
稻田土壤氮循环与砷形态转化耦合的研究进展
2023-10-07 08:38朱忆雯尹丹胡敏杜衍红洪泽彬程宽于焕云
生态环境学报 2023年7期
朱忆雯,尹丹,胡敏,杜衍红,洪泽彬,程宽,于焕云*
1.广东工业大学环境科学与工程学院,广东 广州 510006;2.广东省科学院生态环境与土壤研究所,广东 广州 510650;3.长江大学农学院,湖北 荆州 434000;4.常州大学环境科学与工程学院,江苏 常州 213164
砷是一种剧毒的类金属物质,普遍存在于自然环境中,长期接触会对人类健康产生极大威胁。农业环境中,利用砷污染地下水灌溉农作物是导致土壤与粮食砷含量超标的主要原因之一。与其他农作物相比,水稻更容易吸收土壤中的砷,导致稻米中砷超标,并通过食物链进入人体可造成慢性中毒,引发多种疾病(Sharma et al.,2021)。水稻作为最主要的农作物之一,全球一半的人口以稻米为主食,随着全球人口不断增加,水稻的需求也在增长(Bao et al.,2022),因此稻田砷污染问题备受关注。
稻田土壤中砷主要以无机砷(三价砷As(Ⅲ)、五价砷As(Ⅴ))存在,其次为有机砷(一甲基砷酸(MMA(Ⅴ))、二甲基砷酸(DMA(Ⅴ))等)。稻田土壤中砷转化一般包括砷氧化、砷还原、砷甲基化和脱甲基化等过程,主要是由微生物驱动,且受多种因素影响(Yamaguchi et al.,2011;Yang et al.,2022)。稻田常常施用大量氮肥,但仅有35%氮肥被水稻所吸收利用,过量施用氮肥可造成环境氮污染(朱兆良,2008)。稻田氮循环主要包括了硝化作用(Nitrification)、反硝化作用(Denitrification)、厌氧氨氧化(Anmmox)、铁氨氧化(Feammox)以及硝酸盐异化还原成铵(DNRA)等过程。稻田土壤砷转化与氮循环过程联系紧密。
稻田淹水条件下,砷氧化微生物可以硝酸盐为电子受体氧化As(Ⅲ),从而降低砷移动性(Lin et al.,2018;Li et al.,2019;Wu et al.,2021)。进一步研究发现,硝酸盐还原耦合砷氧化过程主要由具有完整反硝化基因的砷氧化微生物及硝酸盐还原微生物驱动(Li et al.,2019)。此外,砷污染稻田土壤施加硝态氮肥可抑制土壤中铁氧化物的还原溶解,且促进亚铁氧化过程(刘同旭等,2019),从而有利于砷的吸附固定(Chen et al.,2008)。除硝态氮肥外,施加铵态氮肥对砷的移动性亦具有显著影响(Liu et al.,2022)。最新的研究发现,厌氧氨氧化可与砷还原过程耦合,该过程的发现为土壤氮、砷元素生物地球化学行为研究提供了新认识(Zhang et al.,2022)。近年来,厌氧氨氧化与Fe(Ⅲ)还原耦合的铁氨氧化过程对稻田砷地球化学行为的影响也受到广泛关注(Li et al.,2019;Liu et al.,2022)。砷污染稻田土壤淹水条件下,施加铵态氮肥可促进含砷的氧化铁矿物的还原溶解并释放砷,从而导致砷移动性与有效性增强(Chen et al.,2023)。此外,研究发现,在地下水环境条件下,硝酸盐异化还原成铵可与砷还原过程耦合(Gao et al.,2021;Jiang et al.,2022),但该耦合过程在土壤环境的研究报道目前还较少。
本文基于已有的文献报道,对稻田土壤微生物氮循环过程、砷的微生物转化过程及不同氮循环过程对砷迁移转化的影响等进行系统分析和总结,并提出了稻田土壤氮循环与砷形态转化耦合研究的发展趋势。
1 稻田土壤微生物氮循环
氮循环是由微生物驱动的各种氮的氧化还原过程,如图1所示,包括硝化作用、反硝化作用、厌氧氨氧化、铁氨氧化和硝酸盐异化还原成铵等。施加铵态氮肥可促进稻田土壤中铵态氮氧化,如厌氧氨氧化过程、异化Fe(Ⅲ)还原耦合的铁氨氧化过程等;施加硝态氮肥,稻田土壤中则易发生硝酸盐异化还原成铵、反硝化过程等。
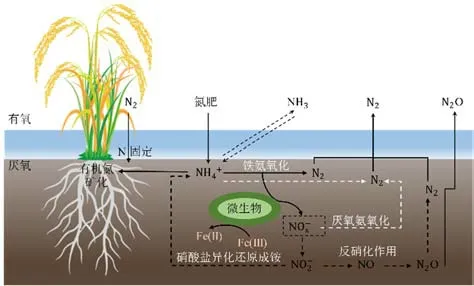
图1 稻田土壤氮循环过程(Yi et al.,2019)Figure 1 Nitrogen cycling process in paddy soil
1.1 硝化作用
