
川西北高原短芒型老芒麦种质形态变异及遗传亲缘关系分析
2023-10-08 09:24:44余静菠陈仕勇桑杰多吉张昌兵姬文琴
草地学报 2023年9期
余静菠, 陈仕勇, 桑杰多吉, 张昌兵, 叶 莉, 姬文琴, 李 进
(1.西南民族大学青藏高原研究院, 四川 成都 610041; 2.西南民族大学畜牧兽医学院, 四川 成都 610041;3.四川省阿坝州林业和草原局, 四川 马尔康 62400; 4.四川省草原科学研究院, 四川 成都 611731)
老芒麦(ElymussibiricusL.)又称西伯利亚披碱草,是禾本科披碱草属中一种多年生异源四倍体牧草(2n=4x=28),含有St和H基因组[1]。它主要分布在我国的北方和青藏高原地区,是我国高寒牧草的代表种之一;具有产量高、营养价值高、抗逆性强等特点,被广泛应用于天然草场改良和人工草场建设,在青藏高原畜牧业和生态工程中发挥着重要作用[2]。
在自然进化的过程中植物身处于不断改变的环境,植物为了提高自身的生存能力,会进化出特定的器官来应对这些变化,例如在外稃顶端形成的芒[3]。芒是幼穗外稃延伸出的坚硬、长针状结构。在自然条件下,芒作为谷物上坚硬的部分,能挂在野生动物的皮毛上,有助于野生种质的种子散布和掩埋,同时能防止虫害以及鸟害[4-5]。但在畜牧业生产中长芒会造成不好的影响,有研究表明长芒对牧草的适口性和动物的营养需求有负面影响[6]。长芒降低了植物中的可消化营养物质,并且家畜在食用过程中可能会刺激胃并产生疼痛[7]。此外,长芒会妨碍收获后的处理、储存和加工活动,在生产应用中往往需要进行断芒或去芒处理,为生产者带来诸多不便[8],所以开展老芒麦短芒种质的筛选和利用具有重要的意义。因此我们需要合理利用野生老芒麦进行短芒型品种的培育,而国内丰富的野生老芒麦资源,也为短芒型种质的筛选和评价提供了便利的条件。
目前,前人对于禾本科植物中芒的调控机理的研究已有一定进展。水稻中通过遗传分析确定了几个与芒的形成和伸长有关的基因,如调控芒形成的An-1和LABA1(An-2)、调控芒伸长的SLP1,DL(OsYABBY),OsETTIN2(OsARF2)RAE2和YABBY基因家族的Tob1(TONGARIBOUSHI1)[9-11]。大麦中,在早期Muller等[12]发现同源异型盒基因Knox3产生的带帽结构,会致使异位分生组织在外稃上发育,从而不能形成正常的芒。大麦中抑制芒伸长的基因Lks2位于7H染色体上,编码SHORTINTERNODES(SHI)家族转录因子[13]。本研究以在川西北高原收集的24份短芒型老芒麦种质为研究材料,以老芒麦品种‘川草2号’和野生老芒麦‘Esxz’为对照,通过对短芒型老芒麦农艺性状和基于目标起始密码子多态性(Start codon targeted polymorphism,SCoT)分子标记的遗传多样性评价,以期为短芒型老芒麦的育种等资源创制工作提供重要的参考。
1 材料与方法
1.1 试验材料
本研究共选用了26份老芒麦种质资源,其中24份为联合攻关课题组前期在四川红原地区经过多年评价筛选出的短芒型老芒麦种质资源,同时选用1份国审品种‘川草2号’及1份采集于西藏的野生老芒材料麦(‘Esxz’)为对照(表1)。

表1 材料编号及名称
1.2 试验地概况
