
水体消毒过程中活的不可培养细菌的形成与复苏机制研究进展
2023-10-07 08:38:10李桂英刘建莹安太成
生态环境学报 2023年7期
李桂英,刘建莹,安太成
广东工业大学环境科学与工程学院/环境健康与污染控制研究院/环境催化与健康风险控制重点实验室/粤港澳污染物暴露与健康联合实验室,广东 广州 510006
有效的水消毒技术对供水质量和人类健康保护至关重要(Cai et al.,2021)。其中,光催化技术具有高效、低毒、无选择性等优点(阳海等,2010)。然而,亚致死水平消毒技术却不能完全去除水中的细菌,因为病原菌能够通过进入休眠状态来逃避消毒技术造成的损害(Cai et al.,2022)。在细菌的休眠状态中,活的不可培养(Viable but nonculturable,VBNC)状态细菌是一个典型的休眠亚群(Oliver,2005)。VBNC细菌存在可能会对饮用水安全、食品安全和生物安全控制均构成一定程度的威胁。值得注意的是,细菌休眠是细菌获得抗生素耐药性的重要途径(Wang et al.,2023a)。抗生素耐药性细菌(ARB)也能进入VBNC状态以避免被消毒技术杀死(Guo et al.,2019a)。ARB能够逃脱抗生素治疗并造成严重疾病,因此VBNC ARB会增加抗生素耐药性风险(Zhong et al.,2022)。
然而,目前却很少有关于消毒技术诱导形成的VBNC细菌发生复苏的相关研究。到目前为止,仍不清楚消毒技术诱导形成VBNC细菌复苏的潜在分子机制。因此,本综述结合课题组在抗生素耐药菌方面的消毒控制机制和细菌休眠方面的研究及其文献报道的综述,详细地介绍了消毒技术如何诱导VBNC细菌的形成。同时,本综述也梳理了VBNC细菌的复苏方法,并阐述了自然复苏、复苏促进因子(Rpf)复苏与自诱导剂复苏的潜在的复苏机制,期望为水处理过程中病原微生物的安全性评估和灭活技术制定更有效的消毒策略提供理论依据。
1 VBNC细菌的形成
在水消毒过程中,细菌的各种应激反应会被激活以保护细菌免受损害和死亡。ARB更能抵抗刺激产生的损伤,因为抗生素耐药作用靶点能够影响应激反应,从而加速细菌细胞的修复(Liu et al.,2022)。也就是说,细菌能够接受亚致死水平的氧化压力(Ji et al.,2022)。亚致死光催化消毒会导致抗生素耐药性的长期维持和通过基因水平转移方式促进抗生素耐药性的传播(Yin et al.,2019)。亚致死消毒无法完全消除ARB,甚至会诱导VBNC状态的形成(Cai et al.,2022)。VBNC细菌普遍存在于在水体中,尤其是生物膜中(Chen et al.,2022)。然而,用于去除水传播致病菌的传统水消毒技术可以诱导细菌进入VBNC状态,包括紫外线消毒(Zhang et al.,2015)、氯和氯胺消毒(Chen et al.,2018)、臭氧消毒(Jäger et al.,2018)等。另外,消毒技术对抗生素耐药基因(ARGs)的去除效果有限(陈蕾等,2018)。光电催化会诱导ARB细菌从可培养状态转变为VBNC状态,这会导致ARGs在水体中残留在和增加继续扩散的潜在风险(Qi et al.,2022)。具有强耐受性的VBNC细菌可能无法被传统的水消毒技术完全消除(Cai et al.,2022)。