
达乌尔黄鼠(Spermophilus dauricus)褐色脂肪年周期变化
2023-10-13 12:11:00宋士一
沈阳师范大学学报(自然科学版) 2023年3期
宋士一, 刘 语, 任 月, 彭 霞, 杨 明
(1. 沈阳师范大学 生命科学学院, 沈阳 110034;2. 山西农业大学 植物保护学院, 太原 030031; 3. 沈阳师范大学 生物系统进化与农业生态重点实验室, 沈阳 110034)
褐色脂肪组织(brown adipose tissue, BAT)广泛存在于小型哺乳动物中[1],分布于颈部、肩胛骨和主动脉周围,其可减少脂肪堆积,并通过非颤抖性产热快速产生热量,为寒冷环境的动物提供热量,提高生存率[2-3]。副交感神经末梢(parasympathetic nerves)释放去甲肾上腺素,与其β3受体结合后,能够激活解耦联蛋白-1(uncoupling protein 1,UCP 1),促进BAT产热[4]。哺乳动物中,低温是调节BAT活性的重要因素[5],研究发现冬季中缅树鼩(Tupaiabelangerichinensis)BAT重量和UCP 1表达显著高于夏季[6]。BAT最初在冬眠哺乳动物冬眠前准备阶段被发现,称为“冬眠腺”,BAT产热对动物冬眠期间的体温恢复有很重要的作用[1],动物觉醒过程中亦伴随着BAT激活[7]。冬眠动物通过增加体内非颤抖性产热来终止冬眠状态[8]。UCP 1活性在动物冬眠和休眠过程中受到调节。例如,黄鼠在冬眠前BAT质量、线粒体丰度和UCP 1表达增加[9],使BAT的产热能力增强[10]。研究发现,北极黄鼠(Urocitellusparryii)[11]、多纹黄鼠(Ictidomystridecemlineatus)[9]和达乌尔黄鼠(Spermophilusdauricus)[12]冬眠期间BAT中的UCP 1显著增加。
冬眠指活跃状态时体温恒定的一些动物在冬季(有时在晚秋或早春)伴随体温和代谢降低出现的一种昏睡状态。现存哺乳动物很多类群,如针鼹、有袋动物、食虫动物、蝙蝠、啮齿动物和食肉动物中普遍存在冬眠的物种。冬眠是一种生理和行为上的适应,动物通过冬眠前的能量储存与冬眠期间代谢抑制的结合,确保在食物短缺的季节生存下来[13-17]。哺乳动物在冬眠期间表现出体温、代谢率、心率和呼吸频率的显著下降。 冬眠期,动物体温接近环境温度甚至可以降至0 ℃以下,如多纹黄鼠[18]、北极黄鼠[15]和刺猬(Erinaceuseuropaeus)在冬眠期体温下降到环境温度。冬眠期,动物平均代谢率可降低到基础代谢率的5.1%[19],心率和呼吸频率可降至3%[20]。研究表明,在深眠阶段,黄鼠、仓鼠和旱獭的呼吸频率可降到3~10次·min-1,有的动物甚至还会出现呼吸停歇现象。哺乳动物冬眠的最大收益是节约能量。深冬眠期间每日能量消耗可以降低至常温状态的1.2%[21]。研究表明,野外瑞士黄鼠(Spermophilusrichardsonii)在长达8 m的冬眠期会节约88%的能量消耗[22],哺乳动物冬眠期间消耗的能量来源于冬眠前贮存的食物或贮存于身体内的脂肪,这2类动物分别称为贮食类冬眠动物和贮脂类冬眠动物[23]。贮脂类冬眠动物生活史的年周期主要包括繁殖、生长、育肥和冬眠等过程[16],其显著特征是在活跃季节尤其是冬眠前大量摄食并在体内贮存脂肪,这个阶段被称为动物育肥期,冬眠动物在秋季冬眠前进行体脂贮存可使体重增加1倍[24-26], 进入冬眠期会停止摄食[27]。研究表明,育肥期的东北刺猬体重、肥满度、消化器官和生殖器官重量显著增加[28]。冬眠期,动物主要以脂质代谢供能为主[14],动物体重呈下降趋势。