试验地设置在四川省草原科学研究院牧草育种基地,基地位于阿坝藏羌自治州红原县境内,红原县海拔3 504 m,年均温为1.1℃,年降水量为744 mm,气候为大陆性高原寒温带季风气候,无绝对无霜期。冬季严寒,夏季凉爽春秋短,日照充足,昼夜温差大。试验地土壤为高山甸草土,pH值约为6.6。
1.3 研究方法
1.3.1形态指标测定 每份材料随机采集10个长势一致的单株进行测定,测定指标分为数量性状和质量性状,其中质量性状通过观察评分。本研究选取了5个质量性状,评分等级如下。
花序颜色:0——青色,1——紫红色,2——紫绿色,3——浅红色。叶片植株被粉:0——无粉,1——有粉。基生叶鞘被毛:0——无毛,1——少毛,2——多毛。杆生叶片被毛:0——无毛,1——少毛,2——多毛。叶片颜色:0——绿,1——蓝绿,2——黄绿。
50个数量性状于乳熟期(Milk stage,MS)测定,花序、小穗、稃片和颖片分别在上、中、底部测量(上部为距离顶端1/3处,中部为距离顶端1/3~2/3处,底端为距离2/3至最远处)。
茎秆性状有:株高(Plant height)、茎长(Stem length)、茎节数(Number of stem nodes)、茎粗(Stem diameter)。
叶片性状有:旗叶叶长(Flag leaf length)、旗叶叶宽(Flag leaf width)、倒二叶叶长(Penultimate leaf length)、倒二叶叶宽(Penultimate leaf width)。
花序性状有:花序小穗数(Panicle number of inflorescence),上、中、底部的花序小穗数(Panicle number of inflorescence),花序节数(Number of inflorescence nodes)。
穗部性状有:上、中、底部的穗节小穗长(Panicle length of inflorescence)、节间长(Internode Length of inflorescence)及小穗小花数(Floret number of inflorescence)。
颖片性状有:上、中、底部的第一颖长(First glume length)、第一颖宽(First glume width)、第一颖芒长(First awn length of glume)、第二颖长(Second glume length)、第二颖宽(Second glume width)、第二颖芒长(Second awn length of glume)。
稃片性状:上、中、底部的外稃长(Lemma length)、外稃宽(Lemma width)、内稃长(Palea length)、内稃宽(Palea width)。每份材料随机采集10个长势一致的单株,用卷尺、直尺和游标卡尺进行测定。
1.3.2DNA提取 每份材料随机选取10株单株叶片进行混合。采用植物DNA提取试剂盒DP305(北京天根生化科技有限公司)提取材料DNA,通过nanodrop测定其浓度与纯度,并用1.2%琼脂糖凝胶电泳检测完整性,根据检测结果将DNA稀释至10 ng·μL-1后,于-20℃冰箱待用。
