
盖塔病毒SC483株cDNA感染性克隆的构建
2023-10-24 08:50:08杜炳辰王铭刘春国王世达魏新宇路雅曼孙振钊刘在斯魏丽丽王靖飞杨德成
中国农业科学 2023年17期
杜炳辰,王铭,刘春国,王世达,魏新宇,路雅曼,孙振钊,刘在斯,魏丽丽,王靖飞,杨德成
盖塔病毒SC483株cDNA感染性克隆的构建
杜炳辰,王铭,刘春国,王世达,魏新宇,路雅曼,孙振钊,刘在斯,魏丽丽,王靖飞,杨德成
中国农业科学院哈尔滨兽医研究所/动物疫病防控全国重点实验室,哈尔滨 150069
【背景】盖塔病毒(Getah virus, GETV)是一种由蚊虫传播的病毒,分类学上属于披膜病毒科甲病毒属成员。该病毒宿主范围广,可感染猪、马、牛、狐狸等多种哺乳动物,人也可以感染,但在人群中造成的危害尚不知晓。动物感染主要临床表现为发热、皮疹、关节炎、繁殖障碍和死胎。GETV在世界范围内流行较为广泛,近年来在我国的流行呈上升趋势,2018年我国南方多个猪场爆发该病,GETV在畜禽养殖及公共卫生方面的危害渐渐受到人们的关注。目前,尚无用于预防和治疗盖塔病毒感染的商业化疫苗和药物。由于对GETV研究较少,其生物学特性、对不同物种的致病性和致病机制以及流行趋势在很大程度上均是未知的。【目的】建立高效的GETV反向遗传操作平台,为深入研究GETV基因组结构与功能、致病机制以及开发新型疫苗奠定基础。【方法】采取化学合成的方式人工合成了两端分别含有锤头状核酶(HamRz)和丁肝病毒核酶(HdvRz)序列的GETV SC483株基因组全长,并克隆至低拷贝pOK12-CMV载体中,从而获得含有GETV SC483株基因组全长cDNA克隆的重组质粒pGETV-SC483。将纯化的重组质粒pGETV-SC483转染BHK-21细胞进行病毒拯救。对拯救病毒进行连续传代、鉴定以及生物学特性分析,并对感染性克隆质粒pGETV-SC483在大肠杆菌中的遗传稳定性进行验证。【结果】重组质粒pGETV-SC483转染BHK-21细胞后,48 h即可观察到GETV感染引起的典型细胞病变,获得的拯救病毒命名为rSC483。分别提取拯救病毒和亲本病毒基因组RNA,对病毒基因组进行RT-PCR扩增、I酶切以及序列测定。结果表明,拯救病毒不同于亲本病毒,其含有人为引入以消去I酶切位点的G4332A突变的分子标记。使用GETV特异性抗体作为检测抗体的间接免疫荧光试验和病毒粒子形态学电镜负染观察结果进一步表明,GETV拯救成功。噬斑形成试验和一步生长曲线试验结果表明,拯救病毒与亲本病毒具有相似的复制能力和增殖特性。另外,感染性克隆质粒pGETV-SC483在大肠杆菌DH5α中连续传代后的测序结果表明,该重组质粒具有良好的遗传稳定性。【结论】成功构建了稳定、高效的GETV SC483株全长cDNA感染性克隆,为GETV生物学特性及致病机理研究以及新型疫苗开发提供了技术平台。
盖塔病毒;cDNA感染性克隆;病毒拯救;反向遗传操作系统
0 引言
【研究意义】盖塔病毒(Getah virus, GETV)为披膜病毒科()甲病毒属()成员,属于虫媒病毒。GETV分布广泛,澳大利亚、菲律宾、印度、日本、韩国、中国以及俄罗斯等地区相继有该病毒分离或动物发病的报道[1-8]。GETV感染谱广,马、猪、牛、蓝狐等均可感染[2, 9-12]。受感染的马表现发热、皮疹、后肢水肿和淋巴结肿大[13-14];而感染猪主要引起新生仔猪发热、关节炎和死亡以及妊娠母猪繁殖障碍[15-17]。猪群在自然界被认为是GETV的主要放大宿主[18]。在流行地区发热病人和健康人血清中均检测到该病毒特异性抗体,表明GETV可以感染人,具有重要的公共卫生学意义[5, 19-22]。GETV病毒粒子呈球形,直径约70 nm,有囊膜和纤突。基因组为单股正链RNA,全长约11 – 12 knt。其基因组5′端具有甲基化帽状结构,3′末端具有Poly(A)尾巴结构。