
果蝇Neuroligin 4 以非依赖于Neurexin 方式调控突触的发育
2023-10-07 01:49张新旺耿俊华李莉张娟彭泽旭
中国实验动物学报 2023年7期
张新旺,耿俊华,李莉,张娟,彭泽旭
(1.山西医科大学医学细胞生物学与遗传学教研室,太原 030001;2.东南大学发育与疾病相关基因教育部重点实验室,南京 210096)
突触的形成、发育及可塑性调节是神经网络构建的关键环节,而突触黏附分子是这一过程的重要组织者与调节因子。迄今为止,已有多种黏附分子被报道在突触的形成、成熟及功能行使中发挥重要作用[1-3]。其中,神经连接蛋白(neuroligin,Nlg)作为一类重要的突触黏附分子,因其与自闭症的紧密联系而在近年来受到广泛关注[4-6]。
Nlg 是一类在进化上高度保守的细胞黏附分子家族,广泛存在于包括线虫、果蝇、啮齿类和人类等多个物种的神经元及其靶细胞膜上[7]。研究显示,Nlg 在神经组织中主要作为突触后分子,通过其胞外端的乙酰胆碱酯酶类似结构域与突触前的配体分子——轴突蛋白(neurexin,Nrx)的胞外端相互作用形成跨突触的蛋白复合体,从而在突触的生长、成熟及功能行使中发挥作用[8-9]。但也有研究显示,Nlg 和Nrx 在突触前后均有定位[10-12],提示这两类黏附分子作用可能具有多样化的模式。
果蝇有4 种Nlg 分子(DrosophilaNlgs,DNlgs),在特征性结构域上与人的NLG1 具有较高的同源性[13]。在果蝇幼虫,DNlgs 广泛定位在脑和腹神经索的神经突触以及体壁肌肉的神经肌肉接头(neuromuscular junction,NMJ)处。在以果蝇幼虫NMJ 为突触模型的研究中发现,DNlg1~ 4 均具有调控突触发育的作用,表现为敲除单个DNlg 编码基因均会造成幼虫NMJ 形态异常与突触传导功能缺陷。其中,DNlg1~ 3 主要定位于NMJ 的肌肉细胞膜上,作为突触后分子起作用[13-15]。前期对DNlg4的研究显示,DNlg4 的缺失也会造成显著的NMJ 发育缺陷,主要表现为突触终扣(bouton)数量的显著减少。但不同于DNlg1~ 3,DNlg4 对NMJ 发育的调控是以突触前分子起作用[16]。考虑到已有研究表明DNlg1 对NMJ 的发育调控作用依赖于突触前的DNrx[13],那么位于NMJ 突触前的DNlg4,其对NMJ 发育的调控是否需要DNrx 的参与尚不清楚。
在本研究中,以果蝇幼虫NMJ 为突触模型,通过分析dnlg4 突变体、dnrx突变体以及dnlg4,dnrx双突变体等基因型果蝇的NMJ 发育缺陷,探究DNlg4 和DNrx 是否具有遗传上的相互作用,并利用免疫共沉淀技术检测二者是否具有在体的相互结合。结果发现,与已经报道的Nlg 作用方式不同,DNlg4 对神经肌肉突触发育的调控不依赖于突触前DNrx 的参与,这增加了人们对Nlg 分子调控突触发育多样性的认识。
1 材料与方法
1.1 材料
1.1.1 实验动物
本研究使用w1118果蝇作为野生型(wild type,WT)对照品系。dnlg4 缺失突变体、dnrx83缺失突变体、UAS-dnlg4 转基因果蝇以及OK6-Gal4 果蝇获自东南大学发育与疾病教育部重点实验室。dnlg4,dnrx双突变体果蝇由dnlg4 突变体与dnrx83突变体经遗传整合获得。所有果蝇品系均培养于25℃、相对湿度60%条件下,用标准玉米培养基饲养。
1.1.2 主要试剂与仪器
