
lncRNA MALAT1对哮喘小鼠Th17/Treg细胞平衡的影响
2023-10-11 06:59:46朱海金王诏玉单文琪王影常明汪雪峰
江苏大学学报(医学版) 2023年5期
朱海金, 王诏玉, 单文琪, 王影, 常明, 汪雪峰
(江苏大学附属医院 1. 儿科, 2. 中心实验室, 江苏 镇江 212001)
支气管哮喘是一种异质性疾病,通常以气道慢性炎症为主要特征,多种细胞和细胞因子在其中发挥作用,尤其是肥大细胞、嗜酸性粒细胞、T淋巴细胞、巨噬细胞、中性粒细胞和上皮细胞[1]。哮喘的发病机制复杂,既往认为Th1/Th2细胞失衡是造成哮喘的免疫学发病基础,且哮喘的进展伴随着T淋巴细胞向Th2方向分化,释放Th2型细胞因子[2-3]。但中重度哮喘患者支气管肺泡灌洗液中发现中性粒细胞增多、Th17细胞因子IL-17A、IL-17F和IL-22增多[4]。近年来研究报道,Th17/Treg细胞失衡也参与哮喘的发病[5],但其具体作用机制尚未阐明。
长链非编码RNA(lncRNA)是转录物长度超过200 nt且缺乏蛋白质编码能力的RNA分子,已成为多种生理和病理过程的关键分子,并参与包括哮喘在内的多种疾病的发病机制[6-7]。与lncRNA家族中的大多数成员不同,lncRNA MALAT1在各种哺乳动物中大量表达并在进化上保守,是最早发现的与人类疾病相关的lncRNAs之一,尤其在各种癌症的发展和进展中起关键作用[8]。Feng等[9]研究发现食物过敏小鼠高表达MALAT1,其可能通过促进树突细胞分泌IL-6,并抑制Treg的调节功能,从而诱导食物过敏反应。本研究还发现lncRNA MALAT1在哮喘小鼠脾组织中高表达,高表达的MALAT1与哮喘小鼠Th17/Treg细胞、转录因子Rorc/Foxp3的变化密切相关。由此,MALAT1可能通过诱导Th17/Treg细胞的平衡,参与哮喘小鼠炎症的发生。本研究通过构建哮喘小鼠模型,探讨Th17细胞、Treg细胞、lncRNA MALAT1在哮喘发病中的作用,为哮喘干预提供一个新的方向。
1 材料与方法
1.1 实验动物
SPF级雄性BALB/c小鼠20只,6~8周龄,体重20~25 g,购自江苏大学实验动物中心,许可证号:SCXK(苏)2018-0008,饲养于SPF级环境中。动物实验按照《实验动物护理和使用指南》进行,并经江苏大学动物研究伦理委员会批准(许可证号:UJS-IACUC-2021041201)。
1.2 主要试剂与仪器
卵清蛋白(美国Sigma公司),RPMI 1640完全培养基和胎牛血清(美国Gibco公司);佛波酯/离子霉素混合物和布雷非德菌素混合液(联科生物技术公司);流式染色荧光抗小鼠抗体CD4-FITC、Foxp3-PE、GATA3-PE、IL-17-PE(美国eBioscience公司),流式染色荧光抗小鼠抗体CD45-BV421(美国Biolegend公司),Trizol(诺唯赞生物科技公司),引物MALAT1、GAPDH(擎科生物科技公司),逆转录试剂盒和荧光定量PCR试剂盒(诺唯赞生物科技公司)。定量PCR仪和离心机(美国Thermo Fisher公司),正置荧光显微镜(日本Olympus公司),流式分选仪(美国BD公司)。
1.3 哮喘小鼠模型建立
将小鼠随机分为对照组和哮喘组,每组10只。参照文献[10]方法,分别在第0、7、14天,对照组小鼠给予腹腔注射0.2 mL PBS,哮喘组小鼠给予腹腔注射0.2 mL卵清蛋白混合液(50 μg V级卵清蛋白溶解于0.2 mL PBS中,并加入10%氢氧化铝凝胶配制成混悬液)。从第21天开始,哮喘组给予2.5%卵清蛋白溶液(溶于PBS)雾化进行哮喘激发,1次/d,每次雾化40 min,持续至第27天,共计7 d。最后一次雾化后次日处死所有小鼠。