基因组包含两个开放阅读框(open reading frame, ORF),分别编码4种非结构蛋白(nsP1、nsP2、nsP3和nsP4)和5种结构蛋白(CP、E3、E2、6K和E1)[23]。近年来,GETV感染引起的猪群发病屡见报道,2018年湖南省某猪场暴发GETV疫情,导致大约200头新生仔猪死亡以及超过150头妊娠母猪产死胎或木乃伊胎;随后在山东某猪厂也出现GETV疫情[24]。目前,对于GETV的研究相对较少,尤其是其基因组的结构与功能、致病性以及致病机理,严重阻碍了有效疫苗和治疗性药物的研发。因此,借助反向遗传技术对盖塔病毒进行相关研究,可为该病的有效防控提供科学依据。【前人研究进展】近期,WANG等[25]、REN等[26]相继利用真核启动子CMV建立了GETV感染性cDNA克隆,但没有很好地解决病毒基因组的精确转录问题,病毒基因组的5′和3′末端含有冗余序列,这些非病毒序列的携带可能会改变病毒的致病性,从而影响病毒基因组结构和功能研究。【本研究切入点】为了建立具有精确病毒基因组全长转录本的GETV反向遗传操作系统,本研究采取基因合成的方式人工合成了两端分别含有锤头状核酶(hammerhead ribozyme, HamRz)和丁肝病毒核酶(hepatitis delta ribozyme, HdvRz)序列的SC483株基因组全长,并克隆至含有CMV启动子的载体中,成功构建了遗传稳定的GETV cDNA感染性克隆,且拯救病毒具有与亲本病毒相似的生物学特性。【拟解决的关键问题】建立GETV SC483株的反向遗传操作系统,为深入研究GETV基因组结构和功能、致病机制以及新型疫苗的研发奠定基础。
1 材料与方法
试验于2021年2月—9月在中国农业科学院哈尔滨兽医研究所动物疫病防控全国重点实验室完成。
1.1 细胞、病毒、菌株及质粒
BHK-21细胞(ATCC CCL-10);猪源GETV毒株SC483(MN478486)由动物疫病防控全国重点实验室分离鉴定并保存;大肠杆菌DH5α感受态细胞购自TaKaRa公司;重组质粒pOK12-CMV由动物疫病防控全国重点实验室构建并保存。
1.2 抗体及试剂
GETV E2蛋白多抗血清由动物疫病防控全国重点实验室制备并保存;Alexa Fluor 488标记的山羊抗鼠IgG购自Thermo Fisher Scientific公司;DMEM培养基、胎牛血清购自Gibco公司;KOD FX Neo高保真PCR酶购自TOYOBO公司;限制性内切酶I购自NEB公司;Simply P total RNA extraction Kit购自BioFlux公司;PrimeScript™ 1st Strand cDNA Synthesis Kit购自TaKaRa公司;Polyethylenimine Transfection Reagent购自Sigma-Aldrich公司;无内毒素质粒大提试剂盒购自TIANGEN公司;2%磷钨酸(pH 6.5)购自中镜科仪公司。
1.3 全长cDNA克隆质粒的构建
在SC483株病毒基因组5′末端引入锤头状核酶(hammerhead ribozyme, HamRz)序列,在病毒基因组3′末端引入丁肝病毒核酶(hepatitis delta ribozyme, HdvRz)序列。在病毒基因组4332 nt位置将G突变为A,使该位置GCGGCCGC(I)变为GCAGCCGC,从而消除I酶切位点作为拯救病毒的分子标记。将设计好的SC483株全基因组全长cDNA序列分为A、B、C、D 4个片段送至吉林库美生物公司进行人工合成并顺次克隆于pOK12-CMV载体中,获得含有正确GETV基因组全长cDNA克隆的重组质粒命名为pGETV-SC483。构建策略见图1。
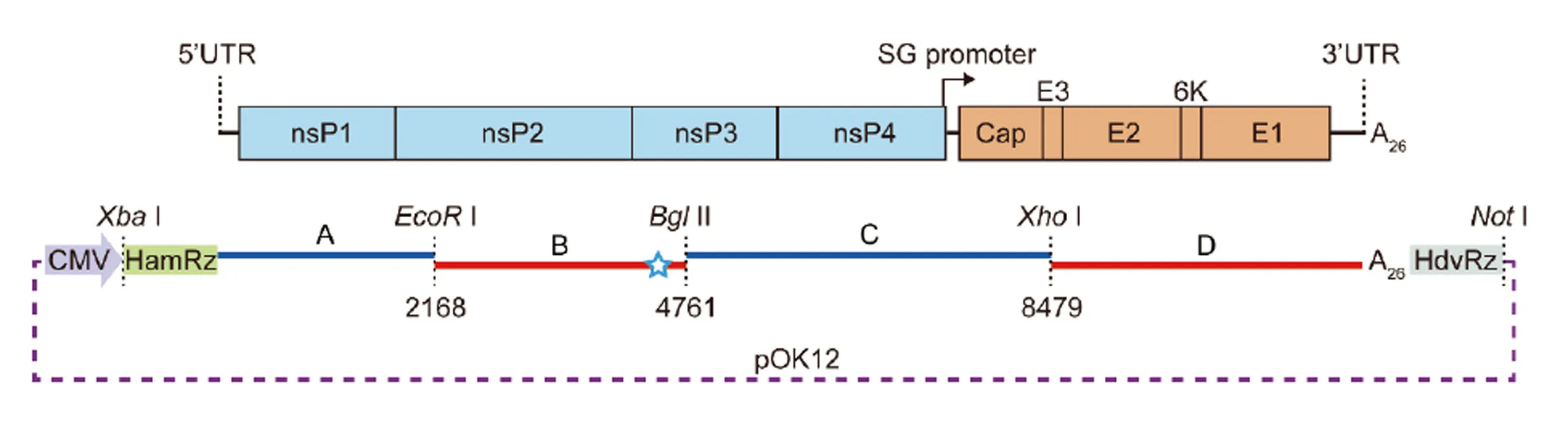
图1 GETV SC483株全长cDNA感染性克隆的构建策略
1.4 病毒的拯救