用于免疫荧光染色的一抗包括兔源DNlg4 抗体(C534;1 ∶100),鼠源DNlg3 抗体(1G8;1 ∶50),兔源DNrx 抗体(1 ∶100)(以上均来自东南大学);鼠源BRP 抗体(nc82;1 ∶50;DSHB),鼠源DLG 抗体(4F3;1 ∶50;DSHB);兔源HRP 抗体(1 ∶1000;Jackson Immuno Research,West Grove,PA);荧光二抗(Invitrogen,1 ∶500)。用于蛋白质印迹的一抗包括兔源DNlg4 抗体(C534;1 ∶1000),兔源DNrx 抗体(1 ∶1000),鼠源β-tubulin 抗体(DM1A;1 ∶10 000;Sigma)。HRP 偶联二抗(31430,31460;1 ∶10 000;Thermo Scientific)。
激光共聚焦显微镜(Zeiss LSM700,德国),体视显微镜(Motic SMZ-168,中国),三恒电泳仪(DYY-11B,北京六一,中国),高速冷冻离心机(Eppendorf centrifuge 5417R,德国)。
1.2 方法
1.2.1 待测样品的准备
(1)三龄幼虫头部样品的准备
将三龄幼虫置于盛有磷酸缓冲液(PBS)的平皿中,用镊子拖拽幼虫头部,使其与躯干分离。去除头部其他的成虫盘。用4%甲醛固定40 min 后备用。
(2)三龄幼虫NMJ 样品的准备
选取健康爬壁的三龄幼虫,用昆虫针固定其头尾,使其背部向上固定于琼脂糖平板上。用小剪沿背部气管中间线剪开背部体壁,除去幼虫内脏器官及成虫盘。将幼虫体壁展开用针固定,用PBS 冲洗1 次。加入4%多聚甲醛固定30~ 40 min 备用。
1.2.2 免疫荧光染色
固定后的样品用0.3% PBT(PBS+0.3%Triton X-100)清洗,用封闭液(1% BSA+PBT)在室温下封闭1 h,加入PBT 稀释的一抗于4℃孵育过夜。用PBT 清洗4 次后加荧光二抗,室温孵育2 h。PBT 清洗4 次后加入抗荧光淬灭剂(Vector)并封片。在激光共聚焦显微镜下进行观察拍照。
1.2.3 蛋白质免疫印迹(Western Blot)
分离20 只完整三龄幼虫头部,加入20 μL 2 ×SDS 上样缓冲液混匀,沸水煮5 min 后进行SDS 聚丙烯酰胺凝胶电泳。蛋白转至PVDF 膜后,将膜置于0.5‰ TBST 稀释的5%脱脂牛奶中封闭1 h。TBST 清洗后置于一抗稀释液(4% BSA,1% Tween-20,1 × PBS)中于4℃孵育过夜。将膜用TBST 清洗4 次后置于二抗稀释液中,室温孵育1 h 时。用ECL发光试剂盒(诺唯赞)进行显色。
1.2.4 免疫共沉淀
收集果蝇成虫,以液氮速冻后分离果蝇头部与躯干,收集果蝇头约1 g。加入裂解缓冲液(150 mmol/L NaCl,50 mmol/L Tris-HCl,0.5% NP-40,0.5 mmol/L EDTA,0.5% Triton X-100,1 mmol/L PMSF,1 × protease inhibitor(Roche),pH=8.0),用匀浆器充分破碎样品,放置冰上30 min 后,以12 000 r/min离心取上清液,并用缓冲液稀释2~ 5 倍。各取1 mL 组织上清液分别加入约3 μg DNlg4 多抗(c534),在 4℃孵 育 过 夜。每 管 加 入 40 μL ProteinA/G,在4℃孵育3 h;离心沉淀ProteinA/G,加入等体积的2 × SDS 上样缓冲液,沸水中煮10 min 后进行SDS-PAGE 凝胶电泳,并用相应抗体进行Western Blot 检测。
1.2.5 荧光信号强度分析
对免疫染色后的幼虫脑及腹神经索(ventral nerve cord,VNC)在相同参数下进行共聚焦成像,用Image J 检测所得图像中靶蛋白及参照蛋白的荧光信号强度,计算相对荧光强度比值并进行标准化处理。所有荧光强度分析均进行3 次以上独立实验。
