
慢性牙周炎与慢性阻塞性肺疾病的潜在共同分子机制及其转录因子初探
2023-10-13 04:52:12张晨候珍珍宗颖睿
华西口腔医学杂志 2023年5期
张晨 候珍珍 宗颖睿
郑州大学第一附属医院口腔预防科,郑州 450052
慢性牙周炎(chronic periodontitis,CP)是一种发生在牙周支持组织,由菌斑为始动因子,多种因素共同作用的慢性炎症性疾病[1],CP 与全身系统性疾病,如慢性阻塞性肺疾病(chronic obstructive pulmonary disease,COPD)等密切相关[2]。COPD的特征为持续性呼吸道症状和进行性气流阻塞,是全球第三大死亡原因[3]。由于缺乏有效的生物标记物,较难进行早期诊断。二者都是慢性进行性炎症疾病,同时具有吸烟等共同危险因素,因此有学者[2]认为,二者可能存在一定的因果关系,且具有共同的病理生理过程,但两者共同的基因调控机制尚不清楚。转录因子(transcription factor,TF)作为一种在特定调控序列上结合DNA的蛋白,可通过反转录激活或抑制转录的蛋白质,进而影响细胞完整性或机体稳态[4],具有巨大的治疗潜力,但相关研究较少。
本研究利用CP(GSE10334 和GSE16134)和COPD(GSE76925)的RNA-seq 数据进行生物信息学分析,在转录组水平上揭示二者的cross-talk基因及其相关通路,并寻找了转录因子,为二者的生物学机制提供了新的见解,为开发诊断和治疗靶点的初步研究奠定了基础。
1 材料和方法
1.1 数据收集
从美国国立生物技术信息中心(national cen‐ter for biotechnology information,NCBI)基因表达综合(gene expression omnibus,GEO)数据库下载CP和COPD相关基因表达芯片数据集:CP的数据集为GSE10334 和GSE16134,COPD 的数据集为GSE76925(表1)。下载GPL570 平台和GPL10-558 平台的注释信息,对数据集进行基因定位,并通过R 语言(v4.2.1)GEOquery 包的“exprs”函数获取各数据集的表达矩阵。

表1 CP和COPD的数据集Tab 1 Datasets for CP and COPD
1.2 DEG分析
