
番茄花序梗部长度的遗传分析
2023-09-26 01:04李艳琪曹晓宇李博宇王亦希张德楷战祥强胡体旭
中国蔬菜 2023年9期
李艳琪 曹晓宇 李博宇 王亦希 张德楷 战祥强 胡体旭
(西北农林科技大学园艺学院,陕西杨凌 712100)
番茄(SolanumlycopersicumL.)是茄科番茄属的一年或多年生草本植物,因其适应性强、产量高,在全世界得到了广泛的栽培。2020 年世界番茄总产量为18 682.1 万t,中国产量为6 486.6 万t(数据来源于联合国粮食及农业组织),位居世界第一。
花序的结构、数量、长度对于番茄果实产量的影响较大。在品种选育时,根据育种目标的果实大小选择不同花序类型的亲本进行配组,使得目标品系的花序类型与果实大小相匹配,从而达到理想的产量与收益。如大果番茄的单果质量较大、果形较大,所以大果番茄的理想花序类型是分枝较少、每穗花序的花数较少、花序后梗部长度短,可以保障番茄的营养供给和果实品质,防止花序梗部被压断的情况发生;小果番茄的单果质量较小、果形较小,因此适宜选择分枝多、每穗花序的花数多、花序总梗长较长的花序类型,以提高产量,增加效益(王晓甜,2021)。番茄作为模式植物,对其花序分枝数量、结构和发育的研究较多。SlSTM3与番茄的花序分枝数量有关,SlSTM3 蛋白通过激活SlFUL1转录从而增加分枝数量,SlSTM3 与花序分枝负调控因子J2 形成STM3-J2 复合物,在调控花序分枝方面发挥了作用(Wang et al.,2021)。植物的茎尖分生组织(SAM)在生长和分化的动态平衡下,产生地上器官,并在营养生长期产生芽、叶、茎和叶腋分生组织(AM),在茎尖分生组织获得繁殖能力时转变为花序分生组织(IM)(Yuste-Lisbona et al.,2016)。