1.2.6 NMJ 形态定量分析
三龄幼虫体壁肌肉经HRP 和DLG 抗体共染后,显示出NMJ 形态。对每种基因型随机选取20~ 30 只幼虫,对每只幼虫A3 体节的NMJ4 进行激光共聚焦成像,用Image J 软件测量每个NMJ 的突触分枝长度并计数其所含全部Ⅰ型bouton 个数。
1.3 统计学分析
使用Graphpad prism 5.0 软件进行统计分析。两组间比较使用Mann-Whitney 检验。不同基因型的NMJ 样本数列于所对应柱上。所有数据均以平均值± 标准差()表示,P<0.05 为具有显著性差异。
2 结果
2.1 DNlg4 与DNrx 互不影响彼此的蛋白水平及在突触区的分布
Nlg 与Nrx 过去作为一对相互作用的突触黏附分子被报道,体外实验显示Nlg 对突触前的诱导分化作用依赖于Nrx[17-18]。假定DNlg4 与DNrx 在突触区为相互作用的一对搭档分子,那么一个分子的缺失是否对另一个分子在突触区的分布有影响呢?为此,分别对dnlg4 突变体中DNrx 的表达以及dnrx突变体中DNlg4 的表达进行了免疫组化检测。由于已有的抗体在NMJ 无法探测到内源性DNlg4 和DNrx,检测了在幼虫神经中枢(包含脑及VNC)的分布和表达情况。结果显示,与对照相比,在dnlg4 突变体中,DNrx 能正常定位在幼虫脑部及VNC 的神经毡(neuronpil)(为突触密集区),且蛋白水平无改变,差异无显著性(见图1A~ 1C)。与此类似,DNlg4在dnrx突变体的分布和含量也没有明显改变(见图1D~ 1F)。说明两个分子各自的缺失对彼此在突触区的定位与蛋白水平没有影响。
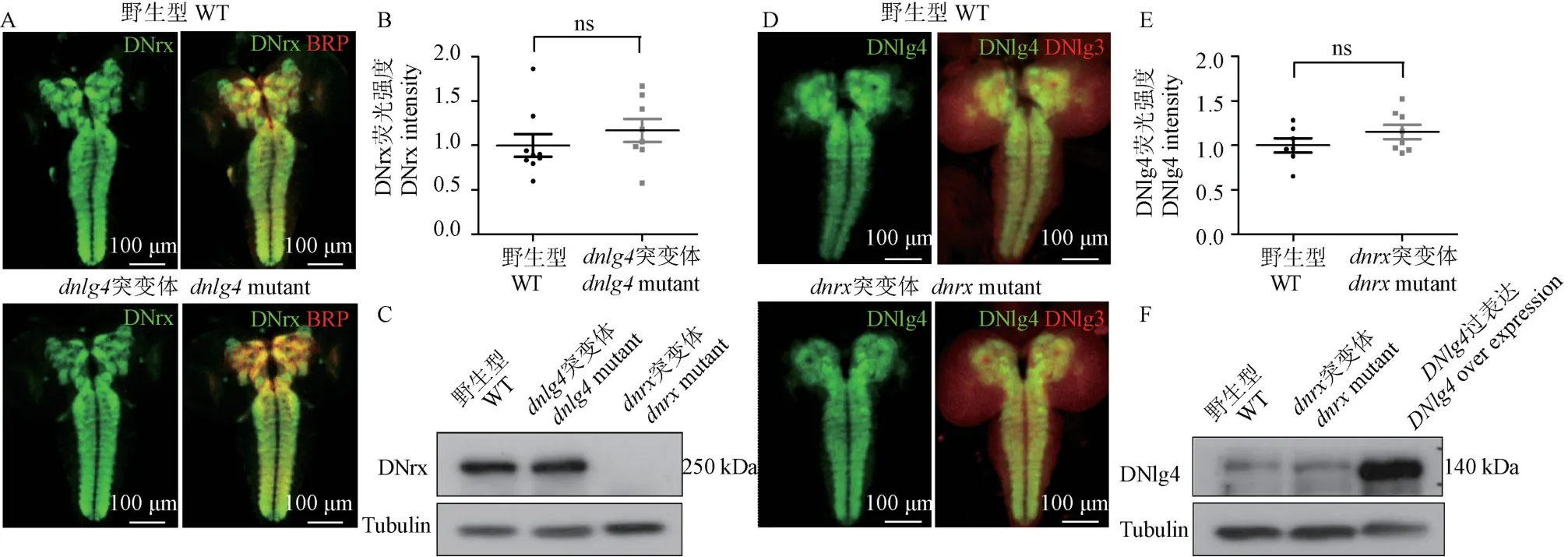
图1 DNlg4 与DNrx 不影响彼此在幼虫神经中枢突触区的蛋白水平与分布Note.A.The protein level and distribution of DNrx(green)in third-instar larval brain and VNC from wide types(WT) and dnlg4 mutants.BRP(red),a synaptic marker molecule is used as internal control.B.Quantification of the fluorescent intensities of DNrx in A experiment.C.Western Blot analysis for DNrx using larval brain extracts from WT and dnlg4 mutants.D.The protein level and distribution of DNlg4(green)in third-instar larval brain and VNC from WT and dnrx mutants.DNlg3(red)is used as internal control.E.Quantification of the fluorescent intensities of DNlg4 in Dexperiment.F.Western Blot analysis for DNlg4 using larval brain extracts from the WT and dnrx mutants.Figure 1 DNlg4 and DNrx do not affect the protein level and distribution of each other in the synaptic area of the central nervous system in Drosophila larvae