硝化作用由氨氧化(Ammonia oxidation)与亚硝酸盐氧化(Nitrite oxidation)两步组成,且硝化作用连接了固氮作用与反硝化作用,所以该过程在土壤-植物系统氮循环中发挥关键作用(Li et al.,2018)。硝化作用分为自养型和异养型。微生物将NH4+/NH3氧化成NO3-或NO2-,并从中获取能量的过程是自养型硝化,而有些微生物不在该过程中获取能量则为异养型硝化(Hayatsu et al.,2008)。稻田土壤中主要由自养型硝化微生物驱动硝化过程(De Boer et al.,2001)。硝化过程的关键微生物有参与氨氧化过程的氨氧化细菌(Ammonia-oxidizing bacteria,AOB)、氨氧化古菌(Ammonia-oxidizing archaea,AOA)和全程氨氧化细菌(Comammox)(De Boer et al.,2001;Li et al.,2018;Dai et al.,2021),以及参与亚硝酸盐氧化的亚硝酸盐氧化菌(NOB)(Hayatsu et al.,2008;Hayatsu et al.,2021)。不同生态系统中,氨氧化菌的类型不同。比如在海洋中,AOA是优势氨氧化菌;而在农田土壤中,AOB氨氧化作用更强(Francis et al.,2007;Jia et al.,2009;Morimoto et al.,2011)。因此,AOB在稻田土壤中发挥的氨氧化作用强于AOA。比如,Li et al.(2006)研究发现,淹水水稻土中AOB的丰度与硝化活性成显著正相关;Hussain et al.(2011)研究水稻根际土壤氨氧化菌与反硝化菌丰度变化时发现,淹水条件下AOB与反硝化菌丰度增加,而AOA无明显变化;近期的研究发现,使用有机肥替代矿质肥料可以提高稻田土壤硝化速率,且AOB对硝化作用的贡献大于AOA(Dai et al.,2021)。
亚硝酸盐氧化过程主要由亚硝酸盐氧化菌(NOB)参与,分布在Nitrobacter、Nitrospina、Nitrococcus和Nitrospira等4个属(Hayatsu et al.,2008)。2009年,Ishii et al.(2009,2011)基于大规模16S rRNA基因测序研究了水稻中亚硝酸盐氧化菌群落,发现水稻土中Nitrospira属微生物具有亚硝酸盐氧化能力。Nahar等报道了孟加拉国水稻土中NOB的相对丰度与硝化速率成正相关(Nahar et al.,2020),证明NOB参与了硝化反应。
1.2 反硝化作用
微生物驱动的反硝化作用是一个呼吸过程,主要发生在厌氧或者缺氧环境中。在该过程中,利用无机碳化合物作为碳源的自养型反硝化微生物(Ma et al.,2020)与利用有机碳源做电子供体的异养反硝化微生物将硝酸盐(NO3-)逐步还原成亚硝酸盐(NO2-)、一氧化氮(NO)、一氧化二氮(N2O)和氮气(N2)(Ren et al.,2022)。其中,硝酸盐还原酶(Nar)、亚硝酸盐还原酶(Nir)、一氧化氮还原酶(Nor)和一氧化二氮还原酶(Nos)发挥了重要作用(Jones et al.,2008;Miralles Robledillo et al.,2021)。研究发现,稻田土壤中反硝化微生物种类多样(Ishii et al.,2011)。比如,Saito et al.(2008)利用稳定同位素探针(SIP)技术分析了稻田土壤反硝化微生物,鉴定出两株以琥珀酸为电子供体的反硝化微生物即Burkholderiales和Rhodocyclales菌。Ishii et al.(2009)通过克隆文库分析,在稻田土壤中发现反硝化微生物在目水平上属于Burkholderiales、Rhodocyclales和Desulfomonadales。在Burkholderiales目中,Herbaspirillumspp.被确定为进行硝酸盐呼吸和反硝化的关键参与者。在水稻土中分离出属水平上的反硝化微生物Bradyrhizobium、Dechloromonas、Herbaspirillum和Pseudogulbenkiania(Tago et al.,2011)。近期基于宏基因组学研究发现,在中国沿海潮滩、盐碱地和内陆水稻土中广泛存在Alphaproteobacteria-affiliating nosZ clade I和Bacteroidetes-affiliating nosZ clade II反硝化微生物(Han et al.,2021)。其中clade I和clade II是Nos编码基因nosZ的两个不同遗传支系(Domeignoz-Horta et al.,2018)。clade I发现已久,存在于多种细菌和古菌当中。clade II是一个新发现的nosZ基因遗传支系,其在基因表达、电子传递和N2O还原活性方面比clade I具有更加明显的特征(Duan et al.,2018)。
1.3 厌氧氨氧化
反硝化作用一直被认为是产生N2的唯一途径,直到厌氧氨氧化的发现。厌氧氨氧化过程是厌氧或缺氧条件下,铵(NH4+)氧化与亚硝酸盐(NO2-)还原耦合产生N2(Mulder et al.,1995;Zhu et al.,2011;Shan et al.,2018;Shen et al.,2020)。迄今为止,已报道了5个厌氧氨氧化菌属Candidatus Brocadia、Kuenenia、Anammoxoglobus、Jettenia和Scalindua,而在水稻土中占优势的是Candidatus Brocadia属(Han et al.,2021)。