完成消毒的饮用水中也检测到了处于VBNC状态的病原体(Guo et al.,2021)。因此水处理系统中的VBNC细菌可能会构成巨大的健康风险,因为它们无法被传统的异养平板计数方法检测到,这导致了总的活细菌数量被明显低估(Dong et al.,2020)。进入VBNC状态后,病原体对多种抗生素具有更强的耐受性(Lin et al.,2017)。进入休眠后,VBNC细菌可通过不同的耐药机制增加对抗生素的耐药性,比如增加外排泵的表达(Yin et al.,2020)和降低代谢活性(Qi et al.,2022)。此外,VBNC细菌可能会引起生素耐药性的传播和造成慢性感染。亚致死消毒诱导了VBNC状态的形成,还可能会导致抗生素耐药性的进一步传播和发展(Ahmed et al.,2021)。这是因为亚致死刺激会引起细菌的氧化应激和促进外排泵基因的表达上调(Yin et al.,2020)。在暴露于亚致死刺激下,细菌会进入休眠状态,其外排泵的活性也会增强,这有助于细菌存活并将进入胞内的抗生素泵出细胞外以降低细菌体内药物浓度,从而导致细菌的抗生素耐药性的增加(Liu et al.,2023)。另外,从休眠状态复苏的ARB可以通过结合转移等方式恢复ARGs的产生及其传播(Wang et al.,2023a)。
消毒可以诱导细胞内活性氧(ROS)的生成(Li et al.,2020)。环境压力会引起细菌的氧化损伤,包括蛋白质损伤、DNA损伤、RNA损伤,从而阻止细菌的正常生长(Chen et al.,2019)。细菌可以通过表型的变化来响应环境的变化和提高细菌的存活率。VBNC细菌的生理特性发生了很大改变。与长杆状的正常细菌相比,VBNC细菌大多表现为短杆状或圆球状,仍保持低代谢活性和毒性(Alvear-Daza et al.,2021)。如果致病性持续存在,VBNC状态下的病原体可能会构成未识别的公共卫生威胁(Zhang et al.,2015)。因为能量限制,细胞生长和死亡、细胞运动、细胞群落、运输和分解代谢等细胞等过程被抑制,从而导致细胞生长、运动和分裂的障碍,这可能导致其不能在平板培养基上形成菌落(Zhu et al.,2022)。信号转导和膜传输过程等耗能活动在VBNC细胞中被显著抑制以节省能量(Liao et al.,2020)。
目前,有少许关于VBNC状态的潜在的形成机制的研究(图1)。环境刺激会增加ROS水平,这会造成DNA、蛋白质和脂质损伤和会诱导SOS反应(Zhao et al.,2014)。当产生的ROS多于消除能力时,氧化应激随之而来。在暴露于消毒后,细菌与氧化应激、细胞修复、DNA修复以及转移调节相关的一系列应激反应被触发,这有助于细菌应对氧化损伤和提高存活率(Yin et al.,2021)。一般应激反应是由聚合酶σS(RpoS)和LysR转录调节因子(OxyR)调控的(Dalebroux et al.,2010)。RpoS赋予了细菌对各种环境压力源的耐受性,而OxyR主要参与使细菌能够应对氧化应激的过程(Liao et al.,2020)。因此,RpoS基因对于激活VBNC细菌的形成很重要,其是调节胁迫反应因子的主要信号。
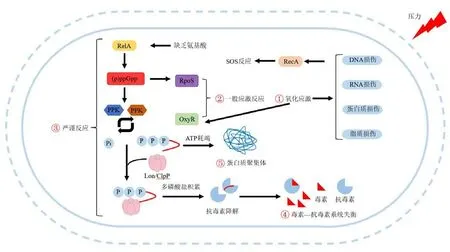
图1 VBNC细菌的形成机制Figure 1 Formation mechanism of VBNC bacteria