2.2 DNlg4 与DNrx 对NMJ 发育的调控作用程度不同
研究表明,DNlg4 与DNrx 对NMJ 发育的影响均有着正向的调控作用,两类分子的单独缺失均会导致幼虫NMJ 发育障碍,主要表现为NMJ 的bouton数量的减少[16,19]。为进一步明晰两种分子对NMJ发育的影响程度,分别对dnlg4和dnrx突变体的NMJ 进行了突触分枝长度的测量和bouton 的计数。结果显示,DNrx 缺失会导致严重的突触发育障碍,其突触分枝长度比野生型减少近34%,bouton 数目减少约36%。相对来说,DNlg4 缺失造成的NMJ 发育缺陷较DNrx 轻,dnlg4 突变体的NMJ 突触分枝长度比野生型减少约18%,bouton 数目减少约22%(见图2)。这反映了相比于DNlg4,DNrx 分子调控NMJ 发育的作用更大。
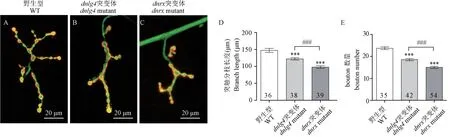
图2 DNrx 缺失导致比DNlg4 缺失更严重的NMJ 发育缺陷Note.A~ C.Representative morphology of NMJ4 in WT, dnlg4 and dnrx mutants.D,E.Quantification of branch length and bouton number at NMJ4 in the indicated genotypes.The number on the column represents the number of NMJ samples detected.Compared with the WT,∗∗∗P <0.001.Compared between the other two groups,###P <0.001.(The same in the following figures)Figure 2 Loss of DNrx leads to the more severe developmental defects of NMJ than DNlg4 deletion
2.3 DNlg4 与DNrx 对NMJ 发育的调控具有协同效应
DNlg4 和DNrx 对NMJ 发育均具有正向的调控作用,作为一对可能的搭档分子,那么二者是否是以同一路径调控突触发育呢? 为回答这个问题,整合制备了dnlg4,dnrx双突变果蝇,对其NMJ 发育情况与dnrx和dnlg4 单突变体分别进行了比较分析。假定DNlg4 和DNrx 沿同一路径调控NMJ 发育,由于dnrx突变体有相比dnlg4 突变体更为严重的发育缺陷,从基因效应来说DNrx 作用于DNlg4 的上游,DNlg4 对突触发育的调控将依赖于DNrx,因此,dnlg4,dnrx双突变体的NMJ 发育情况应该等同于dnrx单突变体。但结果显示,dnlg4,dnrx 双突变体呈现出较dnrx突变体更为严重的缺陷表型,其NMJ4 的突触分枝长度和bouton 数目相比dnrx突变体进一步减少,分别约为dnrx突变体的78%和75%(见图3)。这说明两个分子在调控NMJ 发育中是通过不同路径发挥作用,两个分子具有协同效应。