对病例组和对照组进行DEG 分析。使用R 语言(“limma”包),均设置P<0.05 有统计学意义。GSE10334 和GSE16134 设置|log2FC|>1[5], GSE-76925 数据集中的log2FC 值较低,故设置|log2FC|>0.5[6]。用R 语言中的“VennDiagram”包筛选GSE-10334、GSE16134 与GSE76925 的DEG 交叉基因,即为潜在的cross-talk基因,并绘制韦恩图。
1.3 功能聚类分析
对潜在cross-talk 基因进行基因本体(gene on‐tology,GO)功能富集分析和京都基因与基因组百科全书(Kyoto encyclopedia of genes and genomes,KEGG)通路富集分析。使用R 语言“clus‐terProfiler”包,P值矫正方法采用Benjamini &Hochberg(BH)。在CP 相关数据集GSE10334 和GSE16134 中分别进行|log2FC|相关的功能聚类分析,用R 语言“chordDiagram”包将结果绘制成弦图。
1.4 蛋白质相互作用网络分析
使用STRING 线上数据库(https://cn.string-db.org/),设置置信度得分阈值为0.4,对潜在crosstalk 基因进行差异蛋白质相互作用(protein-protein interaction,PPI)网络分析,并在Cytoscape 软件(版本3.9.0)中可视化。
1.5 核心基因的筛选
使用Cytoscape 的CytoHubba 插件和MCODE插件进一步筛选上述基因。选择4 种拓扑分析算法:最大集团中心性(maximal clique centrality,MCC)、最大邻域分量(maximum neighborhood component,MNC)、度数(Degree)和边缘渗透分量(edge percolated component,EPC),分别选取前10 位基因。通过MCODE 进行模块划分(De‐gree cutoff=2,k-Core=2,node score cutoff=0.2,Max. Depth=100),选取分数最高的2 个模块中的全部基因。将上述结果取交集,得到核心基因,进行功能类聚分析。
1.6 转录因子的预测
利用Cytoscape 中iRegulon 插件预测上述核心cross-talk 基因的重要TF。选择10 k Motifs 数据库(PWM 库),设置富集分数阈值为3.0,AUC 值阈值为0.03,排序阈值为5 000,标准化富集分数(normalized enrichment score,NES)为根据PWM库富集出来计算的得分,选择NES≥7 的转录因子构建调控网络。