采用无内毒素质粒大提试剂盒提取重组质粒pGETV-SC483,并按Polyethylenimine Transfection Reagent试剂盒说明书操作步骤将其转染于80%—90%的单层BHK-21细胞。转染后4 h用含2%胎牛血清的DMEM更换培养基继续培养,观察细胞病变(CPE),大约3 d左右收获细胞上清,并在BHK-21细胞上连续传代,拯救病毒命名为rSC483。
1.5 间接免疫荧光(IFA)
将rSC483与SC483-WT接种BHK-21细胞,继续培养16 h;弃去培养基,加入4%多聚甲醛固定10 min,PBS洗3次;用含0.1% TX-100的PBS透膜20 min,PBS洗3次;用含1% BSA的PBS封闭45 min;加入GETV E2多抗血清(1﹕500)室温孵育2 h,PBS洗4次;加入Alexa Fluor 488标记的山羊抗鼠IgG(1﹕2 000)室温孵育1 h,PBS洗4次后在荧光显微镜下观察。同时设不接毒细胞对照。
1.6 拯救病毒粒子的电镜观察
将拯救病毒感染的BHK-21细胞培养物3 500 r/min和8 000 r/min(4℃)各离心30 min,取上清,在含20%蔗糖的NTE缓冲液(pH 7.4)中24 500 r/min(4℃)离心1.5 h后,用100 μL PBS重悬沉淀。取20 μL重悬样品于碳支持膜铜网上吸附2 min,用2%磷钨酸(pH 6.5)染色15 s,干燥后进行透射电子显微镜(日立H-7650)观察。
1.7 拯救病毒的分子标记鉴定
按照Simply P total RNA extraction kit说明书分别提取拯救病毒rSC483和亲本病毒SC483-WT的基因组RNA,然后用PrimeScript™ 1st Strand cDNA Synthesis Kit进行反转录,以此反转录产物为模板,利用引物BF(TGA ATA CCG ACG AGG AAA GCT AC)、BR(CGA CAT AGA CAC GGT ACC GTT TT)进行PCR扩增B片段,将目的片段胶回收纯化后,使用限制性内切酶I进行酶切鉴定,同时进行测序分析。
1.8 噬斑形成试验
将rSC483与SC483-WT稀释至1000 TCID50后分别取100 μL接种于单层BHK-21细胞,37℃吸附1 h,每孔加入3 mL含1%甲基纤维素的DMEM覆盖,37℃继续培养3 d;弃去培养基,加入4%多聚甲醛溶液固定30 min,用0.5%结晶紫染色30 min,自来水洗去染色液,观察蚀斑形成情况。
1.9 一步生长曲线
BHK-21细胞生长至90%单层时,按病毒感染复数(MOI)为5的剂量分别接种rSC483和SC483-WT,37℃吸附1 h后用PBS洗2次,加入含2% 胎牛血清的DMEM继续培养,分别于感染后4 h、8 h、12 h、16 h、20 h、24 h、28 h、32 h和36 h收获细胞上清,每个时间点设置3个重复,测定病毒半数感染量(TCID50)并绘制病毒的一步生长曲线。
2 结果
2.1 病毒拯救
将含有GETV SC483株全长cDNA的重组质粒pGETV-SC483转染BHK-21细胞,转染后48 h可观察到明显的CPE,72 h病变程度可达95%以上,病变细胞变圆、皱缩,呈现与亲本病毒一致的病变形态。将细胞培养物连续传代,CPE出现的时间缩短,病变更加典型。对照细胞形态正常,轮廓清晰(图2)。
2.2 拯救病毒的IFA鉴定
将拯救病毒与亲本病毒分别感染BHK-21细胞,用GETV E2蛋白多抗血清进行IFA检测。结果显示,E2抗体与亲本病毒和拯救病毒感染后的细胞都发生特异性反应,均可观察到特异性荧光,而空白细胞不能与E2抗体发生特异性反应(图3)。
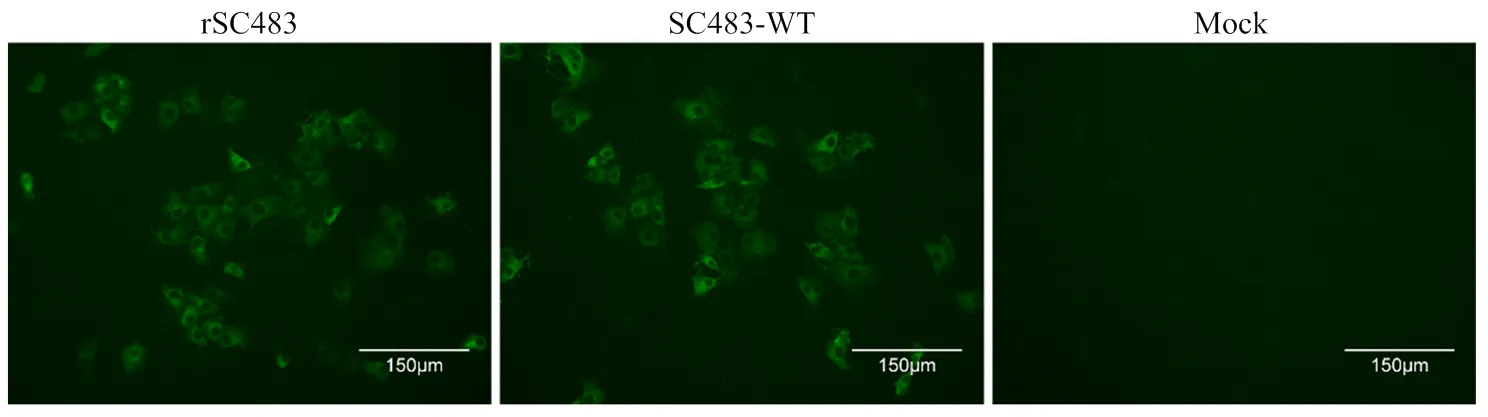
图3 拯救病毒IFA鉴定
2.3 拯救病毒粒子的电镜观察
拯救病毒经浓缩后进行负染色,电镜下可观察到直径约70 nm、有囊膜及明显的纤突的病毒粒子存在,具有典型的GETV的形态学特征(图 4)。
2.4 拯救病毒的分子标记鉴定