1.3.3引物筛选及SCoT-PCR扩增 本研究选用Collard等[14](SCoT1~SCoT36)及Luo等[15](SCoT37~SCoT80)报道的SCoT引物,引物均由南京金斯瑞生物科技有限公司合成。选取8份材料对总共80个SCoT引物进行引物的多态性筛选,共筛选出14对引物。根据扩增条带多态性,条带清晰度来确定对应退火温度。
PCR扩增总体积为15 μL,其中包括:6 μL模板DNA(10 ng·μL-1)、1.5 μL引物(10 μmol·L-1擎科生物技术,北京,中国)、7.5 μLMix预混液(Es Taq DNA Polymerase,Mg2+,dNTPs以及PCR稳定剂和增强剂组成的预混体系)(天根生化科技,北京,中国)。PCR扩增反应在BIO-RAD T100 Termal Cycler PCR仪上进行,扩增程序为:94℃变性3 min;然后进行36个循环的94℃变性1 min,48℃~55℃梯度退火1分钟,72℃延伸2 min;最后在72℃下延伸5 min。待PCR程序结束后,在1倍的TAE缓冲溶液中用1.2%琼脂糖凝胶电泳,通过凝胶成像系统来观察扩增的结果并保存。
1.4 数据处理
形态观察的数据处理与分析在SPSS 26.0进行,对形态指标的遗传多样性评价用变异系数、Shannon-Wiener’s多样性指数,同时进行单因素方差分析。分别对50个数量性状进行等级划分,根据性状平均数(x)和标准差(s)将其划分为10个等级,1级≤x-2 s,10 级>x+2 s,中间每级间差0.5 s,利用SPSS软件基于各形态指标的平均数计算供试材料间的欧氏遗传距离并进行UPGMA聚类分析了解各种质之间的亲疏关系;对质量性状进行赋值,按照公式H=-∑PilnPi来计算Shannon-Weaver’s遗传多样性。
对SCoT标记结果,选取条带清晰且多态性好的引物进行统计,在相同迁移位置上,有带记为1,无带记为0,建立1,0矩阵。通过软件POPGENE 1.32中计算多态性条带百分比(Percentage of polymorphic bands,PPB)、有效等位基因数(Ne)、Nei’ s基因多样性指数(H)和Shannon’s多样性指数(I)。通过NTSYS-pc 2.10e软件计算种质间的遗传相似性系数(Genetic similarity coefficients,GS),并利用非加权组平均法(Unweighted pair group method with arithmetic average,UPGMA)进行聚类分析。利用软件Structure 2.3.4分析老芒麦材料的群体结构和ΔK来确定最优分群数K,最终构建群体结构图[16]。
2 结果与分析
2.1 老芒麦形态指标的变异分析
2.1.1质量性状 对26份老芒麦5个质量性状进行遗传多样性分析结果如表1所示,结果表明,花序颜色遗传多样性指数最高,为 1.24,基生叶鞘被毛遗传多样性为0.72,叶片植株被粉为0.63,叶片颜色为0.49,杆生叶被毛为0.56。由表可知,在5个质量性状中,遗传多样性指数大小为花序颜色>基生叶鞘被毛>叶片植株被粉>杆生叶被毛>叶片颜色(表2)。
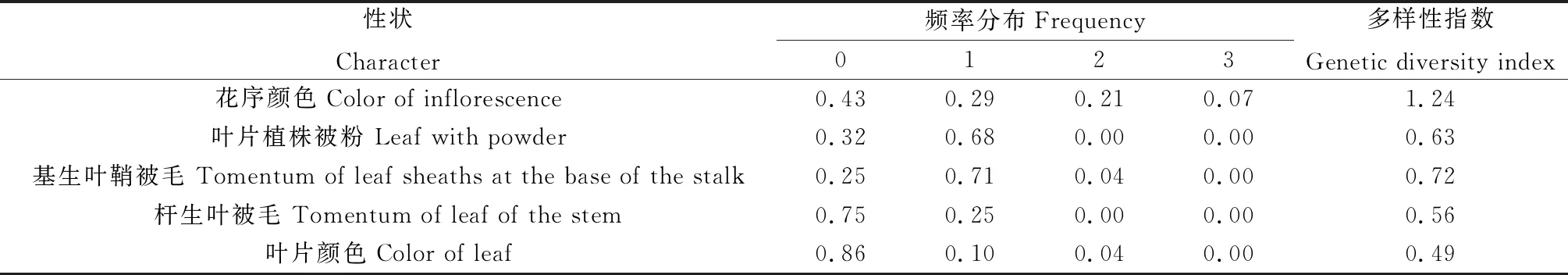
表2 老芒麦5个质量性状的遗传多样性