厌氧氨氧化过程在稻田土壤中很活跃,也被认为是稻田生态系统中氮损失的重要途径(Yang et al.,2015)。由于稻田环境复杂,厌氧氨氧化过程在稻田土壤中也受多种因素影响。例如,不同形态氮肥的施加会影响厌氧氨氧化细菌丰度(Shen et al.,2020)。Shen et al.(2020)在水稻土里分别添加了化肥与有机肥,研究厌氧氨氧化菌的活性与丰度。同位素示踪实验结果表明,厌氧氨氧化活性及厌氧氨氧化细菌丰度在有机肥处理中最高,且发现淹水稻田土壤中厌氧氨氧化过程可产生大量N2。除氮肥形态的影响,厌氧氨氧化活性还会受到稻田土壤pH和有机碳等影响(Shan et al.,2018),这对稻田氮损失研究具有重要指导意义。
1.4 铁氨氧化
稻田土壤富含丰富的Fe与NH4+,周期性淹水与落干使得Fe在水稻土中发生各种转化,厌氧氨氧化与铁还原的耦合被称为“Feammox”反应(Li et al.,2019)。铁氨氧化过程是在厌氧条件下,微生物将NH4+作为电子供体,氧化Fe(Ⅲ),最终产物有N2、NO3-或NO2-(程宽等,2022)。铁与氮循环的耦联已经被多次报道。2005年,Clément et al.(2005)研究河岸森林沉积物中Fe和氮循环的动力学发现,存在将Fe(Ⅲ)作为电子受体同时氧化NH4+的生物过程,且该过程在热力学上是可行的,并且指出该过程可能在土壤氮循环中发挥关键作用。2006年,“Feammox”过程被首次提出(Sawayama,2006)。Sawayama(2006)在厌氧条件下探讨了NH4+氧化与Fe(Ⅲ)-EDTA-Na还原耦合过程,该实验在玻璃反应器中厌氧运行数天,在特定时间向反应器中加入Fe(Ⅲ)-EDTA-Na时,反应器中铵根浓度不断降低,同时亚硝酸盐浓度上升;当反应器中不供给Fe(Ⅲ)-EDTA-Na时,反应器中铵根浓度升高,且亚硝酸盐浓度降低。该实验结果证明铁还原微生物可将Fe(Ⅲ)还原为Fe(Ⅱ)同时将NH4+氧化成NO2-。2012年,Yang et al.(2012)发现Feammox反应在热带旱地土壤中可产生N2、NO3-或者NO2-,并认为生成N2是Feammox反应中主要的氮损失途径。
稻田环境中Feammox反应是氮损失的主要途径之一。2014年,Ding et al.(2014)首次在水稻土中发现Feammox反应,且证明生成N2是稻田Feammox反应的主要途径。据估算,稻田土壤中约3.9%-31%的氮肥是通过Feammox反应损失。2020年,Ding et al.(2020)报道稻田土壤中与Feammox相关的微生物包括Geobacter、Candidatus Solibacter、Nitrososphaeraceae、Leptolyngbya、Pseudomonas、Anaeromyxobacter等。Ding et al.(2021)进一步采用15N标记的同位素示踪技术,研究了Feammox反应在稻田土壤垂直分布的差异及与传统氮损失途径的关系。结果表明,Feammox导致的氮损失在20-40 cm的水稻土中高达26.1%。2015年,Huang et al.(2015)首次报道了Acidimicrobiaceaesp.A6菌在Feammox过程起关键作用。2018年,Acidimicrobiaceaesp.A6菌首次在美国新泽西州的河岸湿地土壤中被分离出来,该菌株在厌氧条件下能氧化NH4+同时还原Fe(Ⅲ)(Senko et al.,2018)。2019年,Shuai et al.(2019)采集美国新泽西州的河岸湿地沉积物,建造了人工湿地对Acidimicrobiaceaesp.A6菌进行富集培养。接种A6菌,高铁含量的人工湿地中NH4+去除率为25.0%±7.3%,而低铁含量的NH4+去除率仅11.0%±9.7%,这表明可以通过促进Feammox反应提高湿地NH4+的去除。Zhang et al.(2021)等采集了不同类型的农田土壤,研究了土壤中自养氨氧化微生物多样性、丰度及影响这些微生物对土壤氨氧化的相对贡献。研究发现,Feammox反应中,Acidimicrobiaceaesp.A6具有最高的氨氧化速率,达到了 (1.23±0.98) μmol·g-1·d-1。综上所述,目前仅分离出一株直接参与Feammox过程的单菌Acidimicrobiaceaesp.A6,参与Feammox反应的微生物仍有待进一步挖掘。
1.5 硝酸盐异化还原成铵
