(图3-B)。
在相同猎物密度下,特别是猎物密度高时(如400头/皿),多异瓢虫成虫捕食性量显著高于其幼虫、异须微刺盲蝽的成虫和若虫、普通草蛉的幼虫。从捕食量可知,多异瓢虫对棉田蓟马的控制潜力最大。
3 讨论
3.1 棉田蓟马及捕食性天敌的种群消长动态
天敌是控制农田害虫的关键生物因子,掌握害虫及天敌在田间的种群消长动态,阐明其发生和控害的关键时期,有助于解析农作物害虫群落演替规律及其生态学机制。本研究通过两年试验明确了新疆南疆棉田蓟马及其主要捕食性天敌的发生消长动态。从棉花苗期开始,蓟马即可危害棉花,高峰期出现在7月底和8月初,此时棉花处于盛花期,大量蓟马栖息在花里取食花粉和花蜜,此后,随着棉株逐渐开始结铃吐絮,蓟马种群缓慢消退,至8月底9月初随着花期结束田间蓟马种群基本完全消退,花铃期蓟马种群密度显著高于苗期-蕾期。天敌昆虫的发生与猎物常有明显的跟随关系[28]。多异瓢虫食性广,除蓟马外,还捕食蚜虫、叶螨等多种昆虫。棉花苗期、蕾期,田间蚜虫密度高,为多异瓢虫的主要猎物[29-30],而蓟马的种群高峰集中在花铃期,因此棉田多异瓢虫的发生高峰期出现在6月底和7月上旬,明显早于蓟马。
3.2 棉田捕食性天敌-蓟马的定量食物网结构
在农田生态系统中,处于较高营养层级的捕食者与低营养层级的害虫组成了食物网。通过解析食物网结构,可以评价不同营养层级物种间的相互作用,阐明害虫所对应的天敌种类及天敌对害虫的控制功能[31]。DNA分子检测技术的进步,显著促进了节肢动物食物网复杂结构的系统解析,有效实现了食物网结构分析由定性到定量的提升,推动了天敌昆虫在害虫防治中的科学利用[32]。本文对棉田采集的捕食性天敌进行了分子检测,组建了捕食性天敌-蓟马食物网。发现在棉花苗期-蕾期,多异瓢虫是烟蓟马和花蓟马的主要捕食性天敌,十一星瓢虫的发生量较低,其捕食比例低于多异瓢虫,对蓟马控制作用有限。在花铃期,除多异瓢虫外,异须微刺盲蝽体内两种蓟马特别是花蓟马的检出率较高,是重要的天敌种类。另外,尽管普通草蛉和黑食蚜盲蝽发生量比较少,但其体内也检出花蓟马。年度间比较发现,2021年蓟马发生密度显著低于2022年,但2021年多异瓢虫体内蓟马DNA检出率明显偏高。田间调查发现,2022年棉田蚜虫发生量显著高于2021年,因此多异瓢虫取食蚜虫的机会增加,而捕食蓟马等其他猎物的机率随之降低,反之亦然。两年棉田蚜虫发生密度变化,可能是导致蓟马高密度发生的2022年多异瓢虫体内的蓟马DNA检出率低于2021年的主要原因。
3.3 捕食性天敌对蓟马的控制效果
天敌对害虫提供自上而下的控制作用,对于农田害虫治理意义重大,明确不同捕食者对猎物的功能反应,深入挖掘本地天敌资源,充分发挥天敌的生防控害潜能,是实现害虫绿色防控的关键所在[33]。室内捕食功能反应是评估天敌控害能力的常用方法,除捕食螨以外[3],也常利用食虫蝽防治蓟马[2,6]。如生产上利用东亚小花蝽防治西花蓟马[19,34],而淡翅小花蝽()对豆大蓟马具有较好的控制作用[35]。本研究发现,棉田捕食性天敌对花蓟马具有较好的控制作用,随着猎物密度增加,天敌捕食量也随之加大,多异瓢虫幼虫、异须微刺盲蝽若虫、普通草蛉幼虫对花蓟马的捕食功能反应均符合Holling-Ⅱ模型,但多异瓢虫成虫和异须微刺盲蝽成虫对花蓟马的捕食量曲线属于S型,更符合Holling-Ⅲ模型。在相同花蓟马密度,特别是较高猎物密度下,如400头/皿,多异瓢虫成虫对花蓟马的捕食量最大。
3.4 棉田捕食性天敌防治蓟马的应用潜力
通过田间种群动态、室内捕食能力评价、天敌体内猎物检出率3个层面的系统研究,证实多异瓢虫从苗期、蕾期到花铃期,均为棉田蓟马类害虫的优势捕食性天敌,花铃期后期异须微刺盲蝽为蓟马的重要捕食性天敌。本文明确了新疆棉田蓟马及其捕食性天敌的发生规律,阐明了以多异瓢虫、异须微刺盲蝽为代表的棉田优势捕食性天敌对蓟马的控制作用,为进一步挖掘和利用天敌资源,提升对棉田蓟马类害虫的绿色防控提供了科学依据。