达乌尔黄鼠是一种典型的贮脂类冬眠动物, 大约冬眠5~6 m,由于分布广、数量多、好采集等特点而成为典型的研究冬眠的实验动物[28]。该物种主要分布在中国北方、蒙古及俄罗斯等地。本组已有的研究结果表明,达乌尔黄鼠在冬眠前通过增加摄食量来增加体重,并且在冬眠期间体重缓慢降低[28-29]。其活动行为、代谢率、体温、摄食量、血清中甲状腺激素和BAT中UCP 1表达亦产生年周期性变化[28]。本研究在之前研究的基础上,采集不同时期的达乌尔黄鼠,分别检测其体重、摄食量、肩胛骨褐色脂肪重量和褐色脂肪中UCP 1的表达,探究达乌尔黄鼠年周期褐色脂肪变化,来阐明冬眠动物褐色脂肪年周期的适应机制。
1 材料与方法
1.1 实验动物
本研究使用的达乌尔黄鼠于2015年5月和2016年5月在内蒙古通辽市(122.16°E,43.37°N)捕获。带回至沈阳师范大学动物房,单笼饲养,饲养笼大小为48 cm×35 cm×20 cm,用碎刨花做巢材,喂饲以充足的标准鼠粮(沈阳前民饲料厂)及饮水。育肥期动物房温度为(23±2) ℃,光照∶黑暗=12∶12,冬眠期动物房温度为(5±2) ℃,全黑。每周测量动物体重一次,以确定其增肥状况。
1.2 动物分组(animal grouping)
动物体重的变化被用来估计育肥期间的3个不同阶段。 包括: 1)起始育肥期(early-fattening, EF,n=8), 此时动物的体重开始增加; 2)快速育肥期(mid-fattening, MF,n=8), 动物体重较EF期显著增加; 3)育肥完成期(post-fattening, PF,n=7), 即冬眠前动物体重下降时期。 PF期结束后立即将实验动物转移到全黑暗、环境温度为(5±2) ℃的寒冷房间, 水食自取。 使用冬眠时间的长度来估计冬眠期间的3个不同阶段, 包括: 1)冬眠早期(early-hibernating, EH,n=8), 动物进入冬眠后1 m; 2)冬眠晚期(late-hibernating, LH,n=8), 动物进入冬眠后4 m; 3)出眠期(arousal, AR,n=8), 动物结束冬眠后1 w。
1.3 体重、摄食量、BAT重量测定
采用电子天平(上表电子仪器厂,SB10001)测量达乌尔黄鼠体重,1 w 1次,连续测量,并记录。以第1 d称食物重量为A(g),喂食3 d后,回收食物,称重为B(g)。每天摄食量(g)=(A-B)/3。采用电子天平测量达乌尔黄鼠BAT重量,并记录。
1.4 UCP1表达检测
具体步骤如下:
1) 蛋白样品制备; 2)安装设备,测漏,配制分离胶和浓缩胶; 3)蛋白Marker和待测样品上样; 4)电泳,58 mV下电泳上样样品,3~5 h; 5)转膜,150 mA下转膜,2 h; 6)封闭液孵育4 ℃,3 h; 7)一抗孵育,4 ℃,过夜; 8)回收一抗,1×TBST缓冲液清洗3次,每次5~10 min; 9)二抗孵育,室温,2 h; 10)回收二抗,1×TBST缓冲液清洗,4次,每次5~10 min; 11)曝光,显影; 12)灰度检测分析,用Image-J软件对所得条带灰度测量分析,并记录。
1.5 数据统计分析
使用SPSS 20.0系统进行数据分析,各时期体重、摄食量、BAT重量和解耦联蛋白1(UCP 1)量,均用单因素方差分析(one-way analysis of variance)的方法来分析,用Excel绘制图表,数据均以平均值±标准误(Mean±SE)形式表示。p<0.05表示差异显著,p<0.01表示差异极显著。
2 结 果
2.1 黄鼠的体重变化
对黄鼠体重测量发现,黄鼠体重(图1)呈年周期变化模式,黄鼠在起始育肥期体重开始增加,达到体重最高峰时,约增长了70%的重量,平均每天增长(1.92±0.21) g,入眠前期体重下降,冬眠阶段黄鼠体重缓慢降低,平均每天降低(0.74±0.05) g。
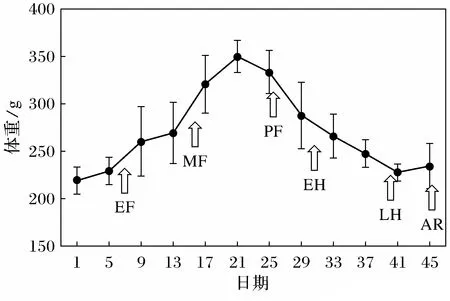