严谨反应也在VBNC细菌的形成中起着重要作用,而过度产生ppGpp的细菌更有可能被诱导到VBNC状态(Boaretti et al.,2003)。这是因为环境胁迫会诱导严谨反应以增强细菌对环境胁迫的抗性。但是,VBNC细菌增强了对环境胁迫的抗性却付出了相应的代价。在严谨反应中,由relA基因编码合成的(p)ppGpp会影响ATP合成过程,进一步抑制DNA复制、细胞分裂、转录和翻译活动以降低能量的消耗(Ayrapetyan et al.,2018)。另外,(p)ppGpp能够促进RpoS的合成,从而调控一般应激反应(Wang et al.,2019)。此外,毒素-抗毒素系统体系受到严谨反应的调控。严谨反应中的(p)ppGpp会导致多磷酸盐的积累,从而激活Lon蛋白酶/ClpX蛋白酶并降解抗毒素(Ayrapetyan et al.,2018)。因此,更多的游离毒素积累,这扰乱了毒素-抗毒素系统的正常比例和导致细菌的生长被抑制(Harms et al.,2018)。毒素-抗毒素系统的失衡会促进细菌进入VBNC状态(Zhang et al.,2020)。
VBNC细菌的生存策略是减少代谢活动(Oliver,2005)。ATP的合成对于修复细胞损伤和细菌生长是必要的能力,但ATP含量的降低会意味着能量代谢途径被阻断(Sun et al.,2017)。ATP的耗竭会导致蛋白质聚集体产生,这会导致细菌进入深度休眠状态(Pu et al.,2019)。比如非热等离子体细菌技术通过代谢抑制和产生氧化应激反应诱导金黄色葡萄球菌的VBNC状态(Liao et al.,2020)。
总的来说,VBNC状态不是单一因素形成,而是由应对氧化应激的一般应激反应、修复DNA损伤的严谨反应、失衡的毒素-抗毒素系统、ATP的耗竭和蛋白质聚集体的产生的全局调控共同促进暴露于消毒的细菌进入了VBNC状态。
2 VBNC细菌的复苏
VBNC细菌可以在外部压力源消失时(例如,在水储存和分配期间)从VBNC状态转变为正常的可培养状态,即发生复苏(Zhang et al.,2018)(图2)。只有VBNC细菌具有复苏能力时,进入VBNC状态才可以被认为是细菌在压力下的一种生存策略(Bodor et al.,2020)。复苏过程可能是一个复杂的生理过程,涉及各个通路的调控,这意味着细胞的生理和代谢过程的恢复到正常水平(Pan et al.,2023)。VBNC细菌的形态恢复是一个重塑的过程,即从VBNC状态复苏过程中细胞分裂能力的恢复需要细胞质蛋白和细胞壁肽聚糖的重新合成(Pan et al.,2023)。
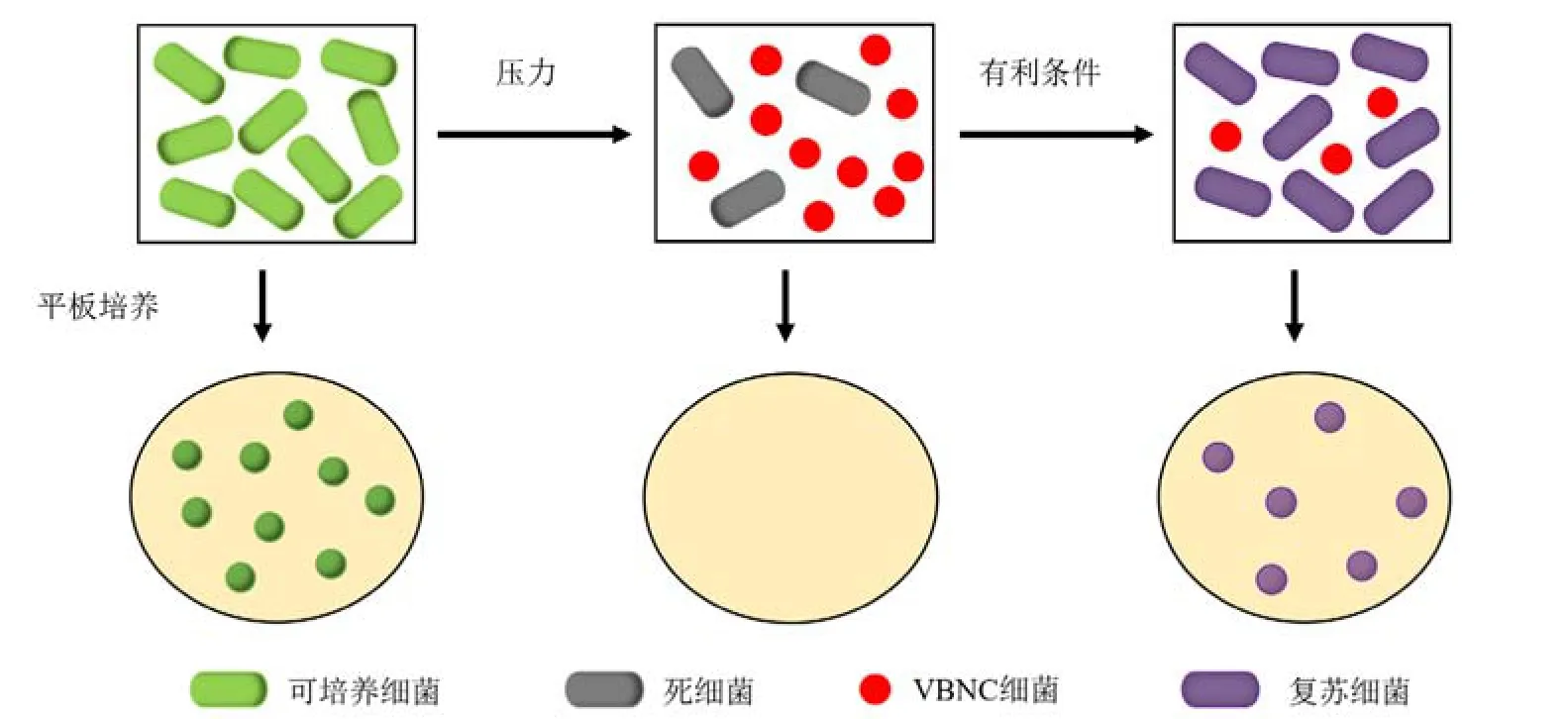
图2 VBNC细菌的复苏示意图Figure 2 Resuscitation diagram of VBNC bacteria
未在水消毒中去除的VBNC细菌可能会在运输过程中复苏,而这些复苏的细菌很可能通过饮用水的途径进入人体并可能产生严重疾病(Liu et al.,2009)。因此,VBNC抗生素耐药菌能够复苏是一个非常重要的公共卫生威胁(Zhu et al.,2022)。发生复苏后,恢复可培养能力的细菌导致大多数蜡蛾幼虫感染疾病并造成其死亡,这说明了VBNC细菌在复苏后恢复了其强致病性(Zhu et al.,2022)。VBNC细菌在恢复可培养能力后仍表现出较好的结合转移能力,这说明了复苏细菌的抗生素耐药性风险依然存在(Zhu et al.,2022)。