番茄的IM 在分化成花分生组织(FM)之前,在IM 的侧边形成1 个新的IM,番茄花序结构变化由起始次生分生组织(SM)和FM 决定(Wang et al.,2018)。拟南芥花发育的一系列具有意义的活动可分为12 个阶段(Smyth et al.,1990)。有报道称SlAG(AGAMOUS)在花发育的不同阶段动态调节SlARF3表达,在花发育的3~4 阶段,SlAG 间接抑制SlARF3表达,对FM 起维持作用;在花发育的5~6 阶段,生长素与SlAG 协同诱导SlARF3表达,直接或间接抑制SlLOGs、SlAHK4和SlIPTs基因的表达,减少细胞分裂素的生物合成和活性,控制FM 维持期间的细胞周期基因和细胞分裂,阐明了FM 中的干细胞维持机制(Zhang et al.,2018)。研究发现,SlDOF9在IM 和FM 表达量较高,将SlDOF9敲除后,突变体产生更多花,促进了SIM 的分化,产生更多的FM,进而增加产量,相反SlDOF9过表达导致植株花的数量变少,SlDOF9可通过调控花序结构调节因子(LIN)以及生长素和细胞分裂素(CK)有关基因从而影响IM 和FM 的分化,起到控制番茄花序发育的作用(Hu et al.,2022)。SlmiR156 通过抑制7 个SlSPL基因的表达,在番茄的生长发育中发挥关键作用,其转基因植株相比于野生型许多性状发生了变化,产生了额外的营养花序芽,叶片变小,节间长度变短,节数增加,果实变少、变小;继续研究发现番茄SlSFT和SlSP基因表达水平的比率降低导致沉默Sly-miR156a 转基因番茄营养花序芽增多(Zhang et al.,2011)。进一步研究发现,Sly-miR156a 的主要靶基因是SlSPL13,SlSPL13可以促进开花基因SlSTF表达,从而调控番茄花序形态发生(Cui et al.,2020)。