2.1.2数量性状 对26份老芒麦种质50个数量性状的分析表明(表3),每份材料各性状存在较大的变异。除花序中部穗节小穗数、花序底部穗节小穗数、上部稃片外稃长、上部稃片内稃长、中部稃片外稃长、中部稃片内稃长、下部稃片外稃长外,其余的性状的变异系数都大于10%,性状变异系数的范围为4.62%~45.93%,平均值为19.97%。其中,变异系数在30%以上的性状有9个分别为:上部颖片第二颖宽(33.10%),上部颖片第一颖芒长(33.66%),上部稃片外稃芒长(36.70%),中部颖片第二颖芒长(37.16%),下部颖片第一颖芒长(37.67%),下部颖片第二颖芒长(37.92%),上部颖片第二颖芒长(39.69%),中部颖片第一颖芒长(40.08%),花序上部穗节小穗长(45.93%),可见26份老芒麦种质变异主要来自于这9个性状。数量性状多样性指数的平均值约为1.80,其中农艺性状的遗传多样性指数最高是茎长(2.01),最低的是花序底部穗节小穗数(0.66)。
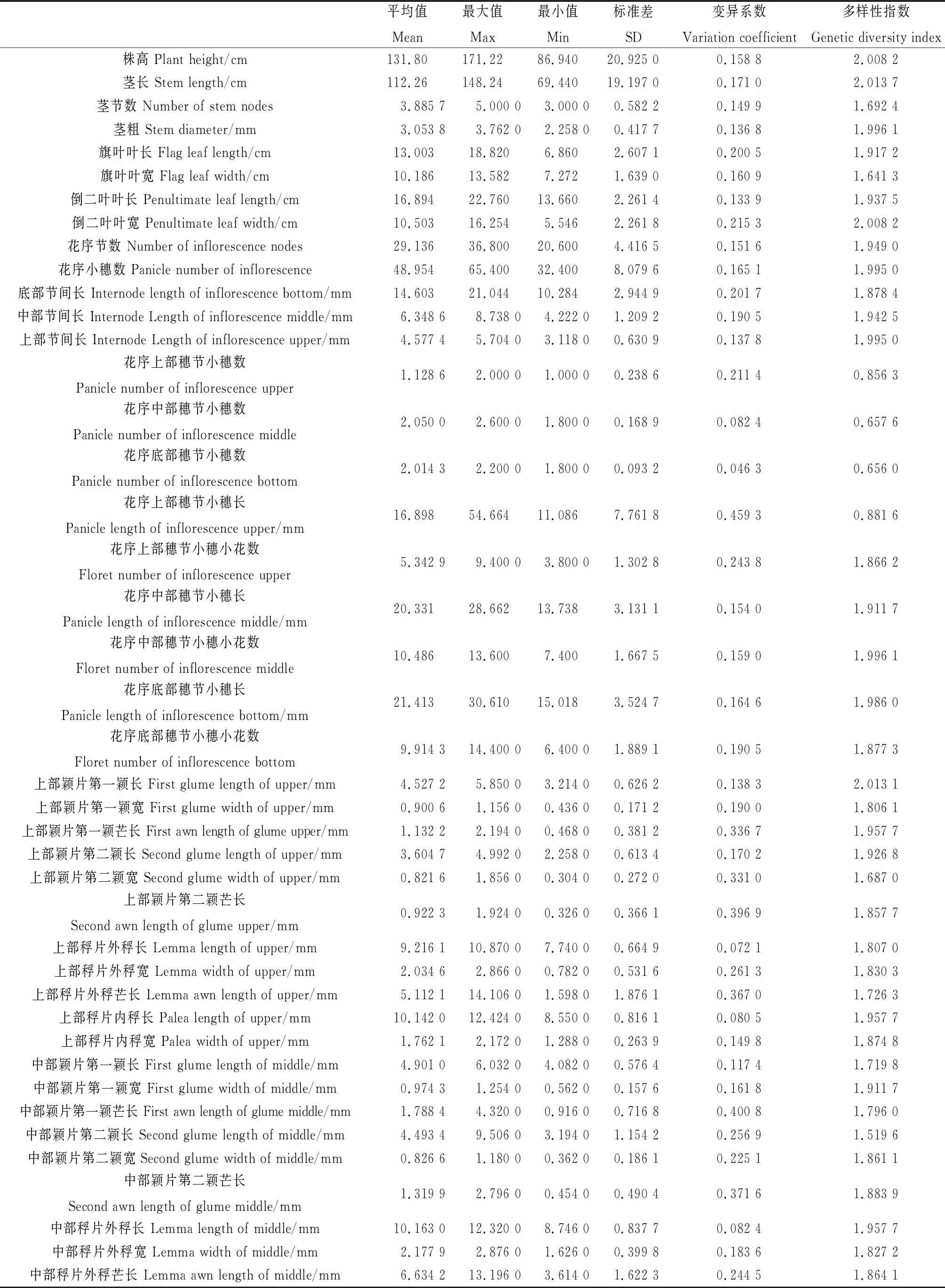
表3 老芒麦 50 个数量性状的遗传多样性分析
对材料间外稃芒长进行分析(图1),外稃芒长平均值‘Esxz’最大,材料8最小,将供试材料分为5个差异显著的组合(P<0.05),其中‘川草2号’及‘Esxz’外稃芒长平均值显著大于本研究24份短芒型老芒麦外稃芒长(P<0.05),且8,24,26这三份材料外稃芒长平均值小于其他材料(P<0.05)。表明供试的老芒麦种质具有芒短的稳定形态特性。
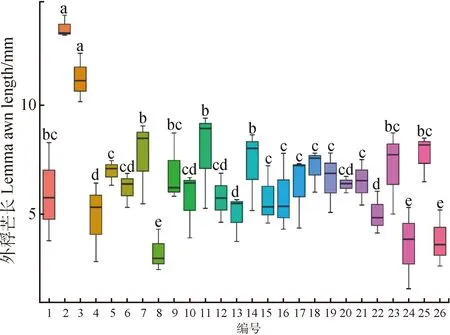
图1 26份材料外稃芒长的形态学特征
2.1.3基于农艺性状的聚类分析 聚类分析是研究种质资源分类的有效方法之一,性状越多,越能体现其实际情况,从而避免了仅以个别性状进行经验性分类的弊端。依据50个数量性状对26份种质材料进行聚类分析,可将供试种质分为五大类(图2)。类群Ⅰ包括7份材料,材料编号分别为21,15,9,13,5,20,12,占26.92%,在形态学上表现为,植株高大,花序节数多,穗长。类群Ⅱ包括4份材料,材料编号为11,7,19,17,占15.38%,在形态学上表现为,稃片和颖片发达。类群Ⅲ包括5份材料,材料编号为14,10,25,2,23,占3.846 %,在形态学上表现为植株茎秆粗、叶片长且宽、小穗数多,产量相关性状优越。类群Ⅳ包括5份材料,编号分别为26,8,4,1,24,占23.08%,在形态学上表现为内外颖、外稃长、芒长都是5个类群中最短,可以作为选育短芒型老芒麦的目标亲本加以利用。类群Ⅴ包括6,3,22,18,16,在形态学上表现为植株矮小。
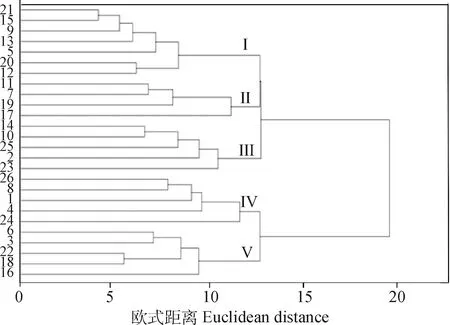
图2 26份材料的聚类分析
2.2 基于SCoT标记评价老芒麦种质遗传多样性
2.2.1SCoT引物多态性分析 利用14个SCoT引物共扩增出142条清晰条带,平均每个引物扩增10.1条,变幅为8条(SCoT7)~ 12条(SCoT38);多态性条带共96条,平均每个引物扩增多态性条带为6.9条,变幅为3(SCoT60)~10条(SCoT10),多态性比率(PPB)为67.61%;Ne变异范围为1.306~1.661,平均为1.498;H范围在0.165~0.325之间,平均为0.274;I范围在0.237~0.500之间,平均为0.397。这说明了SCoT标记对老芒麦具有良好的通用性与多态性,能够用于老芒麦种质的遗传变异分析(表4)。