提取拯救病毒基因组RNA,用RT-PCR方法扩增基因组B片段,胶回收后用I酶切。结果如图5-A所示,拯救病毒的B片段不能够被I切开,而亲本病毒的B片段可被切成2段。对拯救病毒的B片段进行测序分析,结果显示拯救病毒含有人为引入的G4332A分子标记(GCAGCCGC),消除了I酶切位点(图5-B)。酶切和测序结果表明,拯救的病毒不是来自亲本毒株的污染。
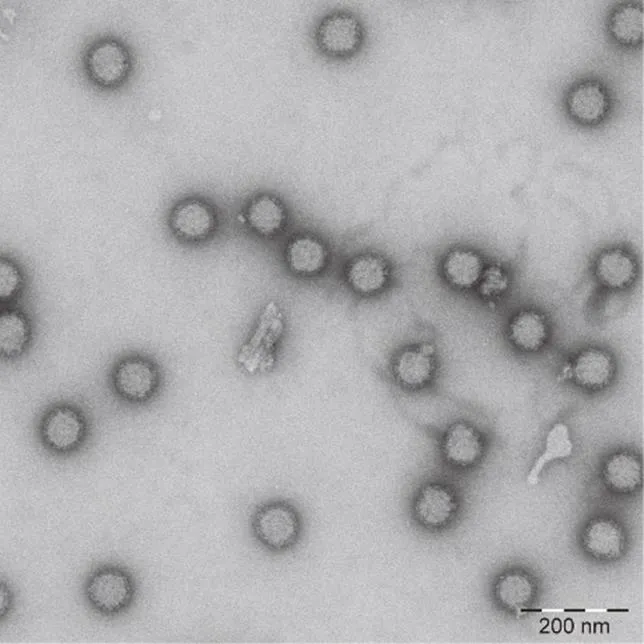
图4 拯救病毒粒子电镜观察
2.5 拯救病毒的蚀斑表型
拯救病毒感染BHK-21细胞后可形成明显的蚀斑,且蚀斑形态大小与亲本病毒相似(图 6)。
2.6 拯救病毒的体外增殖特性
为进一步了解拯救病毒的生长特性,将拯救病毒rSC483与亲本病毒SC483-WT以MOI = 5的剂量感染BHK-21细胞,于不同时间点收集细胞上清,测定其TCID50,绘制拯救病毒和亲本病毒的一步生长曲线。结果显示,拯救病毒与亲本病毒具有相似的复制能力和生长特性(图7)。
2.7 感染性克隆的遗传稳定性
将含有GETV SC483株基因组全长cDNA的重组质粒pGETV-SC483在大肠杆菌DH5α感受态细胞中连续传代10次(P10),对P10代重组质粒进行全序列分析发现,无任何碱基发生突变或缺失,这表明该重组质粒在大肠杆菌中具有良好的稳定性。另外,将P10代重组质粒转染BHK-21细胞,48 h后可观察到明显的CPE,表明病毒拯救成功。这些试验结果表明,GETV SC483株cDNA感染性克隆具有良好的遗传稳定性。
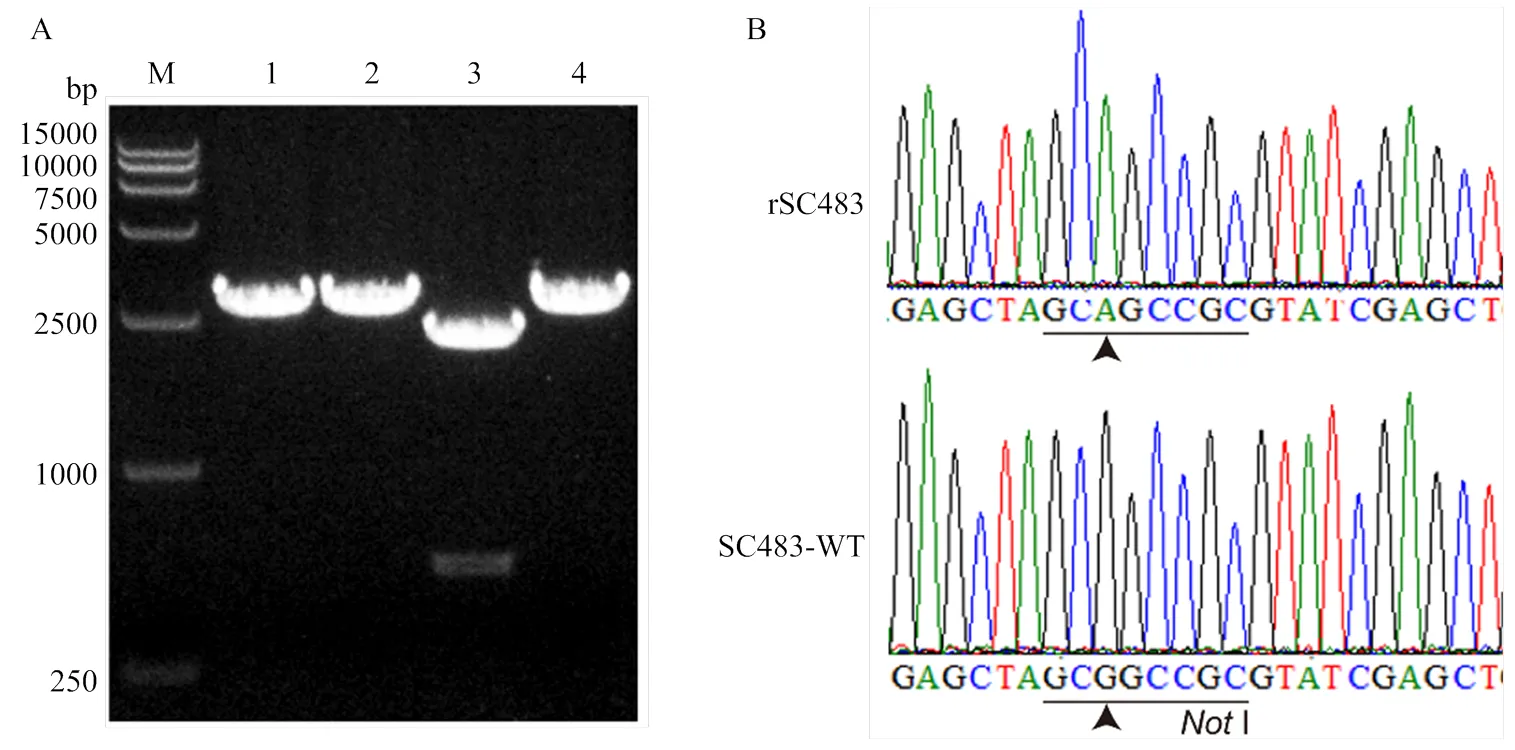
A:Not I酶切鉴定;B:测序鉴定。M:DNA分子量标准;1:亲本病毒RT-PCR产物;2:拯救病毒RT-PCR产物;3:亲本病毒RT-PCR产物Not I酶切;4:拯救病毒RT-PCR产物Not I酶切
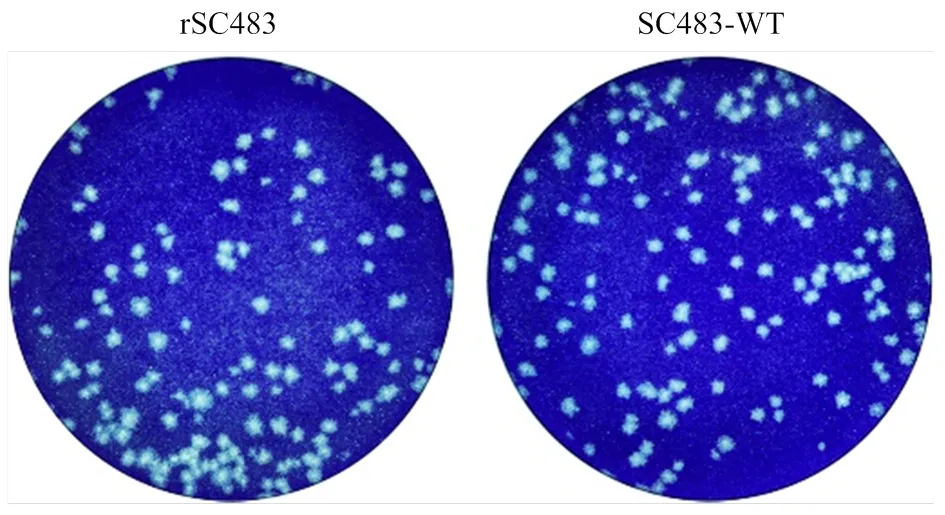
图6 拯救病毒在BHK-21细胞上的蚀斑表型
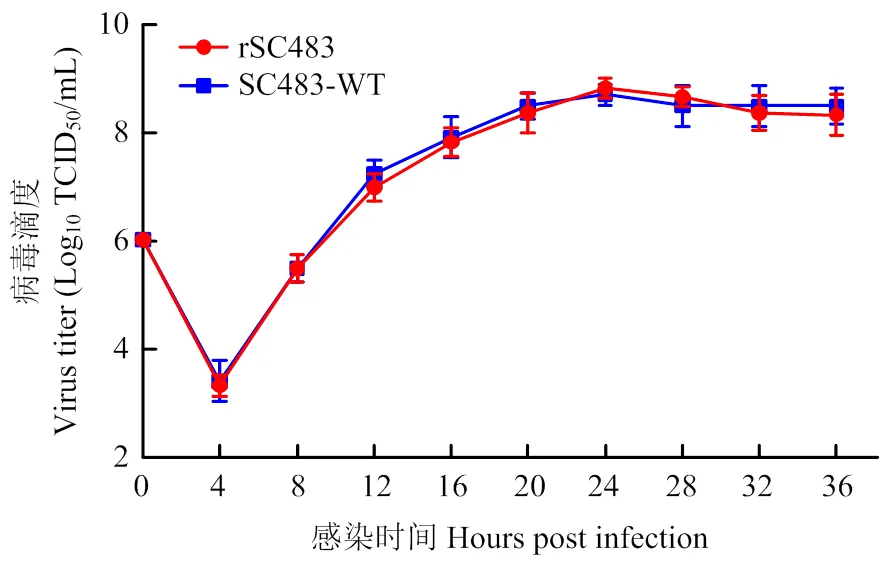
图7 拯救病毒在BHK-21细胞上的一步生长曲线
3 讨论
3.1 GETV反向遗传操作系统建立的意义
GETV是一种人兽共患病病原。近年来,GETV的流行呈上升趋势[9,11,24,27-30]。猪群在自然界中被认为是GETV的主要放大宿主,而我国又是养猪大国,因此GETV对畜牧业的影响以及其潜在的公共卫生威胁应引起足够重视。目前,关于GETV基因的结构与功能以及其致病机理知之甚少。建立GETV的反向遗传操作平台可以在DNA水平上对其基因进行修饰和改造,通过拯救病毒的表型变化来确定相应基因的功能,从而实现对GETV基因组的结构和功能以及致病机制的深入理解,以应对GETV对我国畜禽健康养殖和公共卫生安全日益增长的威胁。
3.2 决定病毒拯救效率的关键因素
RNA病毒反向遗传技术实质上就是将体外或体内获得的病毒基因组RNA导入易感细胞,模拟真实病毒感染,获得完整的具有感染性病毒的过程[31]。转录本的真实性,尤其是其5′末端和3′末端核苷酸准确性对病毒的拯救效率甚至成败起着关键性作用[32]。REN等[26]将GETV基因组直接克隆到真核表达载体pBR322 CMV启动子下游,以BGH Poly(A)信号序列终止。该感染性克隆在靶细胞内无法获得精确的GETV基因组转录本,基因组的两端均含有冗余序列,可能会影响病毒的拯救效率。WANG等[25]通过在GETV感染性cDNA克隆3′末端插入具有自我剪接能力的HdvRz核酶序列以获得准确的病毒3′末端转录本,但基因组5′末端仍含有冗余序列。本研究通过在病毒基因组的5′末端引入具有自我剪切功能的HamRz核酶,在病毒基因组的3′末端引入HdvRz核酶,从而能够在靶细胞中获得精确的GETV基因组全长转录本RNA,进而有利于启动病毒的自我复制实现病毒的高效拯救。此外,感染性cDNA克隆的稳定性也是影响病毒拯救的关键因素。本研究在构建GETV SC483株感染性cDNA克隆的过程中使用低拷贝pOK12载体和慢速生长的受体菌DH5α作为克隆菌株,将GETV SC483株感染性cDNA克隆在DH5α中连续传代10次,没有任何碱基发生突变,表明该感染性cDNA克隆具有良好的遗传稳定性。