然而,仍不清楚促进VBNC细菌复苏和致病性恢复的因素及其复苏机制和缺乏控制VBNC致病菌复苏的有效手段。因此,需要进行VBNC细菌的复苏机制研究来制定更有效的消毒方法以预防或控制VBNC细菌的形成和复苏。异养菌平板计数通常用于监测细菌的消毒效果,VBNC致病菌由于其不可培养特性造成了消毒效果的误判。因此,复苏研究对于饮用水中微生物的安全性评估来说是非常必要。监测在水消毒技术下形成VBNC致病菌的复苏有利于确定该消毒技术的强度和处理时间是否足以完全杀灭水环境中的细菌,这有助于制定更有效的消毒策略。同样在临床医疗环境中,细菌可以通过进入休眠状态来逃脱宿主的免疫系统和抗生素治疗(主要针对活跃生长的细菌),但是这些休眠细菌会复苏,造成疾病的复发(Epstein,2009)。因此很有必要研究休眠细菌复苏的分子机制,这非常有助于研究人员设计一种可能的方法来降低休眠细菌复苏造成疾病复发的频率和进行有效抗菌。
3 VBNC细菌的复苏方法进展
VBNC细菌对有利的环境和条件下刺激做出反应,变得代谢活跃且可培养(Pan et al.,2023)。目前复苏VBNC细菌的方式各种各样,见表1。总体来说,富含营养的培养基对于细菌的复苏是优先的刺激。在添加了LB肉汤后,VBNC大肠杆菌发生了复苏(Zhang et al.,2018)。升高温度能促进低温诱导的VBNC细菌发生复苏(Wei et al.,2018)。复苏促进因子能够增加细菌的活力并促使细菌从VBNC状态转变成可培养状态(Ye et al.,2020)。此外,添加一些促进生长的物质也可以帮助VBNC细菌复苏,包括过氧化氢酶、蛋白酶、YeaZ蛋白、氨基酸、丙酮酸(Ramamurthy et al.,2014)。群体感应自诱导剂AI-2可复苏VBNC创伤弧菌。值得注意的是,VBNC细菌在进入宿主体内后恢复了可培养能力(Hwang et al.,2006)。在河口环境中进行原位复苏(添加营养和原位培养)治疗后,VBNC创伤弧菌恢复了可培养能力和katG表达(Oliver et al.,1995a)。当紫外线辐射诱导细菌形成VBNC状态后,可在适当的温度(37 ℃)和营养物质(LB肉汤)下,使得铜绿假单胞菌和大肠杆菌都可以发生复苏,但大肠杆菌表现出比铜绿假单胞菌更强的复苏能力(Zhang et al.,2015)。氯诱导的VBNC大肠杆菌在37 ℃时可以在LB肉汤中复苏(Chen et al.,2018)。上述的复苏条件与人体体内环境相似,这就揭示了消毒技术诱导的VBNC细菌也是可以在人体内恢复培养能力而具有潜在的健康风险。VBNC细菌代表了绝大多数有效的微生物资源,其在生物治疗中具有重要意义(Su et al.,2013)。总的来说,复苏VBNC细菌的研究大多是复苏条件的探索,包括原位复苏(Nichols et al.,2010)。但目前仍缺乏VBNC细菌复苏机制方面的研究。因此我们接下来重点对复苏机制进行详细的综述。
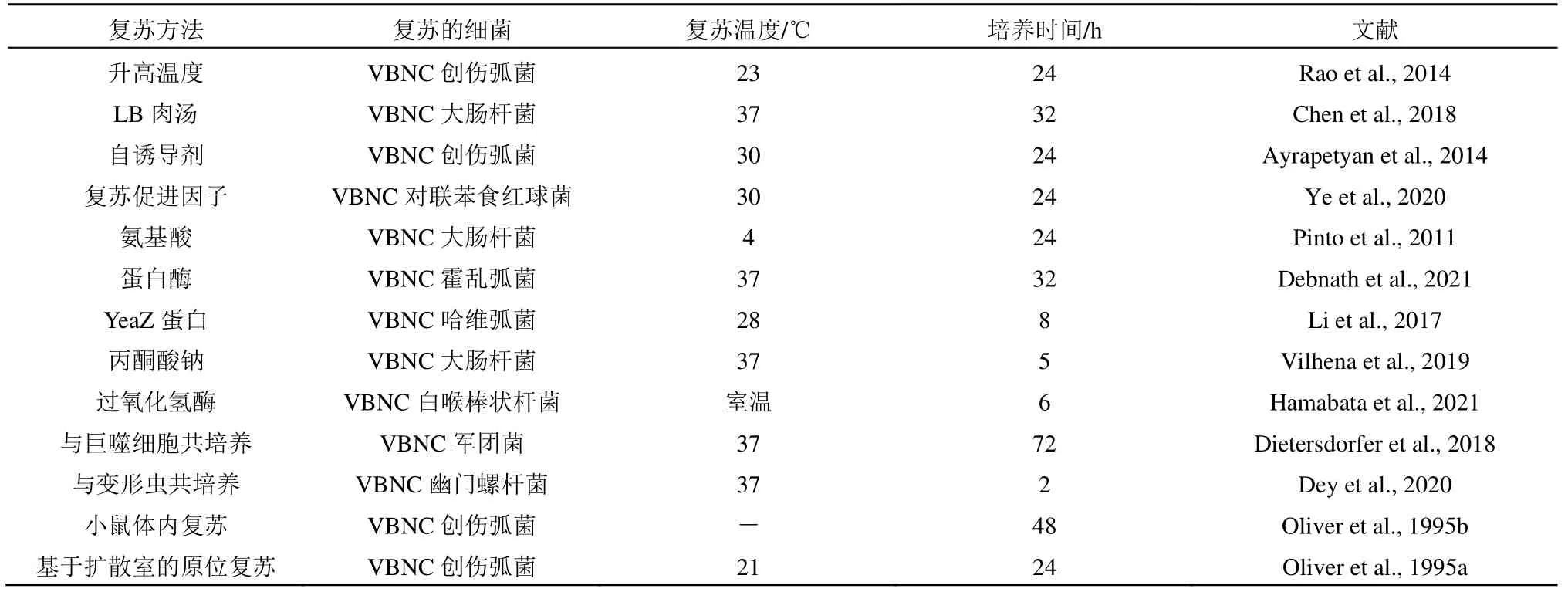