硝酸盐异化还原成铵是微生物氮循环的重要过程之一。与反硝化作用不同,DNRA是在厌氧条件下,微生物将NO2-/NO3-作为电子受体生成NH4+的过程,由nrfA基因编码的硝酸盐还原酶Nrf催化(Pandey et al.,2018;Zhao et al.,2022)。研究发现,在碳源充足而氮源缺乏的环境条件下,更容易发生DNRA而不是异养反硝化作用(Pandey et al.,2020;Broman et al.,2021)。驱动DNRA的微生物是发酵型细菌而参与反硝化过程的菌是呼吸细菌。反硝化可造成环境的氮损失而DNRA可保留氮素,对喜铵的农作物更有利(周利等,2020)。
纯培养下的DNRA现象早在1938年已被发现,随着研究的不断深入,驱动该过程的其他功能微生物也逐步被发现(Woods,1938;Pandey et al.,2020)。由于水稻是喜铵作物,而DNRA过程能为水稻提供铵,因此DNRA在稻田环境中具有重要作用(Nojiri et al.,2020)。2020年,Nojiri et al.(2020)研究发现,参与DNRA的相关功能微生物主要是Anaeromyxobacter和Geobacter,并发现驱动DNRA和反硝化作用的微生物有所不同,但二者的反应速率相近。稻田土壤中,DNRA可以有效得降低因反硝化及硝酸盐淋失造成的氮损失,有利于稻田氮损失的控制与农作物的低施肥管理,同时缓解环境氮素污染(Nojiri et al.,2020)。
2 稻田土壤砷的微生物转化
砷(As)是一种有毒且致癌的类金属物质,对人体健康可产生极大的威胁(Hussain et al.,2021;Rokonuzzaman et al.,2022)。世界上很多地区都存在砷污染的困扰。例如,孟加拉国地下水砷污染严重,灌溉含砷污染的地下水导致水稻与土壤中富集大量的砷,人体摄入砷超标大米可造成严重的健康问题(Islam et al.,2017)。此外,长期暴露在砷污染的环境中可导致人体的神经系统、呼吸系统、皮肤等病变(Shahid et al.,2018;Hussain et al.,2019)。水稻-土壤系统中淹水-落干交替条件下,砷的生物地球化学行为发生周期性的变化(Hussain et al.,2021)。如图2所示,当稻田处于淹水条件时,铁氧化物还原溶解并释放As至孔隙水中,且砷以亚砷酸盐(As(Ⅲ))的形式存在(Herath et al.,2016;Beiyuan et al.,2017;Lin et al.,2018);当稻田处于落干环境时,砷主要以砷酸盐(As(Ⅴ))的形式存在且生物利用度降低(Dixit et al.,2003;Oremland et al.,2005)。水稻-土壤系统中砷的形态受各种因素的影响,例如pH、Eh、天然有机物(DOM)、铁(锰)氧化物和微生物等,特别是稻田土壤微生物对砷的迁移与生物转化有着重要的影响(Upadhyay et al.,2018)。
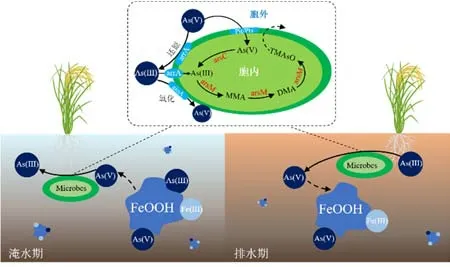
图2 稻田土壤砷微生物转化过程Figure 2 Microbial transformation process of arsenic in paddy soil
2.1 砷还原
微生物介导的As(Ⅴ)还原有两种机制,一种是微生物介导细胞质As(Ⅴ)还原的解毒机制,另一种是微生物介导异化As(Ⅴ)还原的呼吸机制。微生物对砷解毒机制主要通过自带的砷抗性编码酶和外排泵将细胞质中的砷排出,这种机制主要存在于革兰氏阴性和革兰氏阳性菌中(Mateos et al.,2017)。解毒过程主要通过磷酸盐转运蛋白以砷酸盐的形式吸收As(Ⅴ),再通过砷酸盐还原酶将As(Ⅴ)还原成As(Ⅲ)排出(Rosen,2002;Zhang et al.,2015;Qiao et al.,2018)。该过程与arsC基因相关,且普遍存在于好养、厌氧微生物中(钟松雄等,2018)。呼吸作用通过砷酸盐呼吸还原酶(arrA)进行,厌氧条件下将As(Ⅴ)作为微生物呼吸的末端电子,并通过呼吸还原作用为自身提供能量(Zhang et al.,2015;Kumarathilaka et al.,2018)。例如,很早被分离出来的异化砷酸盐还原菌Bacillussp.SF-1具有强烈的砷酸盐还原能力(Soda et al.,2006)。