1.4 HE染色与PAS染色观察肺组织病理变化
取出肺组织固定于4%甲醛溶液中,常规石蜡包埋。石蜡切片,常规HE染色后置于荧光显微镜下观察小鼠肺组织炎症变化。PAS染色步骤:将染色的肺组织浸泡在二甲苯中10 min后取出,用无水乙醇浸泡洗脱二甲苯后分别放入95%、85%、70%乙醇溶液中进行水化。将高碘酸溶液滴加在水化后的肺组织切片,进行染色15 min,用自来水冲洗5 min,然后浸入蒸馏水洗2次,最后擦去切片表面的残留水分。滴加Schiff氏液后静置10 min,用流水冲洗5 min,冲洗晾干后滴加苏木素液染色3 min,用流水冲洗5 min。染色完毕后,需对肺组织切片采取梯度脱水处理,分别在80%、95%以及无水乙醇中浸泡5 s、2 min和2 min。用二甲苯浸泡脱水后的肺组织,浸泡2次,每次浸泡时间为4 min,然后将肺组织切片进行风干,并使用中性树胶进行封片。封片后在荧光显微镜下进行观察。
1.5 制备小鼠脾脏单细胞悬液
小鼠处死后浸泡于75%乙醇消毒,在无菌环境下取出小鼠脾脏并置于筛网中进行研磨。研磨后用3 mL PBS冲洗筛网,收集至15 mL离心管中,以1 500 r/min、4 ℃离心5 min,去除上清液。用1 mL PBS重悬细胞,加入红细胞裂解液,静置10 min,充分裂解红细胞后以1 500 r/min、4 ℃离心5 min,去除上清液。用1 mL PBS重悬细胞得到小鼠脾脏单细胞悬液。
1.6 ELISA法检测血清中卵清蛋白特异性IgE抗体
小鼠采血后,静置至凝固,4 000 r/min离心25 min收集上清液,96孔板包被4 ℃过夜,PBST洗板,用5%牛奶封闭,加入血清,加入山羊抗小鼠IgE抗体,37 ℃温育2 h,PBST洗板,再加入兔抗山羊抗体,37 ℃温育1 h,PBST洗板,加入TMB显色液,37 ℃孵育15 min,加入终止液终止反应,在450 nm波长检测光密度值。
1.7 流式细胞术检测脾细胞中ILC-2、Th17和Treg细胞
参照文献[11]方法,对2型天然淋巴细胞(ILC-2)、Th17和Treg细胞经CD4-FITC、CD45-BV421、GATA3-PE、IL-17-PE、Foxp3-PE染色后,用流式细胞分析仪进行细胞检测。ILC-2表型为CD4-CD45+GATA3+,Th17细胞表型为CD4+IL17+,Treg细胞表型为CD4+CD25+Foxp3+。
1.8 qRT-PCR检测脾组织中MALAT1及Rorc/Foxp3相对表达量
提取小鼠脾组织的总RNA,逆转录成cDNA,按照试剂盒说明检测lncRNA MALAT1及Rorc/Foxp3的表达,用GAPDH对MALAT1、Rorc/Foxp3的表达进行标准化。Rorc和Foxp3购自美国Genecopoeia公司。MALAT1及GAPDH引物由中国擎科生物合成,引物序列如下:MALAT1上游5′-TCCAATCTGCTGCTATTAG-3′,下游5′-CAACAACCACTACTCCAA-3′;GAPDH上游5′-AGCTTGTCATCAACGGGAAG-3′,下游5′-TTTGATGTTAGTGGGGTCTCG-3′。PCR反应体系:2×ChamQ Universal SYBR qPCR Master Mix 10 μL,上、下游引物各0.4 μL,模板2 μL,加水至20 μL。PCR扩增程序:95 ℃预变性30 s;95 ℃变性30 s,60 ℃退火和延伸10 s,40个循环。用2-ΔΔCt半定量的方法确定各基因的表达水平。
1.9 统计学分析

2 结果
2.1 卵清蛋白成功诱导小鼠气道炎症变化