表1 复苏的VBNC细菌的方法Table 1 Resuscitation method of VBNC bacteria
4 VBNC细菌的复苏机制
消毒过程诱导的VBNC细菌会进入到水体环境中,有可能通过自身修复来进行复苏。在水体环境中,其他细菌分泌的自诱导剂-2可以促进这些VBNC细菌复苏。藤黄微球菌能够分泌Rpf,这使得VBNC细菌可以在Rpf作用下恢复可培养能力。
4.1 VBNC细菌的自然复苏潜在机制
消毒技术会引起不同细胞水平的损伤,包括膜损伤、蛋白质碎裂、ATP生成抑制和核酸损伤(Yoon et al.,2021)。细菌已经发展出氧化还原平衡系统,以防止活性氧(ROS)损伤。但是在环境刺激下,细菌内的ROS对脂质、蛋白质、RNA、DNA和细胞膜造成的损伤会超过细菌的修复能力时,就会发生氧化应激。氧化应激攻击细菌细胞成分,这会导致细胞损伤甚至死亡(Sun et al.,2017)。受损的DNA无法被替换,需要修复才能保持活力(Lee et al.,2019)。为了存活并适应环境刺激,细菌会发生严谨反应并关闭大量基因的表达并伴随着表型的变化(Irving et al.,2021)。细菌的新陈代谢和所利用的蛋白质去响应环境的变化和提高细菌的存活率(Yang et al.,2021)。SOS反应可以修复消毒过程中细菌的DNA损伤并促进细菌存活,但会抑制细胞分裂(Dörr et al.,2009)。还原系统能够修复细胞质和细菌细胞膜中氧化损伤的半胱氨酸和蛋氨酸残基(Ezraty et al.,2017)。
VBNC细菌能够复苏的前提就是要修复这些损伤,恢复氧化还原平衡和代谢活性。因此,我们综述了VBNC细菌发生自然复苏的潜在机制(图3)。为了使细胞离开休眠状态并恢复活力,首先需要清除蛋白质聚集体和恢复蛋白稳态(Pu et al.,2019)。严谨反应的过程中,细菌通过仅仅维持最低量的活性来节约其资源,直到条件改善时,它们可能又会恢复正常代谢(Wang et al.,2023b)。通常,细菌的应激反应允许细胞适应并从特定应激中恢复。光消毒诱导的DNA损伤可以根据消毒后条件(比如合适的光照条件)进行修复(Sousa et al.,2017)。有文献报道UVC诱导形成的VBNC大肠杆菌确实发生了复苏(Zhang et al.,2018)。VBNC细菌能够修复亚致死损伤并恢复可培养能力。但是,随着紫外线剂量的增加和暴露时间的延长,恢复速度似乎越来越慢(Suss et al.,2009)。也就是说,光的持续刺激会对细菌造成严重的损伤以至于细菌很难恢复(Karaolia et al.,2018)。

图3 VBNC细菌的自然复苏潜在机制Figure 3 Potential resuscitation mechanism by natural resuscitation
总体而言,复苏能力取决于外部应激强度和VBNC状态的持续时间(Pinto et al.,2011)。在不同强度的外应激下,细菌损伤的程度是不同的,因此休眠的深度也不一样(Yin et al.,2022)。随着细菌的ATP浓度的降低,高压二氧化碳技术处理后VBNC比值增加,这表明了ATP含量与VBNC比值呈高度负相关(Yang et al.,2023)。VBNC细菌进入休眠所需的时间不同,因此休眠的深度也可能是不同的(Wang et al.,2010)。