2 结果
2.1 DEG的筛选
对3个组织样本数据集进行数据分析,根据设定的阈值进行筛选,GSE10334 包含63 个CP 和27个侵袭性牙周炎患者,共183 个病例样本和64 个对照样本,共筛选出164 个DEG,其中119 个上调,45 个下调;GSE16134 包含65 个CP 和55 个侵袭性牙周炎患者,共241 个病例样本和69 个对照样本,共筛选出208个DEG,其中154个上调,54个下调;GSE76925 有111 个COPD 病例样本,40个对照样本,共筛选出1 408 个DEG,其中557 个上调,851 个下调(表1)。DEG 的火山图如图1A~C 所示。将得到的DEG 取交集,共有21 个潜在cross-talk基因(图1D)。
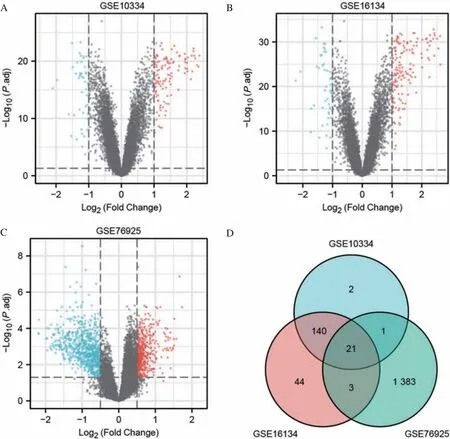
图1 CP和COPD的火山图及其DEG的韦恩图Fig 1 Volcano plots of CP and COPD, Venn diagram of DEGs
2.2 潜在cross-talk 基因的功能聚类分析和其PPI网络
本研究将21 个潜在cross-talk 基因进行GO 分析和KEGG 分析,图2A 显示前3 位的结果:最显著富集的生物学过程为整合素激活(GO:00336-22,校正P=8.71×10-4),细胞定位显著富集在细胞外围(GO:0009897,校正P=0.026),最显著富集的分子功能是CXCR3 趋化因子受体结合(GO:0045236,校正P=0.003)。KEGG 通路最主要富集于细胞因子-细胞因子受体相互作用(hsa04060,校正P=0.007)。图2B、C 分别展示了潜在crosstalk 基因及其在CP 相关数据库GSE10334 和GSE-16134中的log2FC。
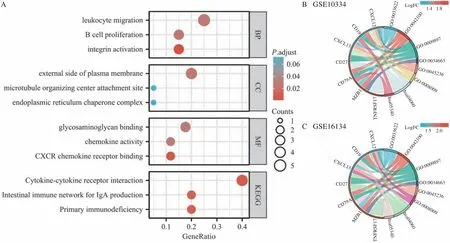
图2 功能富集分析Fig 2 Functional enrichment analysis
STRING 线上数据库构建的PPI网络包括21 个节点和20 条边,结果采用Cytoscape 软件可视化(图3)。筛选出连接最多的前10 个基因为CD19、IRF4、CD79A、CD27、TNFRSF17、SELL、POU-2AF1、CXCL13、FCRLA和CXCL12。表2显示了4 种算法的PPI 网络中前10 个基因的拓扑特征。MCODE 进行模块划分,第一模块包含的基因为CD79A、FCRLA、CD19 和IRF4(得分为3.333),第二模块包含的基因为CD27、SELL 和CXCL13(得分为3.000)。取交集后得到7 个核心基因为CD79A、FCRLA、CD19、IRF4、CD27、SELL 和CXCL13(图4A)。