4 结论
本研究首次将HamRz和HdvRz核酶序列引入盖塔病毒SC483株全基因组两端,构建了遗传稳定的盖塔病毒感染性cDNA克隆pGETV-SC483,并实现了病毒的高效拯救。盖塔病毒高效反向遗传操作平台的建立为深入研究该病毒基因组结构与功能、致病机制以及开发新型疫苗提供了技术支撑。
[1] KANAMITSU M, TANIGUCHI K, URASAWA S, OGATA T, WADA Y, WADA Y, SAROSO J S. Geographic distribution of arbovirus antibodies in indigenous human populations in the Indo-Australian archipelago. The American Journal of Tropical Medicine and Hygiene, 1979, 28(2): 351-363.
[2] NEMOTO M, BANNAI H, TSUJIMURA K, KOBAYASHI M, KIKUCHI T, YAMANAKA T, KONDO T. Getah virus infection among racehorses, Japan, 2014. Emerging Infectious Diseases, 2015, 21(5): 883-885.
[3] BROWN C M, TIMONEY P J. Getah virus infection of Indian horses. Tropical Animal Health and Production, 1998, 30(4): 241-252.
[4] L'VOV S D, GROMASHEVSKI'I V L, ARISTOVA V A, MOROZOVA T N, SKVORTSOVA T M, GUSHCHINA E A, PETROVA E S, L'VOV D K. Isolation of Getah virus (Togaviridae, Alfavirus) strains in North- Eastern Asia. Voprosy Virusologii, 2000, 45(5): 14-18.
[5] LI X D, QIU F X, YANG H, RAO Y N, CALISHER C H. Isolation of Getah virus from mosquitos collected on Hainan Island, China, and results of a serosurvey. The Southeast Asian Journal of Tropical Medicine and Public Health, 1992, 23(4): 730-734.
[6] KSIAZEK T G, TROSPER J H, CROSS J H, BASACA-SEVILLA V. Isolation of getah virus from nueva ecija Province, republic of the Philippines. Transactions of the Royal Society of Tropical Medicine and Hygiene, 1981, 75(2): 312-313.