遗传分析已经应用在番茄的多个性状中,前人研究以节间长度有差异的串黄和大黄番茄为亲本,构建6 世代遗传群体,对节间长度进行遗传分析发现,番茄的节间长度符合MX2-ADI-ADI 模型(张宁 等,2022)。也有研究报道番茄长花柱的最适遗传模型为MX2-ADI-AD,并定位出10 个QTL 与番茄长花柱相关(马雅琳 等,2020)。还有研究显示番茄封顶花序数为MX2-ADI-AD 模型(张娜,2020)。花序分枝的长短影响番茄的机械化采收、坐果率、果实品质,最终对农业生产力造成影响,但是针对番茄花序梗部长度的遗传分析鲜有报道,本试验以Ailsa Craig(AC)和LA1964 为亲本,构建4 世代群体,对其花序梗部长度进行遗传分析,旨在明确番茄花序梗部长度的遗传规律,为番茄花序梗部长度的基因定位提供参考,同时也为番茄品种选育提供依据。
1 材料与方法
1.1 试验材料
将花序梗部长度分为花序前梗部与花序后梗部,前梗部与后梗部之和为花序总梗长,如图1所示。
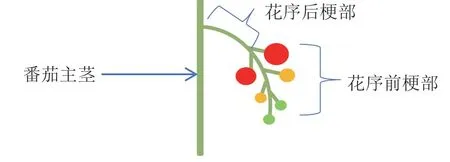
图1 番茄花序梗部长度示意图
亲本材料为栽培品种AC(母本P1)和类番茄茄(Solanumlycopersicoides)LA1964(父本P2),AC 的花序后梗部较短,LA1964 的花序后梗部较长(图2)。二者杂交获得F1,F1自交获得F2,由此构成4 世代群体。其中,母本AC 的花柱长度≤雄蕊,可正常自花授粉结果;父本LA1964 及F1的柱头外露且自交不亲和,人工异株授粉后结果;F2分离群体柱头均外露,无法自花授粉结果,需要人工辅助授粉。供试材料均由西北农林科技大学园艺学院蔬菜逆境生物学课题组提供。
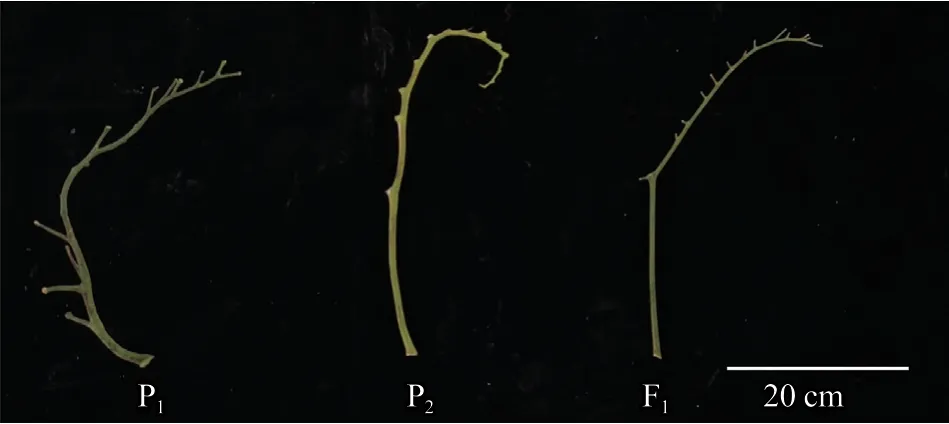