微生物介导的异化As(Ⅴ)还原可直接影响砷在稻田土壤中的移动性,并影响As与碳(C)、氮(N)、铁(Fe)、硫(S)等元素循环的耦合过程(Zhu et al.,2017)。淹水稻田中有效态砷浓度升高,一方面由于异化砷还原微生物将吸附在氧化铁矿物上的As(Ⅴ)还原成As(Ⅲ)导致土壤溶液中的砷浓度升高;另一方面与异化铁还原菌(DIRB)驱动的铁还原过程密切相关。异化铁还原菌可将Fe(Ⅲ)氧化物还原成Fe(Ⅱ),从而导致吸附在铁氧化物上的As(Ⅴ)释放到孔隙水中(Qiao et al.,2018)。例如,Wang et al.(2016)利用ShewanellaoneidensisMR-1和ShewanellaoneidensisMR-4两株典型的DIRB菌,研究它们对砷还原释放的影响,研究发现在这两种菌存在条件下,砷可快速的从氧化铁矿物上释放。厌氧条件下将乳酸加入富含铁还原微生物的沉积物中,在第1天内As(Ⅴ)开始还原且在第2天就到达高峰。因此,厌氧环境下(如淹水稻田),吸附在铁氧化物上的As(Ⅴ)可以被异化铁还原过程释放(Ko et al.,2019)。
除了上述异化砷还原微生物,水稻土中已有很多与arsC基因相关的砷还原微生物被鉴定出。2015年,Zhang et al.(2015)在全国范围内采集了13个砷污染的水稻土,利用T-RFLP和克隆文库方法研究水稻土中参与砷转化的微生物,研究发现细胞质砷还原微生物主要属于变形菌门包括Hoeflea、Sinorhizobium、Mesorhizobium、Polymorphum、Enterobacter等微生物,而arrA基因相关的微生物主要为Geobacteruraniireducens,且arrA基因丰度明显低于arsC基因。这可能是因为arrA基因只存在厌氧微生物中,而arsC基因则可以存在厌氧或好氧微生物中。
2.2 砷氧化
环境中微生物砷氧化是影响砷迁移转化的重要因素。通常认为As(Ⅲ)比As(Ⅴ)移动性更强,毒性更大(Zhang et al.,2022)。微生物氧化As(Ⅲ)由AioA和AioB亚基组成的As(Ⅲ)氧化酶催化(Zhang et al.,2017;Li et al.,2019)。除此之外,学者还发现亚砷酸盐氧化酶ArxA能催化As(Ⅲ)氧化与硝酸盐还原耦合 (Zhang et al.,2015)。驱动砷氧化的微生物包括化能自养型和异养型。化能自养型砷氧化微生物能利用氧气(有氧条件下)或硝酸盐(厌氧条件下)等作为电子受体将As(Ⅲ)氧化并为自身提供能量(Slyemi et al.,2012)。异养型砷氧化被认为是一种细胞解毒的机制,不能为自身供能(Muller et al.,2003)。
在稻田土壤中,砷氧化微生物被广泛研究。在非淹水条件下,空气进入土壤形成有氧条件,此时Fe(Ⅱ)与As(Ⅲ)会被生物或非生物氧化;在淹水条件下,As(Ⅲ)会在特定条件下被微生物氧化,添加硝酸盐可激活一些砷氧化微生物氧化Fe(Ⅱ)或As(Ⅲ)(Lin et al.,2018)。2015年,Zhang et al.(2015)在砷污染水稻土中分离出一株自养型砷氧化菌Paracoccussp.SY,该菌株在厌氧条件下,利用硝酸盐作为电子受体,将As(Ⅲ)氧化成As(Ⅴ),且在好氧与厌氧条件下都具有氧化亚砷酸盐功能。该研究对降低稻田砷的移动性及水稻生长过程中砷积累具有重要意义。Lin et al.(2018)在淹水条件下向砷污染稻田土壤中加入不同浓度硝酸盐,硝酸盐的添加促进了孔隙水中As(Ⅲ)氧化,As(Ⅴ)浓度也随之升高,且高浓度硝酸盐处理组As(Ⅲ)氧化酶基因aioA的丰度增加了1.6-4.2倍。除上述淹水水稻土中的砷氧化微生物,许多根际微生物例如Rhizobiales和Burkholderials等均能在好氧和厌氧条件下驱动土壤中的砷氧化(Zhang et al.,2015)。
2.3 砷甲基化
稻田土壤砷的形态转化除As(Ⅴ)还原和As(Ⅲ)氧化外,还包括无机砷转化为有机砷。有机砷主要包括一甲基砷(MMA)、二甲基砷(DMA)和三甲基胂氧化物(TMAO)(Zhu et al.,2017)。砷甲基化被认为是一种解毒过程,土壤微生物砷甲基化由亚砷酸盐甲基转移酶(arsM)催化(Zhao et al.,2013)。在淹水稻田中砷甲基化很容易发生,并容易产生气态砷挥发到大气中(Mestrot et al.,2011)。
砷甲基化基因(arsM)广泛分布在稻田土壤微生物中,并与甲基砷浓度成正相关(Wang et al.,2018)。2020年,Hashmi et al.(2020)采集了15个水稻土样品研究土壤中砷形态及浓度水平、砷代谢基因及其与土壤理化性质的相关性。研究发现,15个水稻土中不同砷形态及arsM基因丰度具有显著差异,且总砷浓度与arsM基因丰度呈显著负相关。水稻土中参与砷甲基化的微生物种类多样,比如Proteobacteria、Gemmatimonadales和Firmicutes等水稻根际微生物(Zhang et al.,2015)以及Desulfovibrio、Terrisporobacter和Clostridium等厌氧内生菌(Chen et al.,2022)。2015年,Wang et al.(2015)从水稻土中分离出一株硫酸盐还原菌Clostridiumsp.BXM,该菌株普遍存在于厌氧水稻土中,具有砷甲基化能力。2016年,Huang et al.(2016)在水稻土中分离出一株Cytophagaceae科的菌株SM-1,其在稻田土壤中含量丰富,具有砷甲基化和砷挥发能力,接种该菌株可显著促进土壤中砷甲基化及砷挥发。SM-1菌株的发现对于砷污染土壤生物修复具有重要意义。