表4 14个SCoT引物扩增结果
2.2.2聚类分析 基于DICE遗传相似系数构建了各供试老芒麦种质的UPGMA聚类树状图。如图3,在遗传相似系数为0.74处大致可将所有种质划分为3个类群,其中类群Ⅰ中包括材料26,3,13等,表明类群Ⅱ中的材料和审定品种‘川草2号’亲缘关系非常近;类群Ⅱ中包括材料7,6,5等材料;材料‘Esxz’与其他种质老芒麦的遗传距离最远,单独聚为第Ⅲ类。

图3 SCoT标记对26份老芒麦种质亲缘关系聚类图
2.2.3群体结构分析 根据Structure Harvester网页的分析结果表明,供试材料被划分为2个类群(K=2),分别标记为Q1,Q2并利用Structure绘制26份老芒麦种质的群体结构图(图4)。本研究参照刘丽华等[17]的划分方法,当Q≥0.6表明材料血缘相对单一,Q<0.6则具有混合来源。Q1类群(Q<0.6)中有17份材料,65%的材料拥有混合来源,遗传背景复杂;Q2类群(Q值>0.6)中有9份材料,表明材料来源单一,遗传背景简单,与其他类群基因交流较少。Q2类群(0.18)芒长平均数小于Q1类群(0.19),但Q2类群(3.50)芒长变异系数大于Q1类群(2.93);Q2类群(0.14)小穗平均数小于Q1类群(0.16),且Q2类群小穗数变异系数(48.69)小于Q1类群(49.69)。

图4 26份老芒麦材料群体结构分析
3 讨论
植物表型性状是其外部形态特征的综合体,能够直观的表现其遗传多样性。植物在适应和进化的过程中会产生表型性状的变异,增加群体表型的多样性。变异系数的大小与群体表型多样性丰富度呈正相关关系[18]。通过对植物表型性状的分析研究,能够在短时间内了解植物的遗传变异情况[19]。本研究将26份老芒麦种植于同一地点,使其生在相同的生境下,消除环境不同带来的影响。对24份野生短芒型老芒麦55个形态指标进行表型性状多样性研究结果表明,供试的种质具有丰富的形态学变异水平,质量性状的多样性指数在群体间的变化范围为0.49~1.24,数量性状的多样性指数在群体间的变化范围为0.66~2.01,表明供试老芒麦的数量性状比质量性状存在更大的变异,这与尹婷婷等[20]在老芒麦遗传多样性分析中的结果一致。鄢家俊等[21]在对野生老芒麦居群穗部形态数据的研究中发现,13个野生老芒麦的居群的芒长均大于1 cm,和本研究中两份对照老芒麦测量数据一致,供试短芒型老芒麦的外稃芒长均值为0.604 cm,这一性状显著小于两份对照材料,为短芒型老芒麦新品种的选育提供帮助。研究得到26份材料的数量性状变异系数的范围为4.62%~45.93%,相比于李瑶等[22]对59份老芒麦的研究变异系数更大,可能是本研究所测定形态指标数量更多,产生了更丰富的形态多样性,其中以中部颖片第一颖芒长、花序上部穗节小穗长最为突出,与芒相关性状的变异系数也较大。表明材料中芒长的可塑性强,对于已收集到的老芒麦材料应根据其不同的特性加以利用,通过杂交育种和合理的栽培管理措施能达到理想的选择效果。