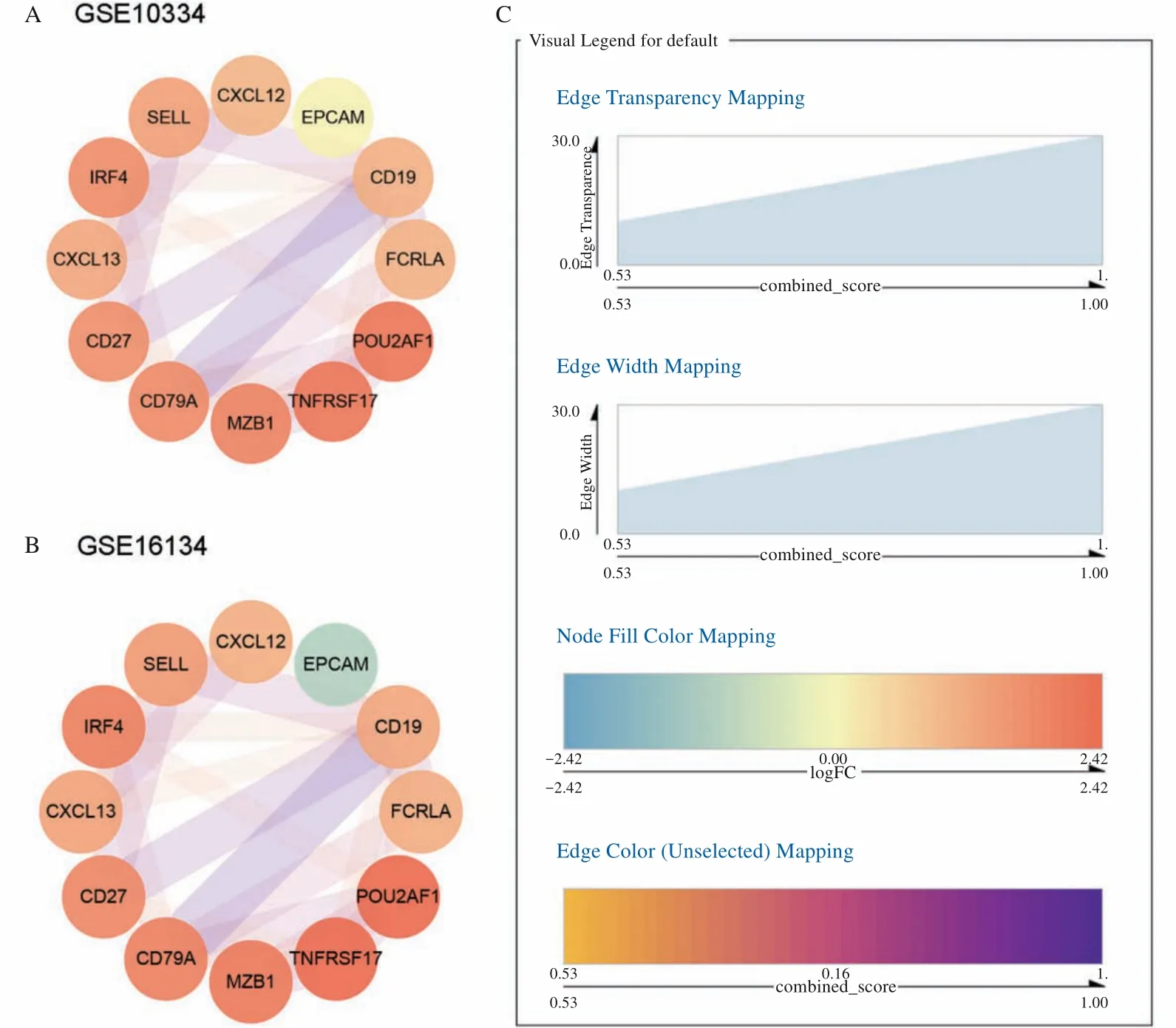
图3 构建的PPI网络Fig 3 Constructed PPI network
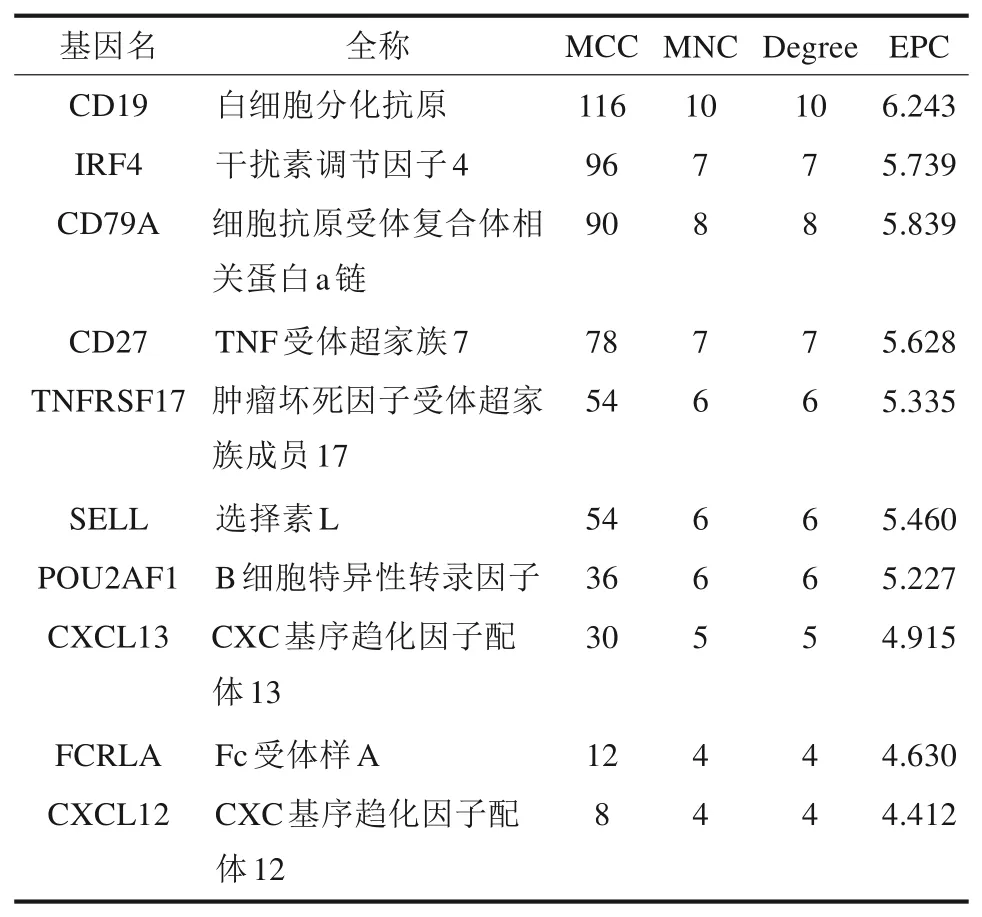
表2 PPI网络中前10个基因的拓扑特征Tab 2 Topological characteristics of top 10 genes in PPI network
2.3 核心基因的功能聚类分析
将得到的7 个核心基因再次进行功能聚类分析,图4 和表3 可见,显著富集的生物学过程包括淋巴细胞分化(GO:0030098,校正P=7.38×10-4)、基于免疫受体体细胞重组的适应性免疫应答(GO:0030098,校正P=7.38×10-4)和B 细胞分化(GO:0030098,校正P=0.001);细胞定位显著富集在细胞外围(GO:0009897,校正P=0.004)、脂筏(GO:0030098,校正P=0.019)和膜微区(GO:0030098,校正P=0.019);最显著富集的分子功能是肝素结合(GO:0008201,校正P=0.025)、糖胺聚糖结合(GO:0005539,校正P=0.025)和硫化合物结合(GO:1901681,校正P=0.025)。KEGG 通路富集分析最主要富集于原发性免疫缺陷(hsa05340,校正P=0.003)、B细胞受体信号通路(hsa04662,校正P=0.007)和细胞因子-细胞因子受体相互作用(hsa04060,校正P=0.060)。
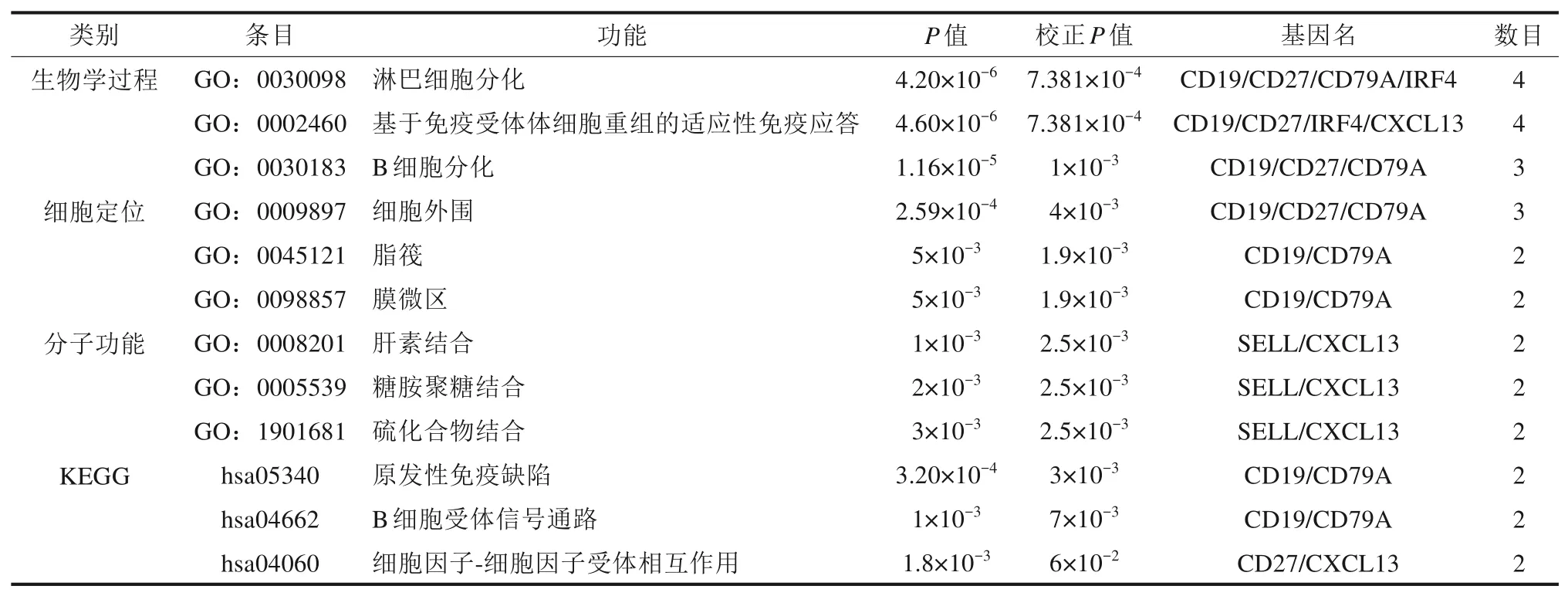
表3 核心基因的GO分析和KEGG分析Tab 3 GO analysis and KEGG analysis of the hub genes
2.4 转录因子的预测结果
共得到7 个符合条件的主调控因子,分别为IRF4、ELF1、GSC、GATA1、PAX3、SRF 和PAX8,其中IRF4 的NES 最高(9.880),调控4 个目标基因(FCRLA、IRF4、CXCL13 和CD19)(表4)。