稻田土壤砷甲基化过程受土壤理化性质等多种因素影响,如pH、DOC等。2013年,Zhao et al.(2013)在6个不同采样点采集6个不同的水稻土,研究稻田土壤淹水条件下,砷甲基化过程的影响因子。研究发现arsM基因丰度与土壤pH正相关,土壤孔隙水中甲基砷与pH和arsM基因丰度负相关,与溶解性有机碳正相关。因此,相比arsM基因丰度,可显著影响砷甲基化微生物活性的土壤条件比如pH、溶解性有机碳等对土壤砷甲基化过程影响更大。已有研究表明,水稻土中硫酸盐还原菌及 产甲烷古菌分别参与了砷甲基化及脱甲基化过程,并共同控制土壤中甲基砷的动态变化(Chen et al.,2019)。
2.4 脱甲基化
稻田土壤微生物介导砷甲基化以生成一甲基砷(MMA)和二甲基砷(DMA)为主,土壤中部分微生物比如产甲烷古菌也能将甲基砷进行脱甲基化(Chen et al.,2019)。2011年,Yoshinaga et al.(2011)采集MAs(Ⅴ)污染的高尔夫球场土壤研究了MAs(Ⅴ)脱甲基化,研究发现MAs(Ⅴ)脱甲基化过程首先由Burkhoderia菌将MAs(Ⅴ)还原成MAs(Ⅲ),然后由Streptomyces将MAs(Ⅲ)脱甲基化成As(Ⅲ),这是首次报道甲基砷的脱甲基过程。
淹水稻田中,MMA与DMA也可被脱甲基。向水稻土中外源加入二甲基砷并在厌氧条件下淹水培养,研究砷甲基化与脱甲基化的主要功能微生物。随着甲烷的生成,孔隙水中DMAs迅速降低,研究发现硫酸盐还原菌与产甲烷菌分别参与了砷的甲基化与脱甲基过程(Chen et al.,2019)。Chen et al.(2021)从土壤中分离出一株厌氧菌Bacillussp.CZDM1。该菌基因组中含有C-As裂解酶基因arsI,在厌氧硝酸盐还原条件下具有MMAs(Ⅲ)脱甲基化功能。据研究,C-As裂解酶基因BcarsI在反硝化细菌中普遍存在,可实现MMAs(Ⅲ)的脱甲基化与反硝化过程的耦合,且该耦合过程在稻田土壤十分常见。除此之外,一些好氧内生菌也能将MMAs(Ⅲ)脱甲基化(Chen et al.,2022),可见,在复杂的稻田土壤环境中,砷脱甲基化的过程并不依赖于某个单一的物种,而是存在多种微生物共同参与。
3 稻田土壤氮循环过程对砷形态转化的影响
在土壤环境中,铁(Fe)、碳(C)、氮(N)循环对砷的形态转化均具有重要影响。例如,淹水稻田中微生物驱动的硝酸盐还原耦合As(Ⅲ)的氧化可固定砷(Lin et al.,2018);NH4+与Fe(Ⅲ)共存的稻田土壤中可发生Feammox反应,从而提高砷的移动性(Ding et al.,2014;Zhang et al.,2021)。如图3所示,稻田氮循环耦合砷形态的转化在砷的生物地球化学循环中发挥着重要作用,解析氮、砷耦合机制可为土壤砷污染控制提供科学依据。
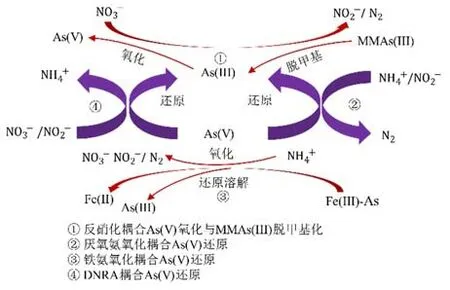
图3 稻田土壤中氮砷耦合过程Figure 3 The coupling process of nitrogen and arsenic in paddy soil
3.1 硝化过程
稻田土壤具有干湿交替的周期性变化特征,其中氮循环过程与砷形态转化存相互耦合。氮循环通过参与铁氧化物和溶解Fe(Ⅱ)之间的动态转化,可以影响土壤中As的富集(Smith et al.,2017)。在有氧条件下,稻田土壤硝化作用产生NO3-(Devkota et al.,2013;Verhoeven et al.,2018),NO3-可抑制土壤中氧化铁矿物的还原溶解,从而减缓砷的释放(Liu et al.,2019)。Weng et al.(2017)采集了砷污染的浅层地下水,利用稳定同位素技术发现含氧地下水中砷的迁移转化与氮循环中的硝化作用密切相关。在含氧的近端水层中硝化作用提高了NO3-浓度,高浓度的NO3-降低了含氧地下水中砷的移动性。因此,稻田土壤中硝化作用有利于砷吸附在氧化铁矿物上,从而降低砷的移动性(Zhu et al.,2017;Wang et al.,2019)。此外,不同酸、碱性及好、厌氧条件的稻田土壤中,硝化微生物活性与丰度具有显著差异。弱碱性稻田土壤中AOA活性明显优于AOB,而AOB在中性和酸性土壤中活性更高(Zhang et al.,2019)。综合以上研究可知,一方面,在砷污染的环境下硝化作用会直接与砷耦合影响砷的迁移转化。另一方面,砷污染稻田土壤淹水和落干的周期性变化造成土壤pH、Eh等理化性质的变化,会影响土壤硝化微生物驱动的硝化反应过程,从而间接影响吸附在氧化铁矿物上砷的还原释放。也会对硝化作用和砷形态转化的耦合产生间接影响。