图2 番茄亲本及F1 花序梗部对比
1.2 试验方法
供试材料于2022 年1 月播种,3 月定植于陕西省杨陵区曹辛庄试验农场,株距35 cm,行距50 cm,按常规番茄种植方法进行田间管理,未进行人工摘心工作,植株所有花序均保留。AC 定植24株,LA1964 和F1分别种植10 株。F2群体为320 株,田间序号从F2-1415 至F2-1734,待花序最前端花朵开放完毕,测量花序前梗部及后梗部的长度,并完整剪下拍照。F2群体各花序有n 个分枝的频率计算公式:n 个分枝株数/ F2群体总株数(n 为2 或3以上)。
1.3 数据分析
将田间测量所得数据记录到Excel 软件内,采用SEA-G4F2(曹锡文 等,2013)软件包对本试验中4 世代联合群体第2、3、4 花序的总梗长、前梗长和后梗长进行遗传分析,根据赤池信息准则(Akaike’s Information Criterion,AIC)最小原则及模型适合性检验选出最优模型,估算其一阶遗传参数和二阶遗传参数。
2 结果与分析
2.1 番茄4 世代群体花序长度分析
由图3 可知,番茄两个亲本的花序后梗部长度差异较大,F1花序后梗部较长,偏向于亲本P2,且F1的花序总梗长从第2 花序到第4 花序越来越长,说明杂种优势逐渐明显。F2分离群体的花序梗部长度呈现连续性,符合数量性状特征。第2 花序有2 个分枝的频率为16.76%,有3 个分枝及以上的频率为1.18%;第3 花序有2 个分枝的频率为23.24%;有3 个分枝及以上的频率为3.24%;第4花序有2 个分枝的频率为41.47%,有3 个分枝及以上的频率为10.00%,第4 花序分枝数较第2、3花序明显增多。
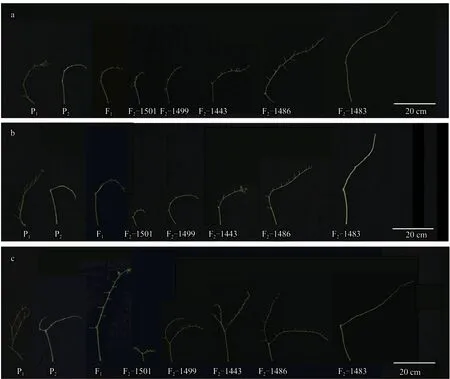