表4 核心基因的NESTab 4 The NES of hub gene
3 讨论
Takeuchi等[2]通过一项基于900人、5年随访的队列研究表明,CP 为COPD 的独立危险因素,其严重程度与COPD 的风险正相关。本研究整合了CP和COPD的基因表达谱数据,通过分析其crosstalk 基因、富集生物学过程和相关通路,探讨了二者潜在的共同关键分子机制。为了更准确地筛选模块内的核心基因(hub 基因),本研究采用了4种不同的拓扑分析算法,最终结果表明,CD79A、FCRLA、CD19、IRF4、CD27、SELL 和CXCL13可能为连接CP 和COPD 的核心cross-talk 基因。这些基因的潜在相关性基于其在疾病样本中的表达量。其中,CD79A、CD19和CXCL13在CP或COPD中的作用已被报道。同时,参与CP和COPD的通路可能包括原发性免疫缺陷、B细胞受体信号通路和细胞因子-细胞因子受体相互作用。基于结果的多样性及其复杂性,本文将集中讨论这3条信号通路。
原发性免疫缺陷(primary immunodeficiency,PI)可以通过影响免疫系统器官和细胞(T 细胞、B 细胞和自然杀伤细胞等)的成熟、分化和功能,进而破坏细胞和体液免疫或非特异性防御机制,增加患者对自身免疫性疾病、感染和恶性肿瘤的易感性[7]。PI 常伴有组织坏死、溃疡和细菌侵袭,导致早发性、侵袭性牙龈炎和牙周炎[8]。由于缺乏抗体,患者易发生上呼吸道和下呼吸道感染,且反复发作[9]。
虽然CP 的始动因子为牙菌斑,但宿主的免疫反应能影响其进展和严重程度,从而决定机体对CP 的易感性。固有和适应性免疫应答的性质和控制决定了宿主对细菌的应答方式。其中B 细胞是适应性免疫的重要组成部分,可以和活化的CD4+辅助性T 细胞协同产生可溶性抗体,在身体各个部位中和感染因子(体液免疫)[10]。B 细胞受体是由2 种跨膜蛋白(CD79A 和CD79B)结合免疫球蛋白M 组合而成的异源二聚体信号复合物[11],识别抗原后可激活SRC 家族激酶、蛋白酪氨酸激酶和TEC 家族激酶,并招募与CD19 或BCAP 结合的脂质激酶PI3Kδ,进而激活相关基因的表达,这些基因可参与B 细胞增殖、分化、免疫球蛋白产生等[12]。