在农田害虫防治实践中,常通过设计天敌保育技术对策,提升天敌对害虫的生物防治服务[36]。可种植功能植物来涵养田间天敌,加强对害虫的生态调控[37-38],例如甘草和棉花邻作能提升棉田多异瓢虫等捕食性天敌的数量和生物控害作用[29],大麦和洋葱间作能显著增加洋葱田里的食蚜蝇丰富度及对烟蓟马的控制效果[39]。结合本研究结果,建议重点发展以农田生态调控、选择性杀虫剂使用等措施为主的多异瓢虫和异须微刺盲蝽种群保育技术,提升天敌生物控害服务功能,促进新疆棉田蓟马的绿色防控。
4 结论
新疆棉田蓟马高峰期出现在7月下旬至8月初的花铃期。多异瓢虫在棉花整个生育期均为优势捕食性天敌,对蓟马具有较强的控制作用,异须微刺盲蝽是花铃期后期棉田蓟马的重要捕食性天敌。生产上,应重点保育利用这两种捕食性天敌防控棉花蓟马。
[1] 陈学新, 任顺祥, 张帆, 彩万志, 曾凡荣, 张文庆. 天敌昆虫控害机制与可持续利用. 应用昆虫学报, 2013, 50(1): 9-18.
CHEN X X, REN S X, ZHANG F, CAI W Z, ZENG F R, ZHANG W Q. Mechanism of pest management by natural enemies and their sustainable utilization. Chinese Journal of Applied Entomology, 2013, 50(1): 9-18. (in Chinese)
[2] 杨磊, 邵雨, 李芬, 陈德鑫, 李方友, 吴少英. 缨翅目害虫蓟马生物防治的研究进展. 中国生物防治学报, 2021, 37(3): 393-405.
YANG L, SHAO Y, LI F, CHEN D X, LI F Y, WU S Y. Advances on biological control of thrips pests. Chinese Journal of Biological Control, 2021, 37(3): 393-405. (in Chinese)
[3] 黄建华, 陈洪凡, 王丽思, 柳岸峰, 沈华喜. 应用捕食螨防治蓟马研究进展. 中国生物防治学报, 2016, 32(1): 119-124.
HUANG J H, CHEN H F, WANG L S, LIU A F, SHEN H X. Advances in controlling thrips using predatory mites. Chinese Journal of Biological Control, 2016, 32(1): 119-124. (in Chinese)
[4] VAN DRIESCHE R G, LYON S, STANEK E J, XU B, NUNN C. Evaluation of efficacy offor control of western flower thrips in spring bedding crops. Biological Control, 2006, 36(2): 203-215.
[5] 黄建华, 罗任华, 秦文婧, 黄水金, 秦厚国, 付志飞. 巴氏钝绥螨对芦笋上烟蓟马捕食效能研究. 中国生物防治学报, 2012, 28(3): 353-359.
HUANG J H, LUO R H, QIN W J, HUANG S J, QIN H G, FU Z F. Predation efficacy ofon asparagus thrips,. Chinese Journal of Biological Control, 2012, 28(3): 353-359. (in Chinese)
[6] 刘梅, 张昌容, 尚小丽, 郭军, 曾广, 石乐娟. 南方小花蝽对非洲菊上西花蓟马控制效果评价. 中国生物防治学报, 2020, 36(6): 992-996.