[7] TURELL M J, O'GUINN M L, WASIELOSKI L P Jr, DOHM D J, LEE W J, CHO H W, KIM H C, BURKETT D A, MORES C N, COLEMAN R E, KLEIN T A. Isolation of Japanese encephalitis and Getah viruses from mosquitoes (Diptera: Culicidae) collected near Camp Greaves, Gyonggi Province, Republic of Korea, 2000. Journal of Medical Entomology, 2003, 40(4): 580-584.
[8] LI Y Y, LIU H, FU S H, LI X L, GUO X F, LI M H, FENG Y, CHEN W X, WANG L H, LEI W W, GAO X Y, LV Z, HE Y, WANG H Y, ZHOU H N, WANG G Q, LIANG G D. From discovery to spread: The evolution and phylogeny of Getah virus. Infection, Genetics and Evolution, 2017, 55: 48-55.
[9] LIU H, ZHANG X, LI L X, SHI N, SUN X T, LIU Q, JIN N Y, SI X K. First isolation and characterization of Getah virus from cattle in northeastern China. BMC Veterinary Research, 2019, 15(1): 320.
[10] FUKUNAGA Y, KUMANOMIDO T, KAMADA M. Getah virus as an equine pathogen. Veterinary Clinics of North America: Equine Practice, 2000, 16(3): 605-617.
[11] SHI N, LI L X, LU R G, YAN X J, LIU H. Highly pathogenic swine getah virus in blue foxes, Eastern China, 2017. Emerging Infectious Diseases, 2019, 25(6): 1252-1254.
[12] 李楠, 何于雯, 左媛媛, 孟锦昕, 徐天刚, 王静林. 蠓虫源盖塔病毒(SZC30)结构基因分子生物学特征分析. 中国畜牧兽医, 2021, 48(4): 1149-1160.
LI N, HE Y W, ZUO Y Y, MENG J X, XU T G, WANG J L. Analysis of molecular biological characteristics of structural genes of getah virus (SZC30) isolated from midge. China Animal Husbandry & Veterinary Medicine, 2021, 48(4): 1149-1160. (in Chinese)
[13] WADA R, KAMADA M, FUKUNAGA Y, ANDO Y, KUMANOMIDO T, IMAGAWA H, AKIYAMA Y, OIKAWA M. Equine Getah virus infection: Pathological study of horses experimentally infected with the MI-110 strain. Nihon Juigaku Zasshi the Japanese Journal of Veterinary Science, 1982, 44(3): 411-418.