CD19 与CP 的促炎细胞因子紧密相关[13],CD19+B 细胞是CP 的主要B 细胞成分之一[14]。B 细胞处于免疫系统和骨系统的核心位置,参与CP 的发生发展,其作用机制包括破坏细胞外基质、骨平衡调节[核因子κB 受体活化因子配体(recep‐tor activator of nuclear factor-κB ligand,RANKL)-核因子-κB 受体活化因子(receptor activator of nu‐clear factor-κB,RANK) -骨保护素(osteoprote‐gerin,OPG)],甚至可以作为破骨细胞样细胞发挥吞噬作用[15],RANKL-RANK-OPG 三聚体通过与内皮细胞、上皮细胞和破骨前体细胞相互作用,调节成骨细胞、血管生成和牙周袋形成。B细胞间相互作用释放淋巴毒素(lymphotoxin,LT)和基质细胞,产生稳态趋化因子(CXCL13、CXCL12、CCL21 和CCL19)[16]。研究[17]表明,CXCL13 分泌细胞分布在B细胞区,负责B细胞趋化,与牙龈炎相比,牙周炎患者的牙周袋上皮下结缔组织中含有更多的CXCL13 分泌细胞,其与B 细胞数量相关。Litsiou 等[18]检测了COPD 患者肺滤泡,发现CXCL13 水平升高,并且与淋巴滤泡所占面积相关。B 细胞产生LT 并向CXCL13 趋化,CXCL13又增加B 细胞中膜结合型LT 的表达,进一步刺激CXCL13的产生。因此,本研究结合已被实验验证的结果可以得出,B 细胞受体信号通路可能为CP和COPD的潜在共同分子机制。
细胞因子在CP 和COPD 的进展过程中发挥关键作用,其作为内环境和验证过程的中枢调节因子,可将组织细胞与淋巴细胞和辅助细胞群连接起来,从而影响机体的免疫反应。病变牙周组织中产生的细胞因子等促炎介质可通过血行转移,增加机体的炎症负担,刺激肝脏产生C 反应蛋白、白细胞介素(interleukin,IL)6、转铁蛋白和淀粉样蛋白A等,进而促进肺组织的炎症反应[19]。代表性促炎细胞因子为IL-1 家族和IL-6 家族等,对淋巴细胞的促进和组织破坏具有多效性。其中,牙周炎的严重程度与患者龈沟液中的IL-1β 含量有关[20]。免疫小鼠和大肠杆菌相互诱导产生IL-1β,诱导辅助T 细胞(helper T cell,Th)1 和Th2 的增殖[21],进而造成牙周病的发生。COPD 严重程度也与患者痰中的IL-1β含量有关[22],IL-1β可显著激活巨噬细胞,并分泌趋化因子、炎症因子和基质金属蛋白酶9[23]。另外IL-6 在牙周炎的起始期和急性期起关键作用,促进成骨细胞中受体激活因子的表达,进而促进骨吸收[24]。IL-6可与其他细胞因子协同作用,不仅参与先天免疫和后天免疫,而且参与COPD 的一些全身性特征和共病的恶化,例如胰岛素抵抗、骨质疏松、内皮细胞功能受损和抑郁等[25]。综上所述,CP 和COPD 可能存在IL-1和IL-6等相同的致病细胞因子。