LIU M, ZHANG C R, SHANG X L, GUO J, ZENG G, SHI L J. Control ofonby. Chinese Journal of Biological Control, 2020, 36(6): 992-996. (in Chinese)
[7] HERRICK N J, CLOYD R A, CONNER M A, MOTOLAI G. Insidious flower bug,(Say) (Hemiptera: Anthocoridae), predation on western flower thrips,(Pergande) (Thysanoptera: Thripidae), on Transvaal daisy,, cut flowers and chrysanthemum,×, plants under laboratory and greenhouse conditions. Biological Control, 2021, 163: 104739.
[8] CHOW A, CHAU A, HEINZ K M. Compatibility of()(Athias-Henriot) (Acari: Phytoseiidae) and(Hemiptera: Anthocoridae) for biological control of(Thysanoptera: Thripidae) on roses. Biological Control, 2010, 53(2): 188-196.
[9] ARTHURS S, MCKENZIE C L, CHEN J, DOGRAMACI M, BRENNAN M, HOUBEN K, OSBORNE L. Evaluation ofand(Acari: Phytoseiidae) as biological control agents of chilli thrips,(Thysanoptera: Thripidae) on pepper. Biological Control, 2009, 49(1): 91-96.
[10] SHIPP J L, WHITFIELD G H. Functional response of the predatory mite,(Acari: Phytoseiidae), on western flower thrips,(Thysanoptera: Thripidae). Environmental Entomology, 1991, 20(2): 694-699.
[11] 禹云超, 郅军锐, 曾广, 岳文波, 叶茂. 斯氏钝绥螨对西花蓟马和豆大蓟马若虫的捕食功能反应. 应用昆虫学报, 2019, 56(6): 1317-1323.
YU Y C, ZHI J R, ZENG G, YUE W B, YE M. The functional predatory response oftoandnymphs. Chinese Journal of Applied Entomology, 2019, 56(6): 1317-1323. (in Chinese)
[12] 王嘉阳, 蓝碧云, 方晨, 胡恒笑, 杨瑞彤, 母凯琴, 苏杰, 陆宴辉, 张建萍. 双尾新小绥螨对花蓟马的捕食控害评价. 中国生物防治学报, 2023, 39(2): 271-279.
WANG J Y, LAN B Y, FANG C, HU H X, YANG R T, MU K Q, SU J, LU Y H, ZHANG J P. Predation and control ofby. Chinese Journal of Biological Control, 2023, 39(2): 271-279. (in Chinese)
[13] MAGALHãES S, VAN RIJN P C J, MONTSERRAT M, PALLINI A, SABELIS M W. Population dynamics of thrips prey and their mite predators in a refuge. Oecologia, 2007, 150(4): 557-568.
[14] BOSCO L, GIACOMETTO E, TAVELLA L. Colonization and predation of thrips (Thysanoptera: Thripidae) byspp. (Heteroptera: Anthocoridae) in sweet pepper greenhouses in Northwest Italy. Biological Control, 2008, 44(3): 331-340.
[15] 余金咏, 吴伟坚, 梁广文. 不同食物对杂食性中华微刺盲蝽生长发育及繁殖力的影响. 中国生物防治学报, 2015, 31(4): 495-500.
YU J Y, WU W J, LIANG G W. Effects of food on development, survivorship and fertility of the omnivorous mirid bugSchuh. Chinese Journal of Biological Control, 2015, 31(4): 495-500. (in Chinese)
[16] 孙英, 胡昌雄, 吴道慧, 陈国华, 殷红慧, 徐天养, 陈婷, 张晓明. 南方小花蝽和黄蓟马的种群动态及捕食功能反应. 中国生物防治学报, 2021, 37(3): 451-458.
SUN Y, HU C X, WU D H, CHEN G H, YIN H H, XU T Y, CHEN T, ZHANG X M. Population dynamics ofandand predatory function response. Chinese Journal of Biological Control, 2021, 37(3): 451-458. (in Chinese)
[17] 胡昌雄, 李宜儒, 吕布典, 殷红慧, 徐天养, 胡加云, 陈国华, 张晓明. 辣椒田蓟马及主要捕食性天敌昆虫种类与时间生态位. 植物保护, 2022, 48(5): 38-46, 74.