3.2 反硝化过程
已有研究表明,厌氧环境中亚砷酸盐氧化与硝酸盐还原过程耦合可降低砷的移动性(Zhang et al.,2015;Zhang et al.,2017)。Senn et al.(2002)报道硝酸盐可以在厌氧条件下通过微生物可以对As(Ⅲ)进行氧化。硝酸盐耗尽情况下,一般有As(Ⅲ)存在;但硝酸盐丰富且厌氧情况下,As(Ⅴ)则为主要产物。Oremland et al.(2002)研究发现,一些依赖硝酸盐的氧化细菌(如MLHE-1等)能将As(Ⅲ)氧化与部分反硝化相耦合。而Rhine团队(Rhine et al.,2006)则分离了两种自养As(Ⅲ)氧化菌株DAO1和DAO10,发现其能够将As(Ⅲ)氧化与完全反硝化相耦合的同时来固定二氧化碳。Zhang et al.(2015)在砷污染水稻土中分离出一株化学自养型砷氧化菌SY,该菌株可以在好/厌氧条件下分别以氧气或硝酸盐为电子受体氧化三价砷,并从中获取能量。在淹水土壤中接种该菌株,可显著促进三价砷的氧化过程。Li et al.(2019)采集未污染的水稻土,在厌氧条件下添加As(Ⅲ)及NO3-进行微宇宙实验。研究发现,添加了NO3-后,As(Ⅲ)氧化速率显著升高,宏基因组分析发现硝酸盐还原菌主要为Pseudogulbenkiania,砷氧化菌主要为Azoarcus spp.。该研究表明硝酸盐还原菌和砷氧化菌共同驱动硝酸盐还原耦合砷氧化过程。
此外,最近的研究发现,反硝化作用也可耦合MAs(Ⅲ)脱甲基化。Chen et al.(2021)分离出反硝化细菌Bacillussp.CZDM1,能够在厌氧条件下将MAs(Ⅲ)脱甲基化。因此,反硝化作用在砷污染土壤中对砷的迁移转化也具有重要影响,对以上耦合过程的主要影响因子还有待进一步研究。除以上微生物因素,在砷污染稻田中不同土壤理化条件也可影响反硝化速率,从而影响砷的固定释放。Liang et al.(2021)研究发现,反硝化微生物活性受气候因子、土壤pH和nirS基因生物多样性影响,且反硝化基因nirS和nirK丰度与土壤碳/氮比成显著负相关。综上所述,砷污染的稻田中,反硝化作用不仅可直接耦合砷转化,还可以通过土壤的碳/氮比等理化条件影响反硝化微生物的丰度和活性,从而间接将反硝化作用与砷进行耦合。
3.3 厌氧氨氧化过程
最近的研究发现厌氧氨氧化可以与As(Ⅴ)还原过程耦合。Zhang et al.(2022)采集砷污染的稻田土壤,外源加入15N-尿素及As(Ⅴ)厌氧培养,研究发现同时加入15N-尿素和As(Ⅴ)可显著提高砷还原基因arrA及厌氧氨氧化基因hzsB的丰度。稳定同位素示踪技术和高通量测序技术进一步揭示水稻土中驱动厌氧氨氧化与砷还原耦合的关键微生物主要包括Halomonas,Pelagibacterium和Chelativorans。研究还发现,水稻土中还同时存在传统厌氧氨氧化过程,Ca.Brocadia是水稻土中优势的厌氧氨氧化菌。该研究拓展了稻田土壤中氮砷循环耦合过程的认识。此外,与之前描述的过程类似,土壤pH、有机碳等土壤理化性质均可影响厌氧氨氧化活性(Shan et al.,2018)。其厌氧氨氧化主要发生在NO3-或NO2-与NH4+共存的还原环境中,在高浓度的NO3-或NO2-条件下,Fe(Ⅱ)易被氧化为铁氧化物,从而吸附固定砷,降低砷的移动性。目前在稻田砷污染环境中,厌氧氨氧化耦合砷转化过程的研究报道较少,需要进一步关注该过程。
3.4 铁氨氧化过程
稻田土壤通常施加铵态氮肥作为农用氮肥。厌氧条件下,Feammox过程可促进氧化铁还原溶解从而导致吸附态砷释放(Xue et al.,2020;Chen et al.,2023),因此,Feammox过程在砷污染的环境中能够提高砷移动性。Weng et al.(2017)基于氮、氧、氢同位素及氮、铁、砷形态与价态分析,研究了台湾浊水溪冲积扇砷污染地下水中氮素转化过程及其对砷移动性的影响。研究发现,Feammox反应促进铁还原释放砷可能是造成扇央区地下水中高浓度砷的主要原因。Chen et al.(2023)基于大田试验研究了施加不同形态氮肥对稻田土壤中砷移动性与有效性的影响。研究发现施加铵态氮降低土壤Eh,有利于促进含砷的氧化铁矿物还原溶解释放砷。厌氧条件下,氨氧化可耦合砷还原过程,促进As(Ⅴ)被还原为移动性较强的As(Ⅲ);此外,厌氧氨氧化可耦合铁还原过程,促进砷从氧化铁矿物上被释放。