[14] KAMADA M, ANDO Y, FUKUNAGA Y, IMAGAWA H, AKIYAMA Y. Studies on Getah virus-pathogenicity of the virus for horses. Experimental Reports of Equine Health Laboratory, 1981, 18: 84-93.
[15] KUMANOMIDO T, WADA R, KANEMARU T, KAMADA M, HIRASAWA K, AKIYAMA Y. Clinical and virological observations on swine experimentally infected with Getah virus. Veterinary Microbiology, 1988, 16(3): 295-301.
[16] IZUMIDA A, TAKUMA H, INAGAKI S, KUBOTA M, HIRAHARA T, KODAMA K, SASAKI N. Experimental infection ofin swine. The Japanese Journal of Veterinary Science, 1988, 50(3): 679-684.
[17] SHIBATA I, HATANO Y, NISHIMURA M, SUZUKI G, INABA Y. Isolation offrom dead fetuses extracted from a naturally infected sow in Japan. Veterinary Microbiology, 1991, 27(3/4): 385-391.
[18] KUWATA R, SHIMODA H, PHICHITRASLIP T, PRASERTSINCHAROEN N, NOGUCHI K, YONEMITSU K, MINAMI S, SUPRIYONO, TRAN N T B, TAKANO A, SUZUKI K, NEMOTO M, BANNAI H, YOKOYAMA M, TAKEDA T, JITTAPALAPONG S, RERKAMNUAYCHOKE W, MAEDA K.epizootic among wild boars in Japan around 2012. Archives of Virology, 2018, 163(10): 2817-2821.
[19] 白志军, 彭翼飞, 林立辉, 田小东, 陈翠华, 方美玉, 徐春华, 蒋廉华, 陈唯军. 广东、海南两省人血清12种虫媒病毒抗体调查. 中国人兽共患病杂志, 2000, 16(1): 83-84.
BAI Z J, PENG Y F, LIN L H, TIAN X D, CHEN C H, FANG M Y, XU C H, JIANG L H, CHEN W J. Prevalence of human antibody to arbovirus in Guangdong and Hainan Provinces. Chinese Journal of Zoonoses, 2000, 16(1): 83-84. (in Chinese)
[20] 梁勇, 张连山, 刘永为, 董运强, 王志强, 王峻巍, 梁国栋. 河北省涉县发烧样病人盖塔病毒IgM抗体筛查. 中国卫生检验杂志, 2010, 20(10): 2562-2563.
LIANG Y, ZHANG L S, LIU Y W, DONG Y Q, WANG Z Q, WANG J W, LIANG G D. The screening ofIgM antibody to patients with fever from She County in Hebei Province. Chinese Journal of Health Laboratory Technology, 2010, 20(10): 2562-2563. (in Chinese)
[21] 梁勇, 张连山, 刘永为, 董运强, 王志强, 王峻巍, 梁国栋. 河北省涉县正常人群盖塔病毒感染状况筛查分析. 河北医药, 2010, 32(21): 3079-3080.
LIANG Y, ZHANG L S, LIU Y W, DONG Y Q, WANG Z Q, WANG J W, LIANG G D. Screening and analysis ofvirus infection in normal population in Shexian County, Hebei Province. Hebei Medical Journal, 2010, 32(21): 3079-3080. (in Chinese)
[22] 赵春生, 白志军, 彭翼飞, 赵文忠, 陈翠华, 方美玉, 田小东, 蒋廉华, 陈唯军, 刘建伟, 林立辉. 我国南方人鼠虫媒病毒血清流行病学调查. 中国公共卫生, 2001, 17(1): 64-66.
ZHAO C S, BAI Z J, PENG Y F, ZHAO W Z, CHEN C H, FANG M Y, TIAN X D, JIANG L H, CHEN W J, LIU J W, LIN L H. The seroepidemiolgical investigation of arbovirus in humans and rats of the south of China. China Public Health, 2001, 17(1): 64-66. (in Chinese)
[23] ZHAI Y G, WANG H Y, SUN X H, FU S H, WANG H Q, ATTOUI H, TANG Q, LIANG G D. Complete sequence characterization of isolates of(Genus Alphavirus, Family Togaviridae) from China. The Journal of General Virology, 2008, 89(Pt 6): 1446-1456.
[24] YANG T, LI R, HU Y, YANG L, ZHAO D, DU L, LI J, GE M, YU X. An outbreak ofinfection among pigs in China, 2017. Transboundary and Emerging Diseases, 2018, 65(3): 632-637.
[25] WANG N N, ZHAI X F, LI X L, WANG Y, HE W T, JIANG Z W, VEIT M, SU S. Attenuation of getah virus by a single amino acid substitution at residue 253 of the E2 protein that might be part of a new heparan sulfate binding site on alphaviruses. Journal of Virology, 2022, 96(6): e0175121.
[26] REN T W, MIN X L, MO Q R, WANG Y X, WANG H, CHEN Y, OUYANG K, HUANG W J, WEI Z Z. Construction and characterization of a full-length infectious clone of. Virologica Sinica, 2022, 37(3): 348-357.
[27] LU G, OU J J, JI J Z, REN Z X, HU X, WANG C Y, LI S J. Emergence of getah virus infection in horse with fever in China, 2018. Frontiers in Microbiology, 2019, 10: 1416.
[28] REN T W, MO Q R, WANG Y X, WANG H, NONG Z R, WANG J L, NIU C X, LIU C, CHEN Y, OUYANG K, HUANG W J, WEI Z Z. Emergence and phylogenetic analysis of a getah virus isolated in southern China. Frontiers in Veterinary Science, 2020, 7: 552517.
[29] REN X J, FU S H, DAI P F, WANG H Y, LI Y Y, LI X L, LEI W W, GAO X Y, HE Y, LÜ Z, CHENG J X, WANG G Q, LIANG G D. Pigsties near dwellings as a potential risk factor for the prevalence of Japanese encephalitis virus in adult in Shanxi, China. Infectious Diseases of Poverty, 2017, 6(1): 100.