转录因子TRF4 与cross-talk 基因的调控有关,干扰素调节因子可以调节先天和适应性免疫反应,促进炎症反应以及调节免疫细胞分化[26],可能促进了CP 和COPD 的发生发展。但具体作用尚未被证实,应深入研究相关转录因子,为各种疾病治疗的潜在靶点开辟新的可能性。
通过对上述病理生理学中共同致病因子和信号通路的探讨,笔者推测CP 和COPD 可能具有相同的分子机制,在疾病的发生发展中有一定的相关性。但由于样本临床资料的局限性,即未纳入患者的年龄、性别、吸烟习惯和药物治疗等影响因素,并不能充分验证这一推测,因此还需要进一步研究,如充分考虑样本异质性,扩大样本量,对二者人口学指标进行倾向性评分匹配后,进行前瞻性研究;利用单细胞测序技术,在单个细胞的层面上,通过对基因组学、转录组学、微生物组学和空间转录组学等研究明确二者的共同发病机制或潜在的转化机制。无论如何,这提示医师在临床实践中,共同认识、跨学科治疗和预防这两种疾病的必要性。
综上,本研究初步表明核心cross-talk 基因(CD79A、FCRLA、CD19、IRF4、CD27、SELL和CXCL13)和转录因子TRF4 可能参与了CP 和COPD 共同分子机制,并讨论了3 条可能的共同通路:原发性免疫缺陷、B细胞受体信号通路和细胞因子-细胞因子受体相互作用。这将作为相关领域未来研究的基础,为未来药物开发和减轻二者转化风险的研究提供新方向,但仍需进一步地实验验证。
利益冲突声明:作者声明本文无利益冲突。
猜你喜欢
现代临床医学(2021年4期)2021-07-31 07:55:54
昆明医科大学学报(2021年6期)2021-07-31 07:40:38
国学(2020年1期)2020-06-29 15:15:30
数学物理学报(2017年6期)2018-01-22 02:26:53
实用口腔医学杂志(2017年6期)2017-09-19 02:51:32
摄影之友(影像视觉)(2017年1期)2017-07-18 11:12:16
中西医结合心脑血管病杂志(2016年20期)2016-03-01 04:20:37
中国继续医学教育(2015年4期)2016-01-07 07:38:01
中国医疗美容(2015年1期)2015-07-12 10:06:47
医学研究杂志(2015年12期)2015-06-10 06:57:46