在相同的处理条件下,同一批次下的VBNC细菌并非都会发生复苏(Chen et al.,2018)。在休眠的初始阶段,这些休眠细菌可以在消除压力后迅速复苏。然而,如果细菌暴露在长期的压力下,会积累更多的损伤,使得休眠程度增加,代谢活性进一步降低,那就是细胞需要更多时间来复苏。一些VBNC细菌无法复苏,可能因为损伤程度超出了细胞恢复生长的能力(Ayrapetyan et al.,2015)。VBNC细菌能否复苏也取决于体内的能量储备。当ATP得到补充,细菌准备复苏生长时,还需要蛋白DnaK和ClpB来完成蛋白质聚集体的解聚(Pu et al.,2019)。撤去应激后,VBNC细胞对有利的环境条件做出反应,然后变得代谢活跃且可培养(Zhang et al.,2015)。复苏过程是由侦察细胞退出休眠而激活的,这些细菌可能是休眠深度较浅的VBNC细菌(Epstein,2009)。如果环境有利,侦察细菌开始繁殖,可以向其他细菌发出信号并促进其他细菌复苏(Epstein,2009)。细菌复苏有最佳的时间(复苏窗口),能量会随着VBNC状态持续时间的延长而消耗,复苏能力逐渐变弱(Pinto et al.,2015)。一旦时间过长,能量耗尽,细菌就可能会失去复苏的能力。在进入VBNC状态22 d后,通过直接活菌计数法检测到的活细胞数量显著减少了(Masmoudi et al.,2010)。这可能是因为休眠细菌无法修复永久性损伤,随之因为能量耗尽而发生死亡。
4.2 VBNC细菌的Rpf复苏潜在机制
细胞壁的改变是休眠细菌的共同特征。进入VBNC状态后,细菌的平均细胞壁变厚了,这有助于细菌增强对环境压力的耐受性(Rittershaus et al.,2013)。此外,VBNC细菌的细胞壁的化学性质也发生了改变。例如,乙酰化的增加、肽聚糖的交联和脂蛋白共价结合多肽的数量等(Jia et al.,2020)。虽然变厚的细胞壁是抵抗环境压力的有效屏障,但这可能不利于细胞生长。因此,细胞壁的重塑对于休眠细胞恢复到活跃生长状态是必不可少的(Sexton et al.,2015)。在进入VBNC状态和复苏过程中,调控肽聚糖合成和形态重塑的相关基因的表达发生变化,从而导致形态变化,也就是说,细胞壁的改变在VBNC状态的维持和退出中起着至关重要的作用(Jia et al.,2020)。
当这些休眠细胞恢复生长时,可能需要调整肽聚糖的种类以进行细胞分裂(Boudreau et al.,2012)。现有研究表明休眠细胞的激活需要肽聚糖水解,这会改变细胞壁的机械特性以促进细胞分裂或释放起抗休眠信号作用的裂解产物(Keep et al.,2006)。值得注意的是,VBNC细菌可以被一种溶菌酶样蛋白重新激活,然后发生复苏(Cohen-Gonsaud et al.,2005)。当Rpf添加到VBNC/休眠微生物的培养物中时,即使在皮摩尔浓度下,含Rpf的细菌培养上清液也是可以刺激细胞生长的(Cohen-Gonsaud et al.,2005)。细胞壁衍生的多肽可以刺激大肠杆菌和铜绿假单胞菌的生长恢复(Jõers et al.,2019)。Rpf有类似于溶菌酶的折叠结构,能够裂解细胞壁肽聚糖,从而产生可以作为生长启动信号分子的多肽(Mukamolova et al.,2006)。