HU C X, LI Y R, LÜ B D, YIN H H, XU T Y, HU J Y, CHEN G H, ZHANG X M. Species and temporal niches of thrips and their main predatory insects infields. Plant Protection, 2022, 48(5): 38-46, 74. (in Chinese)
[18] 胡昌雄, 范苇, 张倩, 陈国华, 殷红慧, 徐天养, 杨进波, 杨航, 吴道慧, 张晓明. 基于两性生命表和年龄-阶段捕食率的南方小花蝽对西花蓟马的控制作用. 中国农业科学, 2021, 54(13): 2769-2780. doi: 10.3864/j.issn.0578-1752.2021.13.007.
HU C X, FAN W, ZHANG Q, CHEN G H, YIN H H, XU T Y, YANG J B, YANG H, WU D H, ZHANG X M. Control effect ofonbased on the two-sex life table and the age-stage-specific predation rate. Scientia Agricultura Sinica, 2021, 54(13): 2769-2780. doi: 10.3864/j.issn.0578-1752.2021.13.007. (in Chinese)
[19] 王杰, 张晨, 朱正阳, 刘俊秀, 王甦, 徐庆宣, 邸宁. 东亚小花蝽对温室辣椒及茄子花内西花蓟马的控害作用. 中国生物防治学报, 2023, 39(2): 264-270.
WANG J, ZHANG C, ZHU Z Y, LIU J X, WANG S, XU Q X, DI N. Control effects ofonin pepper and eggplant flowers in greenhouses. Chinese Journal of Biological Control, 2023, 39(2): 264-270. (in Chinese)
[20] 李善光, 付步礼, 邱海燕, 杨石有, 马晓彤, 周世豪, 唐良德, 张方平, 刘奎. 六斑月瓢虫对黄胸蓟马若虫的室内捕食作用研究. 应用昆虫学报, 2020, 57(5): 1173-1180.
LI S G, FU B L, QIU H Y, YANG S Y, MA X T, ZHOU S H, TANG L D, ZHANG F P, LIU K. The predation of(Coleoptera: Coccinellidae) to(Thysanoptera: Thripoidae) in the laboratory. Chinese Journal of Applied Entomology, 2020, 57(5): 1173-1180. (in Chinese)
[21] LUNA-ESPINO H M, JIMéNEZ-PéREZ A, CASTREJóN-GóMEZ V R. Assessment of(Banks) and(Hagen) as biological control agents of(Pergande) (Thysanoptera: Thripidae) on tomato () under glasshouse conditions. Insects, 2020, 11(2): 87.
[22] 杨海峰, 王惠珍, 马祁. 棉田烟蓟马的发生与防治. 新疆农业科学, 1985(3): 24-25.
YANG H F, WANG H Z, MA Q. The occurrence and control ofin cotton field. Xinjiang Agricultural Sciences, 1985(3): 24-25. (in Chinese)
[23] 李国英. 新疆棉花病虫害及其防治. 北京: 中国农业出版社, 2017: 207-211.
LI G Y. Pest control of Xinjiang cotton. Beijing: China Agriculture Press, 2017: 207-211. (in Chinese)
[24] 游中华, 路虹, 张宪省, 冯纪年, 石宝才, 宫亚军, 黄大卫. 入侵害虫西花蓟马及其他8种常见蓟马的分子鉴定. 昆虫学报, 2007, 50(7): 720-726.
YOU Z H, LU H, ZHANG X S, FENG J N, SHI B C, GONG Y J, HUANG D W. Molecular identification of the introduced western flower thrips,(Pergande) and other eight common thrips species (Thysanoptera: Thripidae). Acta Entomologica Sinica, 2007, 50(7): 720-726. (in Chinese)
[25] 李海强, 王冬梅, 刘建, 潘洪生, 丁瑞丰, 樊国全, 徐遥, 阿克旦·吾外士, 李号宾. 一种基于特异性SS-COI引物的花蓟马PCR快速检测方法[P]: ZL201610609787.0. (2020-04-07)[2023-05-23].