HE染色结果显示,与对照组相比,卵清蛋白诱导的哮喘小鼠气道炎症变化明显,支气管周围存在明显的炎性细胞浸润,管壁增厚,气道上皮细胞破坏或不规则。PAS染色结果显示,与对照组相比,哮喘组的支气管杯状细胞及黏液分泌增多(图1A)。同时,小鼠脾脏细胞中ILC-2比例增多(t=8.35,P<0.01),见图1B。另外,与对照组比较,哮喘组卵清蛋白特异性IgE增高(t=4.17,P<0.05),见图1C。结果表明成功建立哮喘小鼠模型。

A:各组小鼠肺组织病理变化(HE染色和PAS染色,×400);B:流式细胞术检测小鼠脾细胞中ILC-2比例;C:ELISA法检测小鼠血清卵清蛋白特异性IgE图1 小鼠肺组织HE、PAS染色和脾细胞中ILC-2比例
2.2 哮喘小鼠Th17/Treg细胞比例失调
与对照组相比,哮喘组小鼠脾脏中Th17细胞比例显著升高(t=8.08,P<0.01),而Treg细胞比例显著下降(t=16.76,P<0.01)。与之相对应的是Th17细胞的关键转录因子Rorc表达上调(t=3.44,P<0.05),Treg细胞的关键转录因子Foxp3表达下调(t=2.32,P<0.05)。见图2。
2.3 哮喘小鼠脾组织lncRNA MALAT1表达情况
与对照组相比,哮喘组小鼠脾组织lncRNA MALAT1相对表达量显著增加,差异有统计学意义(t=6.10,P<0.01)。见图3。
2.4 哮喘小鼠脾组织lncRNA MALAT1与Th17/Treg细胞的相关性分析
随着小鼠脾组织lncRNA MALAT1表达增加,Th17细胞比例上升,呈正相关(r=0.64,P<0.05),Treg细胞比例下降,呈负相关(r=-0.73,P<0.05)。见图4。
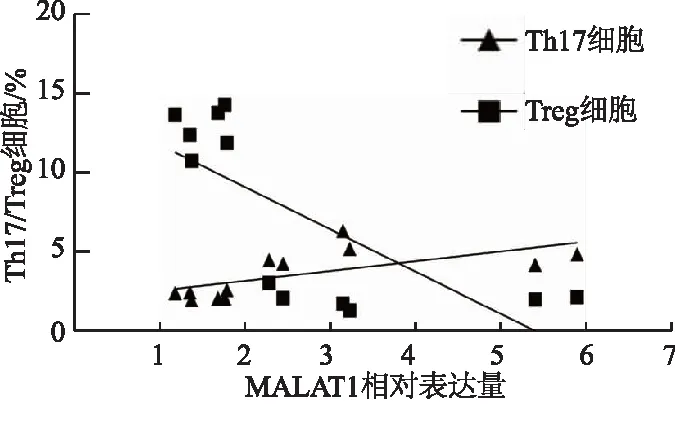
图4 lncRNA MALAT1与Th17/Treg细胞的相关性
2.5 哮喘小鼠脾组织lncRNA MALAT1与Rorc、Foxp3的相关性分析
研究结果表明,随着小鼠脾组织lncRNA MALAT1表达增加,Th17细胞的关键转录因子Rorc表达量也增加,二者呈正相关(r=0.65,P<0.05);Treg细胞的关键转录因子Foxp3表达量减少,二者呈负相关(r=-0.60,P<0.05),见图5。
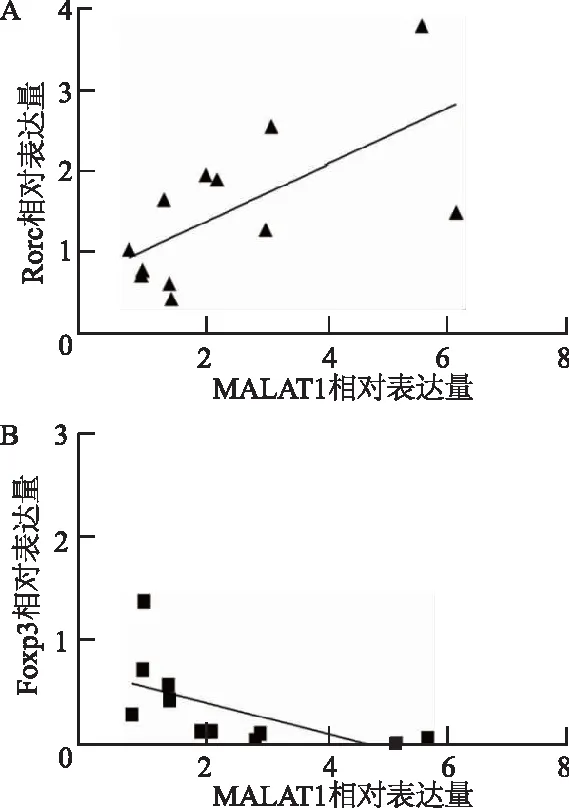
图5 lncRNA MALAT1与Rorc/Foxp3的相关性
3 讨论
哮喘是儿童和成人常见的疾病,在全球范围内呈现出高发病率和高死亡率[12]。目前哮喘的发病机制尚不完全明确,以往普遍认为哮喘的慢性炎症是由嗜酸性粒细胞、ILC-2细胞、Th2细胞以及B细胞共同参与产生[13-14]。ILC-2被认为是Th2细胞对应的先天性免疫细胞,并与Th2细胞一起在过敏性疾病的发病机制中起作用,激活的ILC-2s通过快速产生效应细胞因子,如IL-5在黏膜表面引发过敏性组织炎症[15-16]。
进一步研究发现,不仅Th1和Th2细胞参与哮喘的发病,Th17细胞和Treg细胞在哮喘的发病和进展过程中也发挥了重要的作用。Th17/Treg细胞稳态与哮喘恶化有关,相对于普通哮喘,严重哮喘的IL-17/IL-10和Rorc/Foxp3的比值显著增加[17]。Th17细胞主要分泌细胞因子IL-17,参与哮喘的病理生理学过程[18]。Treg细胞是已知的一类具有免疫抑制作用的T细胞亚群,能够抑制其他细胞的免疫应答,诱导免疫耐受,Treg细胞的缺失或功能异常会导致自身免疫性疾病的发生。本研究通过构建哮喘小鼠模型,探讨Th17细胞、Treg细胞、lncRNA MALAT1在哮喘发病中作用。实验发现,哮喘小鼠不仅在病理上表现为支气管周围出现明显的炎性细胞浸润、管壁增厚、气道上皮细胞破坏或不规则,而且在免疫上表现为ILC-2/Th17细胞的比值增加,Treg细胞比值下降。