图3 番茄4 世代群体花序梗部对比
对番茄4 世代群体的花序长度进行统计分析发现(表1),父本P2的第2、3、4 花序后梗长分别为115.0、100.9、116.1 mm,显著大于母本;F1的第2、3、4 花序后梗长分别为126.4、139.7、191.7mm,均大于父本和母本,表现出超亲优势,且从第2 花序到第4 花序超亲优势越来越明显。
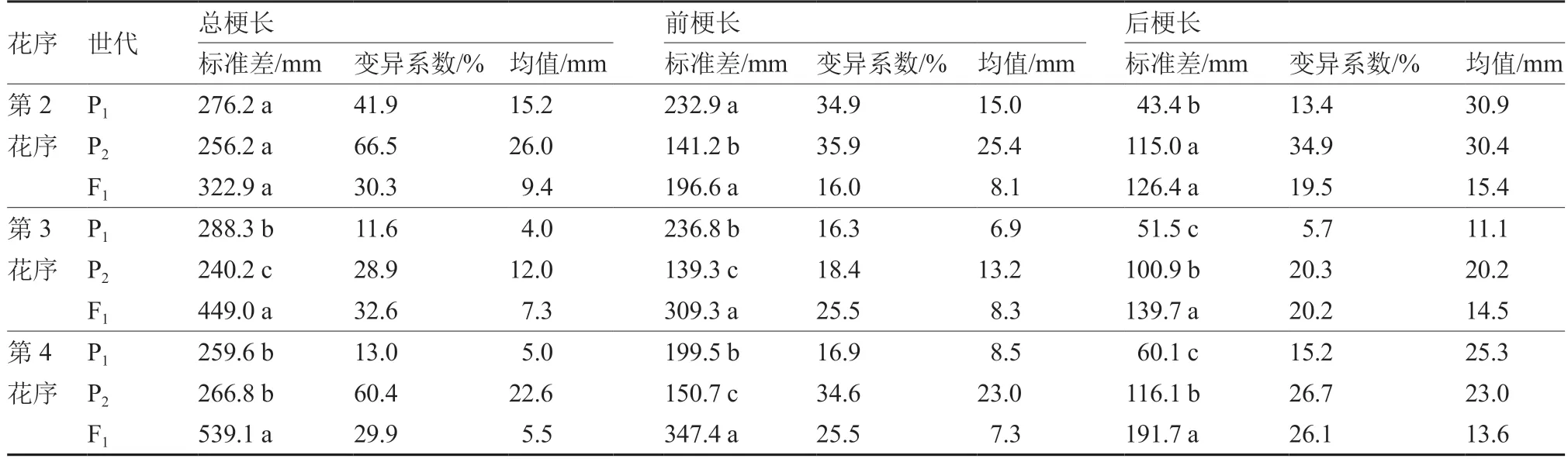
表1 番茄亲本及F1 群体花序梗部长度的基本参数
番茄F2群体的第2、3、4 花序后梗长介于双亲之间(表2),偏向于P2,F2分离群体的变异系数均大于P1、P2、F1且超过35%,说明F2的遗传变异性较强。
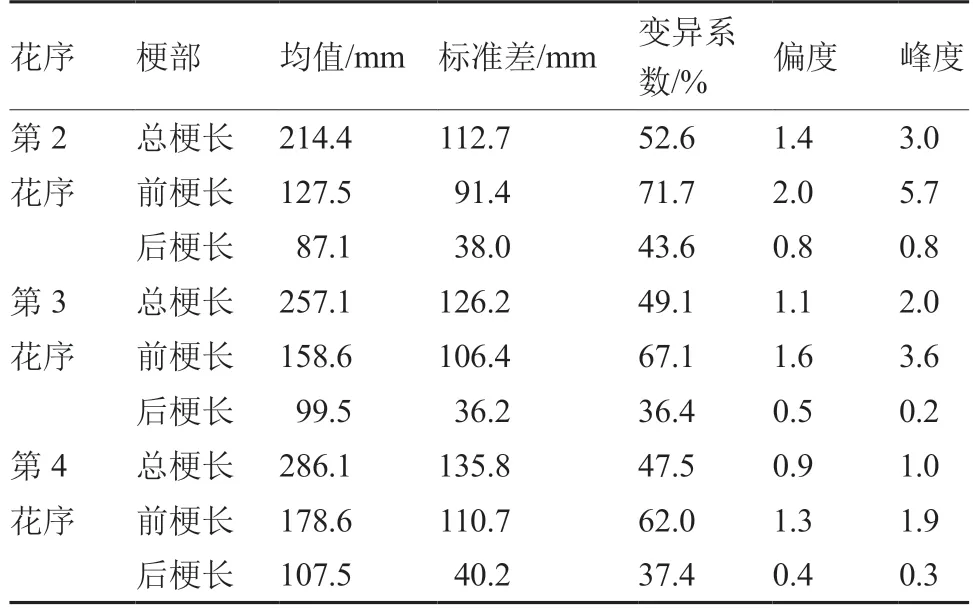
表2 番茄F2 群体第2、3、4 花序梗部长度的基本参数
番茄F2群体的第2、3、4 花序的总梗长、前梗长、后梗长均表现为连续的单峰分布(图4),表明番茄花序梗部长度相关性状均为数量性状,下一步可以进行主基因 + 多基因遗传分析。
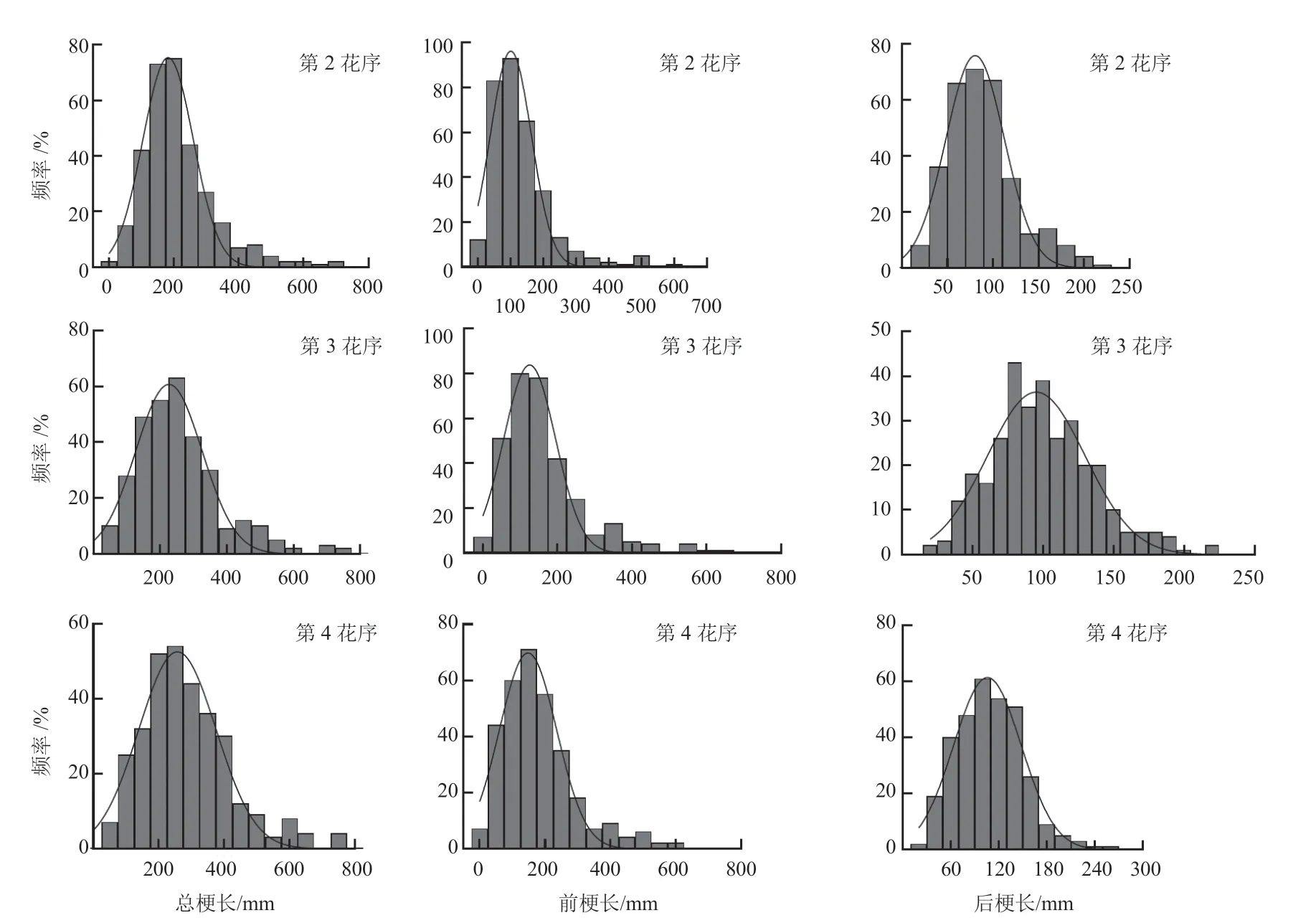
图4 番茄F2 群体第2、3、4 花序梗部长度频率分布
2.2 花序长度遗传模型分析
2.2.1 最优遗传模型的选择 利用SEA 软件包,对番茄F2群体的第2、3、4 花序长度进行4 世代联合分析,根据AIC 值最小原则,初步筛选出3个AIC 值较小的模型作为备选模型(表3),然后进行适合性检验,选择AIC 值最小且适合性检验的显著统计量少的模型作为最优遗传模型。
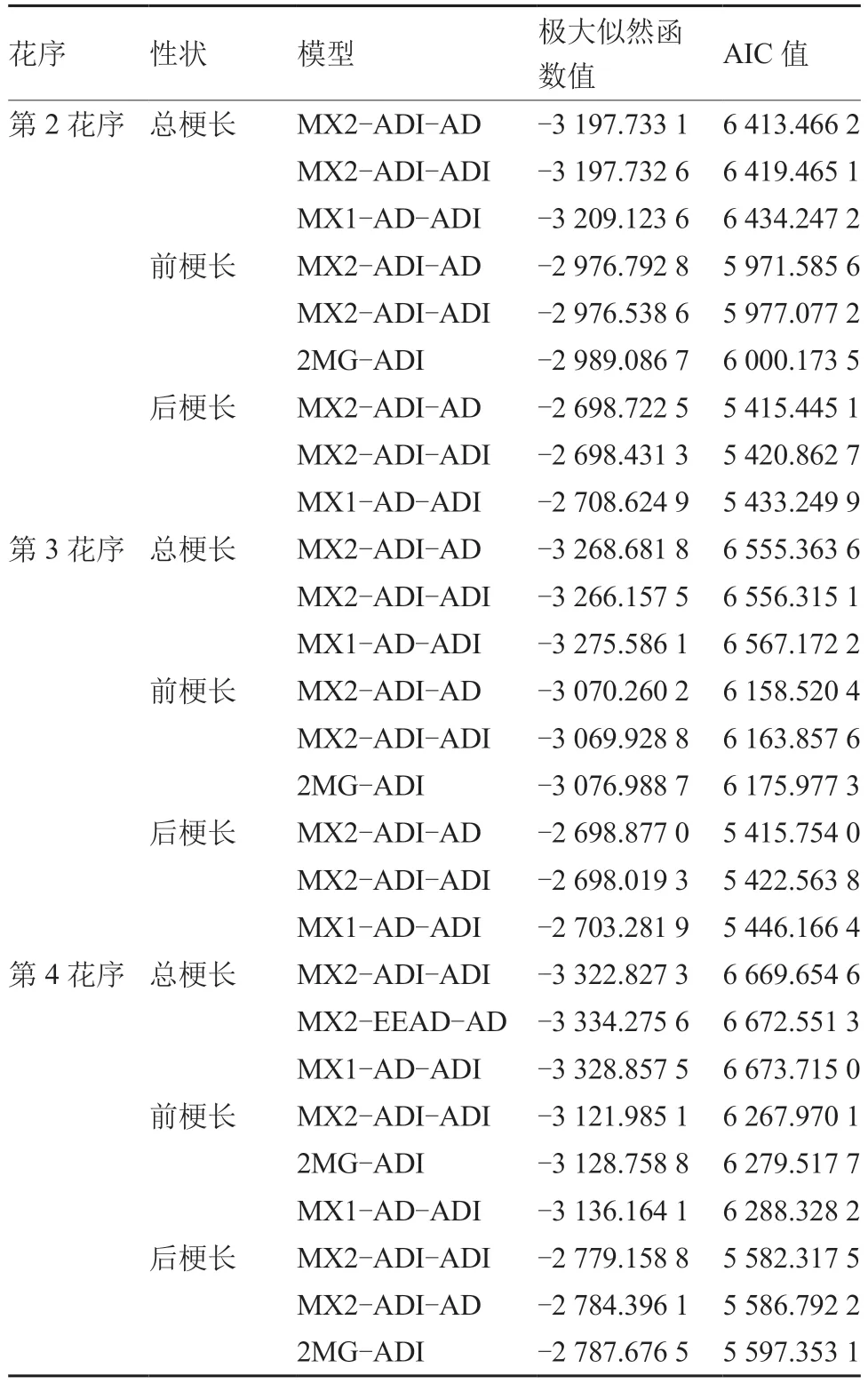
表3 番茄F2 群体第2、3、4 花序备选模型的极大似然函数值和AIC 值