LI H Q, WANG D M, LIU J, PAN H S, DING R F, FAN G Q, XU Y, WUWAISHI·Akedan, LI H B. A rapid PCR method for detection ofbased on specific SS-COI primers[P]: ZL201610609787.0. (2020-04-07)[2023-05-23]. (in Chinese)
[26] HOLLING C S. Some characteristics of simple types of predation and parasitism. The Canadian Entomologist, 1959, 91: 385-398.
[27] R development core team. R: A language and environment for statistical computing, Version 4.2.1. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org, 2022.
[28] Pan H s, Liu B, Jaworski C C, Yang L, Liu Y q, Desneux N, Thomine E, Lu Y h. Effects of aphid density and plant taxa on predatory ladybeetle abundance at field and landscape scales. Insects, 2020, 11: 695.
[29] XUE Z X, PENG T X, LIU B, LIU Y T, ZHANG Z J, WYCKHUYS K A G, WANG P L, LU Y H. Licorice strips enhance predator- mediated biological control in China’s cotton crop. Pest Management Science, 2023, 79(2): 781-791.
[30] Li J h, Wu Y k, Zhang Q, Li H q, Pan H s, Lu W, Wang D m, Zhang J p, Lu Y h. Aphid parasitism and parasitoid diversity in cotton fields in Xinjiang, China. PLoS ONE, 2018, 13(11): e0207034.
[31] 刘冰, 陆宴辉. 农田节肢动物食物网结构与天敌控害功能. 植物保护学报, 2022, 49(1): 97-109.
LIU B, LU Y H. Arthropod food web structure and the biocontrol services of natural enemies in agro-ecosystems. Journal of Plant Protection, 2022, 49(1): 97-109. (in Chinese)
[32] 杨帆, 刘冰, 陆宴辉. DNA分子检测技术在节肢动物食物网结构解析中的应用. 植物保护学报, 2022, 49(1): 110-117.
YANG F, LIU B, LU Y H. Application of DNA-based molecular detection techniques in arthropod food web structure analyses. Journal of Plant Protection, 2022, 49(1): 110-117. (in Chinese)
[33] WYCKHUYS K A, LU Y, MORALES H, VAZQUEZ L L, LEGASPI J C, ELIOPOULOS P A, HERNANDEZ L M. Current status and potential of conservation biological control for agriculture in the developing world. Biological Control, 2013, 65(1): 152-167.
[34] 张安盛, 于毅, 李丽莉, 张思聪. 东亚小花蝽()成虫对入侵害虫西花蓟马()成虫的捕食作用. 生态学报, 2007, 27(5): 1903-1909.
ZHANG A S, YU Y, LI L L, ZHANG S C. Predation ofadult on adults of western flower thrips (), an invasive insect pest. Acta Ecologica Sinica, 2007, 27(5): 1903-1909. (in Chinese)
[35] 邱海燕, 付步礼, 何石兰, 刘奎. 淡翅小花蝽对豆大蓟马的捕食功能反应及猎物偏好性. 中国生物防治学报, 2022, 38(6): 1443-1448.
QIU H Y, FU B L, HE S L, LIU K. Functional response and predation preference ofto. Chinese Journal of Biological Control, 2022, 38(6): 1443-1448. (in Chinese)
[36] SETTELE J, SETTLE W H. Conservation biological control: Improving the science base. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(33): 8241-8243.
[37] ALBRECHT M, KLEIJN D, WILLIAMS N M, TSCHUMI M, BLAAUW B R, BOMMARCO R, CAMPBELL A J, DAINESE M, DRUMMOND F A, ENTLING M HThe effectiveness of flower strips and hedgerows on pest control, pollination services and crop yield: a quantitative synthesis. Ecology Letters, 2020, 23(10): 1488-1498.
[38] CHEN X, JAWORSKI C C, DAI H, LIANG Y, GUO X, WANG S, ZANG L S, DESNEUX N. Combining banker plants to achieve long-term pest control in multi-pest and multi-natural enemy cropping systems. Journal of Pest Science, 2022, 95(2): 685-697.