lncRNA是炎症反应过程中基因转录的关键调控因子,是一种高度保守的核非编码RNA,是肺癌转移发展的预测标志物,通常以非蛋白转录物为特征,调节多种疾病的病理生理过程[19-20]。Guo等[21]发现lncRNA MALAT的下调可促进NK细胞的免疫恢复,促进NK-92细胞中IFN-γ的分泌。近年来已证实lncRNA MALAT1在气道重塑和气道炎症中发挥重要作用,上调的lncRNA MALAT1通过下调microRNA-216a引起平滑肌细胞的过度增殖、迁移和侵袭,减少凋亡,提高气道平滑肌细胞的活力,而敲除lncRNA MALAT1基因可以抑制气管平滑肌的增殖和迁移[22-23]。Liang等[24]的研究发现哮喘患者血清中lncRNA MALAT1上调可以抑制miR-155在CD4+T细胞内表达,减少Th1型细胞因子(IL-2、IFN-γ)表达水平,增加Th2型细胞因子的分泌,参与调节哮喘患者Th1/Th2细胞平衡。lncRNA MALAT1作为一种新的上皮细胞来源的免疫调节因子,参与特异性的炎症免疫气道微环境,调节树突细胞的成熟过程及其促炎因子的分泌[25]。但是lncRNA MALAT1是否参与调节哮喘Th17/Treg细胞的平衡尚未见报道。本研究证实哮喘小鼠脾组织lncRNA MALAT1相对表达量较对照组增加,与Liang等[24]发现哮喘患者血清lncRNA MALAT1上调一致;哮喘小鼠脾细胞中lncRNA MALAT1表达与Th17细胞比例和Rorc呈正相关,与Treg细胞比例呈负相关。另外,本研究发现lncRNA MALAT1表达与Th17细胞的关键转录因子Rorc和Treg细胞的关键转录因子Foxp3表达关系密切,lncRNA MALAT1与Rorc呈正相关,与Foxp3呈负相关。表明lncRNA MALAT1与哮喘小鼠Th17/Treg细胞失衡密切相关,靶向于lncRNA MALAT1可能有助于改善哮喘小鼠Th17/Treg细胞失衡,但MALAT1调节Th17/Treg细胞平衡的机制尚需进一步研究。
综上所述,哮喘小鼠脾细胞中ILC-2细胞、Th17细胞比例上升,Treg细胞比例下降;哮喘小鼠脾细胞中lncRNA MALAT1的表达增加,并且与Th17细胞比例呈正相关,与Treg细胞比例呈负相关,而且lnc MALAT1的表达与Rorc的表达呈正相关,与Foxp3的表达呈负相关。因此,lncRNA MALAT1可能通过上调Rorc的表达或下调Foxp3的表达参与调节哮喘小鼠Th17/Treg细胞平衡,且在哮喘的发生和进展中发挥重要作用。
猜你喜欢
现代临床医学(2021年4期)2021-07-31 07:55:54
国际呼吸杂志(2019年22期)2019-12-09 09:20:26
国际呼吸杂志(2019年5期)2019-03-30 01:38:20
国际呼吸杂志(2019年3期)2019-03-01 05:39:06
国际呼吸杂志(2019年2期)2019-02-14 06:11:26
华东师范大学学报(自然科学版)(2017年1期)2017-02-27 13:41:06
高师理科学刊(2016年8期)2016-06-15 20:27:48
中西医结合心脑血管病杂志(2016年20期)2016-03-01 04:20:37
中国组织化学与细胞化学杂志(2016年4期)2016-02-27 11:16:04
医学研究杂志(2015年12期)2015-06-10 06:57:46