细胞壁衍生的多肽可以充当信使,然后促进VBNC细菌退出休眠状态(Boudreau et al.,2012)。这可能是其观察到的细菌复苏和刺激其增殖的原因,也就是说这可能是Rpf复苏VBNC细菌的潜在机制(图4)。目前Rpf蛋白如何促进VBNC细菌退出休眠的机制有两种说法:(1)Rpf具有细胞壁裂解活性,能够降低因为进入VBNC状态而改变的细胞壁结构施加的物理限制,从而使得VBNC细菌退出休眠;(2)Rpf进入细菌体内,细菌感知Rpf的信号,刺激细菌恢复生长(Dworkin et al.,2010)。简单概括Rpf复苏VBNC细菌的潜在机制:Rpf在整个细菌生长过程中重塑细胞壁,促使VBNC细菌从“不可培养”状态恢复到活跃生长的可培养状态。
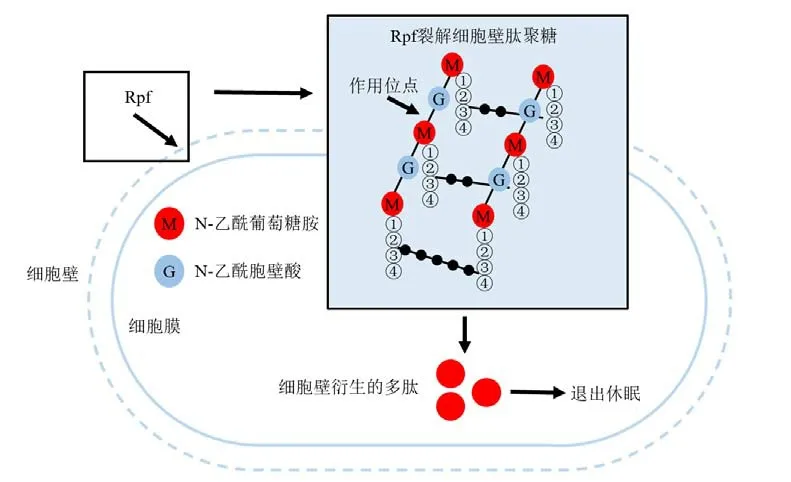
图4 VBNC细菌的Rpf复苏潜在机制Figure 4 Potential Mechanism of Resuscitation by Rpf
4.3 VBNC细菌的自诱导剂复苏潜在机制
自诱导剂是由微生物产生并分泌到环境中被群体中的细菌感知的小分子(Valastyan Julie et al.,2021)。群体感应信号对于复苏过程至关重要。群体感应自诱导剂能够促进环境水样中休眠的霍乱弧菌发生复苏(Bari et al.,2013)。当环境条件允许时,群体感应信号分子自诱导剂2(AI-2)能够介导微生物种群中的细胞间通讯,并通过“群体感应”触发相关基因表达,从而触发VBNC细菌的复苏(Ayrapetyan et al.,2014)。在暴露于含有AI-2的培养基后,自然环境中的VBNC霍乱弧菌的复苏会迅速发生,其细胞形态从圆球状恢复到杆状形状(Ayrapetyan et al.,2014)。如果水体环境中的细菌分泌大量自诱导剂,那么水环境中的休眠霍乱弧菌可能会在此期间复苏并更容易引起人类疾病。
根据先前的研究,VBNC创伤弧菌不能够在实验室培养基上培养的原因是过氧化氢酶(CAT)活性过低(Kong et al.,2004)。CAT活性的降低是因为katG的转录被抑制。但是人为添加CAT可以帮助细菌从VBNC状态中复苏(Hamabata et al.,2021)。蛋白质RpoS调控CAT的产生,而LuxR能够增强RpoS的表达,这对VBNC细菌的复苏来说是很重要的(Joelsson et al.,2007)。添加了AI-2后,野生型VBNC细菌能够发生复苏,然而RpoS突变体(不能表达RpoS基因)的VBNC细菌并不能发生复苏(Ayrapetyan et al.,2014)。当环境中AI-2进入到VBNC细菌体内,VBNC细菌会做出一系列响应。LuxR作为受体与AI-2结合,然后刺激VBNC细菌的RpoS的表达,随后诱导katG表达来增强CAT的活性,降低过氧化氢毒性,从而使生物体从VBNC状态恢复到可培养状态(Ayrapetyan et al.,2014)。这可能是自诱导剂复苏VBNC细菌的潜在机制(图5)。
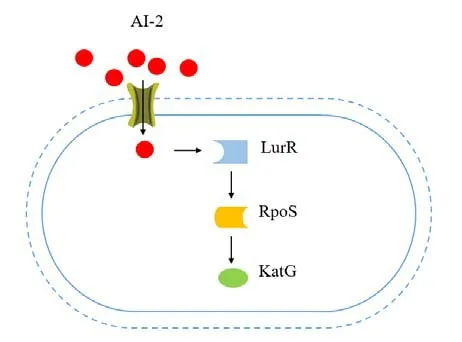
图5 VBNC细菌的自诱导剂复苏潜在机制Figure 5 Potential resuscitation mechanism by self-inducer
5 展望