[30] 左媛媛, 元正菊, 何于雯, 任炜杰, 孟锦昕, 王淑娟, 钟江国, 李楠, 王娟, 王静林, 徐天刚. 广州市及其周边地区马群中盖塔病毒血清流行病学调查. 中国动物检疫, 2021, 38(6): 35-39.
ZUO Y Y, YUAN Z J, HE Y W, REN W J, MENG J X, WANG S J, ZHONG J G, LI N, WANG J, WANG J L, XU T G. Sero- epidemiological investigation on the GTV in horses in Guangzhou city and its surrounding areas. China Animal Health Inspection, 2021, 38(6): 35-39. (in Chinese)
[31] 黄耀伟, 李龙, 于涟. 人类及动物RNA病毒的反向遗传系统. 生物工程学报, 2004, 20(3): 311-318.
HUANG Y W, LI L, YU L. The reverse genetics systems for human and animal RNA viruses. Chinese Journal of Biotechnology, 2004, 20(3): 311-318. (in Chinese)
[32] 祁小乐, 高宏雷, 高玉龙, 邓小芸, 步志高, 王晓燕, 王笑梅. 鸡传染性法氏囊病病毒Gt株感染性分子克隆的构建. 中国农业科学, 2007, 40(10): 2343-2349.
QI X L, GAO H L, GAO Y L, DENG X Y, BU Z G, WANG X Y, WANG X M. Development of infectious molecular clones in infectious bursal disease virus Gt strain. Scientia Agricultura Sinica, 2007, 40(10): 2343-2349. (in Chinese)
Construction of Infectious cDNA Clone of GETV SC483 Strain
DU BingChen, WANG Ming, LIU ChunGuo, WANG ShiDa, WEI XinYu, LU YaMan, SUN ZhenZhao, LIU ZaiSi, WEI LiLi, WANG JingFei, YANG DeCheng
Harbin Veterinary Research Institute, Chinese Academy of Agricultural Sciences/ State Key Laboratory of Animal Disease Control and Prevention, Harbin 150069
【Background】Getah virus (GETV) is a mosquito-borne arbovirus, and belongs to thegenus in thefamily. GETV has an extensive host range, which includes multiple mammals, such as pig, horse, cattle, fox and so on, although GETV is not known to cause disease in human. GETV infection often causes clinical symptoms, such as fever, urticarial rash, arthritis, reproductive disorders, and fetal death. GETV is widely prevalent in the world, and the prevalence in China is on the rise in recent years. In 2018, it suddenly broke out in many pig farms in southern China. The harm for livestock and poultry production and public health caused by GETV has got widespread attention. Currently, there is still no specific antiviral drug or vaccine against GETV. Due to the lack of research on GETV, its biological characteristics, pathogenicity, pathogenesis and epidemic properties are largely unknown.【Objective】The aim of this study was to develop a reverse genetics platform of GETV, which could provide a basis for investigating the structure and function of GETV genome, the genetic determinant of pathogenesis, and developing novel vaccine against the virus.【Method】The full-length genome of the GETV SC483 strain flanked by a hammerhead ribozyme (HamRz) and hepatitis delta ribozyme (HdvRz) sequence was chemically synthesized and cloned into a low-copy plasmid pOK12-CMV to generate the recombinant plasmid pGETV-SC483, which was transfected into BHK-21 cells to rescue the virus. Then, compared with the parental virus, the rescued-virus was serially passaged, identified, and analyzed for biological characteristics. The genetic stability of the recombinant plasmid pGETV-SC483 inwas also verified.【Result】The typical cell pathogenic effect (CPE) of GETV was observed in the cells after being transfected with the pGETV-SC483 for 48 h. The rescued GETV, namely rSC483, was successfully obtained, which had a G4332A mutation resulted in the deletion of aI restriction site to differentiate from the parental virus. The rescued virus was further verified by indirect immunofluorescence assay using GETV specific antibody as detection antibody and electron microscopic observation of virus particles. Furthermore, plaque phenotype and growth curve of the rescued virus were similar to that of its parental virus in BHK-21 cells. In addition, the infectious cloning plasmid pGETV-SC483 had good genetic stability inDH5α.【Conclusion】The successfully constructed full-length infectious cDNA clone of GETV SC483 strain provided a technical platform for the study of biological characteristics and pathogenesis of GETV as well as development of novel vaccine against GETV.
Getah virus; cDNA infectious clone; virus rescue; reverse genetics system
10.3864/j.issn.0578-1752.2023.17.019
2022-08-01;
2022-11-16
黑龙江省自然科学基金(YQ2021C038)、动物疫病防控全国重点实验室自主研究课题(SKLVBP202121)
杜炳辰,E-mail:bingchendu@126.com。通信作者王靖飞,E-mail:wangjingfei@caas.cn。通信作者杨德成,E-mail:yangdecheng@caas.cn
(责任编辑 林鉴非)
猜你喜欢
中国糖料(2023年4期)2023-11-01 09:34:46
传染病信息(2022年6期)2023-01-12 08:59:04
环球时报(2022-09-20)2022-09-20 15:18:57
今日农业(2020年24期)2020-12-15 16:16:00
家庭医学(下半月)(2020年2期)2020-05-11 02:07:24
家庭医学(下半月)(2019年10期)2019-11-16 08:59:46
家庭医学(下半月)(2019年9期)2019-10-12 08:04:24
中国果业信息(2019年11期)2019-01-05 20:47:24
西南农业学报(2016年5期)2016-05-17 05:42:23
兽医导刊(2016年12期)2016-05-17 03:51:50