Huang et al.(2016)研究发现,土壤pH、Fe(Ⅲ)及NH4+是影响Feammox反应活性的关键环境因子且Feammox反应更易发生在低溶解氧或者缺氧条件下。最近的研究发现秸秆和铵态氮共施条件下有利于砷的固定(Liu et al.,2022)。其原因主要有两方面:一是外源加入秸秆可提高土壤中溶解性有机质(DOM)含量,DOM的增加使得Feammox反应微生物更易以DOM而不是Fe(Ⅲ)为电子受体。因此,DOM的增加抑制了Feammox反应,从而不利于吸附态砷的还原释放;二是施加的铵态氮水解降低土壤pH,增加了铁氧化物表面的正电荷从而更易于吸附固定砷。由此可见,Feammox反应受多种环境因子的影响,可以通过调控Feammox反应过程来影响砷的移动性与有效性。
3.5 硝酸盐异化还原成铵
厌氧条件下,DNRA过程普遍存在于水稻土中,特别在硝酸盐浓度较低、DOC浓度高的稻田土壤中更容易发生DNRA过程(Robertson et al.,2016),该过程提升了稻田肥力,有利于水稻植株的生长。但另一方面,在这样的环境中能够促进砷从铁氧化物上解吸导致孔隙水中游离态的砷浓度升高。学者(Jiang et al.,2022)从河套平原高砷地下水中分离出一株异化硝酸盐还原菌Citrobactersp.G-C1,该菌可在DNRA过程中同时还原As(Ⅴ),且该菌也具有有机砷脱甲基化功能。以上结果表明,DNRA可与砷还原过程耦合从而提高砷的移动性。
已有研究表明DNRA与(Cheng et al.,2022)稻田土壤中砷相耦合的同时,也受土壤pH、有机碳及总氮等理化性质影响(Liu et al.,2022;Yang et al.,2022),但目前关于DNRA与砷转化的耦合过程研究还比较少,有待进一步研究探讨。
4 结论与展望
4.1 结论
(1)水稻生产施用大量的尿素或铵态氮,稻田氮素循环为土壤生态系统元素循环的核心过程之一。砷污染的稻田土壤中铵态氮与砷共存是普遍的现象。氮循环可以显著影响土壤中砷的形态转化过程,从而影响其移动性与生物有效性。
(2)稻田土壤中氮的循环过程可以与砷形态转化的多个过程耦合从而对砷的移动性造成不同影响。其中,硝化和反硝化作用有利于砷的吸附固定;而厌氧氨氧化、铁氨氧化及硝酸盐异化还原成铵可促进砷的还原释放。此外,反硝化作用也可以耦合砷脱甲基化过程,从而提高砷毒性。
4.2 展望
综上所述,氮循环与水稻土砷的形态转化密切相关。在砷污染的稻田土壤中,氮循环耦合砷转化的过程也逐步被发现,并得到深入研究。基于本文的综述,笔者认为以下3个方面可作为今后研究的重点:
(1)稻田土壤具有干湿交替特征,在不同的水分条件下,土壤中pH、Eh等差异明显,且不同的土壤深度氧气含量也有差异。如何区分不同的水分条件和不同土壤深度下硝化、反硝化、厌氧氨氧化、铁氨氧化,以及硝酸盐异化还原成铵等氮循环过程;不同氧化还原条件下,驱动氮循环耦合砷转化的关键功能微生物和影响这些耦合过程的关键环境因子,以及这些耦合过程对砷移动性的贡献等,是需要重点关注的科学问题。
(2)铁氧化物在稻田土壤中含量丰富,铁循环在砷的迁移转化过程中发挥着重要作用。Feammox反应可促进铁氧化物的还原溶解并提高砷的移动性,但目前驱动该过程的功能微生物仍有待进一步挖掘。Feammox反应在砷污染稻田土壤中对砷移动性的贡献有多大也仍然不清楚。
(3)已知有机砷毒性小于无机砷,促进砷甲基化过程一定程度上可以降低稻米中无机砷的积累,从而降低人们食用稻米的风险。氮循环耦合砷甲基化或脱甲基化的研究还处于起步阶段,特别是如何调控氮循环耦合砷转化过程以实现降低稻米中总砷含量及无机砷的比例,有待进一步探讨。
(4)虽然目前已发现氮循环的多个过程可以与砷转化耦合,但其耦合机理仍有待进一步研究。特别是影响这些耦合过程的关键环境因子不明确,这些耦合过程对稻田土壤砷移动性的贡献程度不清晰,以及如何通过对这些耦合过程的定向调控实现对砷的固定,有待进一步研究。
猜你喜欢
中老年保健(2022年3期)2022-11-21
国际太空(2021年11期)2022-01-19
环境保护与循环经济(2017年7期)2018-01-22
兽医导刊(2016年12期)2016-05-17
中国环境科学(2016年3期)2016-02-08
华东理工大学学报(自然科学版)(2015年3期)2015-11-07
中国果业信息(2015年12期)2015-01-24
应用海洋学学报(2014年2期)2014-11-26
河南科技(2014年24期)2014-02-27
电信工程技术与标准化(2014年3期)2014-02-27