VBNC细菌的复苏研究结果对饮用水的微生物安全性来说非常重要。但是,评估VBNC细菌复苏所带来的潜在健康风险还不够全面,还未在复苏细菌的抗生素耐药性、毒性表达和致病性方面进行深入研究。而且,进入/退出VBNC状态对细菌的抗生素耐药性的发展和传播的具体机制仍不清楚。VBNC细菌在复苏后发生自然转化、转导的频率等需要进一步研究,这非常有助于更全面地评估VBNC细菌复苏所带来的潜在健康风险。
最近的研究对“多大剂量的消毒技术能够灭活VBNC细菌并避免其复苏”、“复苏的VBNC细菌生理和代谢过程是否都恢复到正常水平”等问题的回答比较模糊。目前,在消毒技术下形成的VBNC细菌的复苏机制的研究仍然比较缺乏。因此,仍需要深入研究在复苏过程细胞的生理特性和代谢过程的恢复程度,然后从基因层面阐述VBNC细菌复苏的潜在机制。微流控技术的延时成像能够提供了大量图像数据,这有助于进行深度学习分析并解析单细胞水平的微生物行为(Kasahara et al.,2023)。因此,可以开发更准确的分析方法来实现在单细胞水平上可视化细菌复苏的过程并利用微流控技术分析细菌的生理特性。
人们对VBNC细菌和复苏细菌的特定代谢特征仍然知之甚少。与可培养细菌相比,VBNC大肠杆菌的代谢发生了变化,这是VBNC细菌减少能量消耗的生存策略(Wang et al.,2023c)。代谢组学可作为新的技术手段来揭示复苏细菌的标志性代谢产物和代谢途径。代谢途径的动态调节可以通过稳定同位素示踪来监测(Trefely et al.,2019)。稳定同位素示踪代谢组学方法可以通过测量同位素标记的代谢物来定量揭示代谢活动(Wang et al.,2022)。VBNC细菌和复苏细菌的特定代谢特征和标志物可以通过质谱和同位素示踪技术直接检测,特别是单细胞代谢分析提供了强有力的技术支持。CTC-流式细胞仪检测技术(CTC-FCM)可以测定VBNC细菌的呼吸活性(Guo et al.,2019b)。可以运用CTCFCM来监测复苏过程细菌的呼吸活性变化并开展细菌代谢网络的动态波动过程及调控机制研究。单细胞拉曼光谱显示出更多的异质代谢活性分布,而一些细胞甚至进入代谢“沉默”(Bao et al.,2023)。重水标记结合拉曼光谱仪可以用来监测复苏过程中单个细胞的代谢活性的恢复程度。荧光能量共振转移技术可以在单细胞水平上定量监测实时监测细菌胞内代谢产物浓度(Bi et al.,2023)。也就是说,这种技术可以监测细菌从VBNC状态转变为复苏状态的代谢过程。
猜你喜欢
小哥白尼(趣味科学)(2021年10期)2022-01-17 02:42:14
数学物理学报(2021年4期)2021-08-30 08:27:48
中老年保健(2021年6期)2021-08-24 06:53:34
新世纪智能(数学备考)(2020年10期)2021-01-04 00:37:50
学生天地(2020年14期)2020-08-25 09:20:56
学生天地(2020年10期)2020-08-25 09:14:34
科技知识动漫(2017年12期)2018-03-07 16:46:23
中成药(2017年12期)2018-01-19 02:06:52
中国交通信息化(2017年8期)2017-06-06 07:16:47
国外医药(抗生素分册)(2016年1期)2016-07-10 12:02:35