[39] UESUGI R, KONISHI-FURIHATA R, TABUCHI K, YOSHIMURA H, SHIMODA T. Predacious natural enemies associated with suppression of onion thrips,(Thysanoptera: Thripidae), in intercropped onion-barley agroecosystems. Environmental Entomology, 2023, 52(2): 183-196.
Dominated Species and the Predation Assessment of Natural Enemies on Thrips in Cotton Fields in Xinjiang
DANG WenWen1,2, LIU Bing2, CHU Dong1, LU YanHui2,3
1College of Plant Health and Medicine, Qingdao Agricultural University, Qingdao 266109, Shandong;2State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193;3Western Agricultural Research Center, Chinese Academy of Agricultural Sciences, Changji 831100, Xinjiang
【Objective】Thrips is a major pest in cotton fields in Xinjiang. The objective of this study is to select their dominated predatory natural enemies and assess the predation efficiency on thrips, and to provide scientific insights for the development of biocontrol on thrips in cotton fields.【Method】The population dynamics of thrips and their predators in Korla cotton field in southern Xinjiang were systematically investigated by visual inspection within five-point sampling in different blocks from 2021 to 2022. Predators’ samples were collected at different growth stages of cotton fields and detected by molecular detection, DNA of the whole natural enemy was extracted first, and PCR amplification was performed using the specific primers ofandto obtain the species information of thrips in various natural enemies, then the quantitative food web of predator-thrips was constructed, and the differences of related food web structure at different growth stages of cotton were compared. In addition, basing on the predator-prey functional response model, the predation function of different predators (including adults and juveniles) on thrips was evaluated in laboratory by setting the prey density of 50, 100, 200, 300 and 400 individuals per Petri dish.【Result】The population density of thrips was low at the seedling and budding stages but was high at the flowering-bolling stage. Annual population peaks occurred from late-July to early-August. Basing on the population dynamic,is the dominated predator for thrips that occurred across the seeding, budding, flowering and bolling stages, whileis the dominated predator in the late of flowering-bolling stage. The results of molecular detection showed the detection rate of thrips was high in the gut ofin the whole growth period of cotton, the detection rate of thrips in the gut ofat seeding-budding stage was high, and the detection rate in the gut ofwas also high at flowering-bolling stage. The predatory function response in laboratory indicated the predation ability of different predators onincreased with the increase of prey density, the predation function of the larvae of, the nymph ofand the larvae ofwas consisted with the Holling-Ⅱ model, whereas the predation function of the adult, such asandwas suited with the Holling-Ⅲ model; when high prey density of 400 individuals thrips per Petri dish was provided, the consumed number of thrips for the adult ofwas significantly higher than the larvae, the adult and nymph ofand the larvae of【Conclusion】is the dominant predator for thrips across the whole cotton growth period, andis the dominant predator in the late of flowering-bolling stage. This study provides a scientific insight for the utilization of predators to suppress thrips in cotton fields in Xinjiang.
cotton; thrips;;; predation; food web
10.3864/j.issn.0578-1752.2023.17.009
2023-05-23;
2023-06-20
国家重点研发计划(2022YFD1400300)、财政部和农业农村部:国家现代农业产业技术体系(CARS-15-21)
党雯雯,E-mail:13319846572@163.com。通信作者陆宴辉,E-mail:luyanhui@caas.cn
(责任编辑 岳梅)
猜你喜欢
中国棉花(2023年8期)2023-09-07 10:03:05
今日农业(2021年11期)2021-11-27 10:47:17
现代园艺(2017年21期)2018-01-03 06:42:17
江苏农业科学(2016年2期)2016-04-11 23:42:28
现代农业科技(2014年13期)2014-09-18 09:33:38
现代农业科技(2014年14期)2014-09-16 22:44:52
现代农业科技(2014年4期)2014-04-29 21:44:42
新疆农垦科技(2014年9期)2014-02-28 19:20:58
新疆农垦科技(2014年5期)2014-02-28 19:19:56
植物营养与肥料学报(2011年2期)2011-10-26 03:52:30