图1 黄鼠年周期体重变化图Fig.1 The annual change of body weight
2.2 不同时期黄鼠BAT重量
黄鼠BAT重量结果如图2所示,EL组黄鼠BAT重量显著高于其他组,为(2.40±0.17) g,AR组黄鼠BAT重量较LH组显著下降。育肥期,黄鼠BAT重量缓慢增加。

图2 不同时期BAT重量变化Fig.2 The weight of BAT at different stages
2.3 不同时期BAT中UCP 1表达
不同时期黄鼠BAT中UCP1表达结果如图3所示,EF组黄鼠BAT中UCP 1表达为(0.844±0.07), MF组BAT中UCP 1表达为(0.960±0.07),PF组BAT中UCP 1表达为(1.169±0.18),EH组BAT中UCP 1表达为(1.151±0.07),LH组BAT中UCP 1表达为(1.18±0.09),AR组黄鼠BAT中UCP 1表达为(0.881±0.08),PF组、EH组、LH组黄鼠UCP 1表达显著高于其他3个组。
3 讨 论
在冬眠前储存大量脂肪、增加体重对动物冬眠有很重要作用[30],本研究表明,达乌尔黄鼠体重呈年周期变化,繁殖期后开始育肥,体重在冬眠前下降,冬眠阶段缓慢下降。Yan等[31]检测金背黄鼠(SpemtophilusLaterali)冬眠阶段体重的变化,在育肥阶段成年雌性金背黄鼠体重平均每天增加(1.22±0.15) g,冬眠阶段金背黄鼠平均每天降低(0.54±0.05) g,与本实验结果一致,提示达乌尔黄鼠作为一种典型的贮脂型冬眠哺乳动物,冬眠前育肥,为冬眠时储存能量,并在冬眠期间消耗脂肪。
BAT产热在动物冬眠和冬眠觉醒中起着重要作用[32-33]。研究发现,金黄黄鼠(Mesocvicetusauratus)在冬眠阶段中消耗约255 mg BAT[34],蝙蝠(Eptesicusvulturnus)出眠后BAT重量显著减少[35]。孙金生和曾缙祥[36]发现东北刺猬入眠前BAT重量显著增加,出眠期BAT重量显著下降。本研究发现达乌尔黄鼠在冬眠阶段BAT重量显著高于育肥阶段,且出眠期黄鼠BAT重量显著降低,结果提示黄鼠在冬眠前储存大量BAT,出眠期消耗BAT,将体温恢复到活动状态水平,进而觉醒。UCP 1是BAT产热蛋白,主要通过消耗质子梯度来产生热量[37]。研究发现,冷驯化下大绒鼠BAT中UCP 1表达显著增加[38],冬眠期多纹黄鼠BAT中UCP 1显著增加。本研究发现黄鼠在完成育肥期UCP 1已显著增加,并在冬眠期保持高水平,出眠期显著下降,推测在完成育肥期黄鼠增加UCP 1蛋白为冬眠做准备,并在出眠期通过UCP 1产热,进而觉醒。本研究组之前发现黄鼠冬眠期UCP 1蛋白的mRNA表达增加[28],与本实验结果相符。
综上所述,达乌尔黄鼠体重呈年周期变化,在冬眠前进行育肥,冬眠阶段体重缓慢下降,冬眠期黄鼠增加BAT重量和UCP 1表达量来维持体温,出眠期通过UCP 1产热,进行觉醒。
猜你喜欢
作文周刊·小学一年级版(2022年24期)2022-06-18 13:11:03
现代畜牧科技(2021年6期)2021-07-16 05:50:12
森林工程(2020年6期)2020-12-14 04:26:52
江苏农业科学(2020年8期)2020-06-01 07:58:51
中外文摘(2019年20期)2019-10-24 08:39:14
云南师范大学学报(自然科学版)(2015年5期)2015-12-26 12:46:16
电源技术(2015年7期)2015-08-22 08:48:24
好孩子画报(2015年6期)2015-05-30 10:48:04
电影(2015年12期)2015-01-05 04:23:58
知识窗(2010年1期)2010-05-14 09:07:46