图3 dnlg4,dnrx 双突变体具有更严重的NMJ 发育缺陷Note.A~ C.Representative morphology of NMJ4 in dnlg4 mutants,dnrx mutants and dnlg4,dnrx double mutants.D,E.Quantification of branch length and bouton number at NMJ4 in the indicated genotypes.Compared with the dnlg4,dnrx double mutant,∗∗∗P <0.001.Figure 3 dnlg4,dnrx double mutants have more severe developmental defect in NMJ
2.4 DNlg4 可部分挽救因DNrx 缺失而产生的NMJ 发育缺陷
前期的研究结果显示,外源表达的DNlg4 可导致bouton 数量增加,促进NMJ 发育[16]。为进一步厘清两个分子在调控NMJ 发育中的相互关系,在dnrx突变体的背景下过表达DNlg4 以观察其对NMJ发育的影响。如果DNlg4 对NMJ 发育的调控依赖于DNrx,由于DNrx 的缺失,过表达的DNlg4 将不会引起bouton 的增生。实验结果显示,当用OK6-Gal4在dnrx突变体背景下驱动外源DNlg4 在运动神经元表达时,虽然NMJ 的突触分枝长度没有显著改变,但其bouton 数量相较dnrx突变体增加了约30%,尽管还达不到野生型水平。说明过表达DNlg4 可以部分挽救因DNrx 缺失造成的NMJ 发育缺陷(见图4)。

图4 DNlg4 可部分挽救因DNrx 缺失而导致的NMJ 发育缺陷Note.A~ E.Representative morphology of NMJ4 in indicated genotypes.F,G.Quantification of branch length and bouton number at NMJ4 in the indicated genotypes.Compared with the WT,∗∗∗P <0.001.Compared with the dnrx mutant,###P <0.001.Figure 4 DNlg4 partially rescue the developmental defects of NMJ caused by DNrx deficiency
2.5 DNlg4 与DNrx 不存在在体的相互结合
上述结果表明,DNlg4 与DNrx 在调控NMJ 发育中并不是以相互配对的搭档分子起作用,而是各自通过不同的路径发挥作用。两个分子的蛋白互作检测结果也佐证了这一点。对成虫头部组织进行裂解后,利用DNlg4 和DNrx 的多克隆抗体,通过免疫共沉淀方法检测了两个分子是否在体内形成结合。结果未检测到DNlg4 与DNrx 有共沉淀现象,说明两个分子不存在在体的相互结合。
3 讨论
Nlg 最早作为Nrx 的配体于1995 年被首次报道[8],大量体外和在体研究表明Nlg 与Nrx 为一对相互作用的突触黏附分子,其中,Nrx 主要定位于突触前,Nlg 则定位于突触后,通过其胞外端的相互作用,在突触的形成、成熟和神经传导中发挥着重要作用。但对线虫和果蝇Nlg 的研究显示Nlg 在神经元的定位不完全局限在突触后,某些Nlg 成员在突触前和突触后可能都有分布[10-12]。另有报道表明,跨突触的Nrx/Nlg 结合并不是这两个分子唯一的作用形式,Nlg 与Nrx 可在突触后形成顺式的结合(cis-binding)[11],这些结果增加了Nlg 与Nrx 作用的复杂性。