遗传多样性是种质资源研究中的重要内容,本研究尝试使用SCoT分子标记,该标记比简单序列重复标记(Simple sequence repeat,SSR)、DNA扩增片段长度多态性(Amplified fragment length polymorphism,AFLP)、内部简单重复序列(Inter-simple sequence repeats,ISSR)成本低、多态性好,重复性高于随机片段多态性标记(Random amplified polymorphic DNA,RAPD)、相关序列扩增多态性标记(Sequence-related amplified polymorphism,SRAP)[23-24]。本研究的结果表明,SCoT标记可以有效地评估老芒麦种质的遗传多样性,该标记的多样性数据高于之前报道的SRAP(I=0.2267,H=0.1508)[25],SSR(I=0.1930,H=0.1296)[25],ISSR(I=0.269,H=0.181)[26]和RAPD(I=0.263,H=0.176)[27]。说明SCoT引物对老芒麦种质具有好的扩增效果,能在野生老芒麦种质中检测出较丰富的遗传多态性,将其用于短芒型老芒麦种质资源种质鉴定、遗传多样性分析等研究是可行的。群体结构分析可以用于评价野生老芒麦种质的遗传背景与亲缘关系,对老芒麦杂交育种中的亲本选配具有重要的指导作用。群体遗传结构分析表明,65%的老芒麦种质资源拥有混合来源,同一地区和类群间的野生老芒麦资源基因间存在着较为广泛的交流。将综合本研究中形态指标对应群体结构来讨论发现,Q1类群芒短且Q1类群产量相关性状更高,与Ntakirutimana等[28]研究结果一致,即芒长的增加导致每单株小花数减少,芒长与种子产量相互影响的结论一致,因此芒长这一性状是影响种子产量的重要因素之一,短芒型材料的选育是高产育种的目标亲本。
使用形态标记对种质资源进行研究长久以来是最有效的途径之一,但环境因素及遗传因素制约着表型性状的表达,具有一定的局限性。然而,消除环境变异的情况下,基于表型性状的聚类和基于SCoT标记的聚类不能完全对应,造成这一结果的原因可能是:(1)表型性状的检测水平有限,且植株生存环境的细微环境也会对其表型造成影响[29];(2)本研究所选SCoT引物不能将整个基因组的变异情况揭示完全,且有些发生在DNA水平的变异和表型性状的改变没有因果关系。尽管如此,本研究通过结合表型性状与基因型变异分析,为其种质资源提供综合性评价,对加强短芒型老芒麦材料选育提供基础。
4 结论
本研究采用形态学和SCoT分子标记揭示了供试的短芒型老芒麦种质具有丰富的遗传变异;其中材料Esdm24,Esdm6,Esdm22这3份种质的芒长及芒相关性状较为优异,可以作为短芒型老芒麦新种质创制和品种选育的优异种质。
猜你喜欢
今日农业(2022年13期)2022-09-15 01:18:00
草业科学(2021年5期)2021-06-18 04:02:12
作物杂志(2020年6期)2020-04-18 04:37:08
江苏农业科学(2019年5期)2019-09-02 14:01:46
中国畜禽种业(2019年7期)2019-01-06 08:43:59
中国麻业科学(2018年6期)2018-04-09 11:22:12
广东农业科学(2017年5期)2017-08-29 10:37:31
西南农业学报(2016年5期)2016-05-17 05:42:21
广西林业科学(2016年3期)2016-03-16 05:43:21
动物营养学报(2015年10期)2015-12-01 02:26:36