对果蝇DNlg1 和DNrx 的研究表明,突触后DNlg1 对NMJ 发育的影响依赖于突触前DNrx,二者有着直接的在体相互作用[13]。而前期对果蝇DNlg4 的研究证明DNlg4 主要定位于NMJ 突触前,作为突触前分子调控NMJ 发育[16]。DNlg4 的这种“异常”的定位引出一个问题,即DNlg4 对NMJ 发育的影响是否与突触前DNrx 有关,本研究的目的即在于解决这一问题。
对相互作用的膜蛋白分子,一个分子的缺失可能会影响其结合的分子在膜上的定位与稳定,例如,在前期研究中发现DNlg4 可结合另一个突触前信号蛋白Tkv,当DNlg4 缺失时,Tkv 在突触区的蛋白水平显著降低,显示了DNlg4 对Tkv 在突触稳定存在的重要性[16]。基于此,分别在dnrx和dnlg4 突变体中检测DNlg4 和DNrx 在幼虫头部突触密集区的蛋白水平,结果未检测到两个分子有显著变化,表明DNlg4 和DNrx 不会影响彼此在突触区的定位与稳定。此前的研究分别报道过DNrx 和DNlg4 缺失会造成NMJ 发育缺陷,为进一步明晰两种分子对NMJ 发育的影响程度,比较了dnrx和dnlg4 突变体NMJ 的突触分枝长度和bouton 数量。结果显示DNrx 缺失会导致比DNlg4 缺失更严重的NMJ 发育缺陷,反映了DNrx 相比DNlg4 具有更大的突触发育调控作用。检测两个相关分子是否具有遗传学相互作用是揭示其是否以同一路径起作用的有效手段。由于两个分子缺失均会造成NMJ 发育缺陷,因而检测两个分子同时缺失时NMJ 的发育状态。结果在dnlg4,dnrx双突变体里观察到了比任一基因单突变更短的突触分枝和更少的bouton 数量,这种表型的累加效应说明两个分子对突触发育的影响有着协同作用,但不完全在同一路径。这个结果得到了挽救实验的进一步验证。当在dnrx突变体运动神经元中过表达DNlg4 时,发现dnrx突变导致的NMJ 发育缺陷可部分被外源表达的DNlg4 挽救,反映了在DNrx 缺失状态下,DNlg4 仍然可促进NMJ发育。这进一步说明DNlg4 对NMJ 发育的调控不依赖于DNrx。值得注意的是在这一结果中,DNlg4可部分挽救DNrx 缺失导致的bouton 数量减少,但却不能挽救其导致的突触分枝变短这一表型,反映了在NMJ 的发育过程中轴突延伸和bouton 的形成是两个相关却并不完全同步的行为,bouton 的形成比轴突延伸对DNlg4 的作用更为敏感。与这种遗传互作结果相一致,并未检测到两个分子存在在体的相互结合。基于以上结果,认为DNlg4 对神经肌肉突触发育的调控以一条不同于DNrx 的信号路径起作用,DNlg4 在正向调控神经肌肉突触发育过程中不依赖于DNrx。本研究结论丰富了对Nlg 和Nrx 的分子功能的认识,即二者除了以跨突触配对结合的黏附分子发挥突触粘附作用外,还存在各自独立的信号分子的作用。
需要注意的是本研究是以NMJ 为突触模型进行的,所以这一结论目前尚限于对神经肌肉突触发育的调控。至于两种分子在神经中枢突触发育调控中是以何种方式进行,尚有待进一步的研究。
猜你喜欢
学苑创造·A版(2022年3期)2022-03-29
烟台果树(2021年2期)2021-07-21
学苑创造·A版(2019年6期)2019-07-11
测控技术(2018年7期)2018-12-09
金色少年(奇趣科普)(2017年4期)2017-06-05
安徽医科大学学报(2016年12期)2017-01-15
山东农业工程学院学报(2016年6期)2016-12-01
农村农业农民·B版(2016年7期)2016-10-21
天津医科大学学报(2015年2期)2015-12-22
山东医药(2015年40期)2015-02-28