为明确第2 花序的最适遗传模型,对其总梗长、前梗长、后梗长的3 个备选模型分别进行适合性检验(表4),第2 花序总梗长的3 个备选模型统计量的显著水平数量均为3 个,通过比较AIC值可知,MX2-ADI-AD 模型的数值最小,因此第2 花序总梗长的最优模型为MX2-ADI-AD;第2花序前梗长的3 个备选模型适合性检验均有6 个差异显著统计量,经过比较AIC 值,MX2-ADI-AD模型的数值最小,因此第2 花序前梗长的最优遗传模型为MX2-ADI-AD;第2 花序后梗长的MX2-ADI-AD 和MX1-AD-ADI 模型均有6 个差异显著统计量,前者的AIC 值较小,MX2-ADI-ADI模型有8 个差异显著统计量,应排除,因此第2 花序后梗长的最优遗传模型为MX2-ADI-AD。综上,第2 花序的总梗长、前梗长和后梗长最优遗传模型均为MX2-ADI-AD。
由表5 可知,第3 花序总梗长的MX2-ADI-AD和MX1-AD-ADI 模型,均有5 个参数达到显著水平,其中MX2-ADI-AD 模型的AIC 值最小,MX2-ADI-ADI 模型有6 个参数达到显著水平,应排除,故第3花序总梗长最优模型为MX2-ADI-AD。第3 花序前梗长的备选模型差异显著统计量均为5 个,且MX2-ADI-AD 模型AIC 值较小,因此MX2-ADI-AD 作为第3 花序前梗长的最优遗传模型。第3 花序后梗长的MX2-ADI-AD 模型和MX2-ADI-ADI 模型均有7 个参数达到显著水平,前者的AIC 值较小,而MX1-AD-ADI 模型有8 个参数达到显著水平,应排除,故MX2-ADI-AD 模型为第3 花序后梗长的最优遗传模型。综上,第3花序的总梗长、前梗长和后梗长的最优遗传模型均为MX2-ADI-AD。
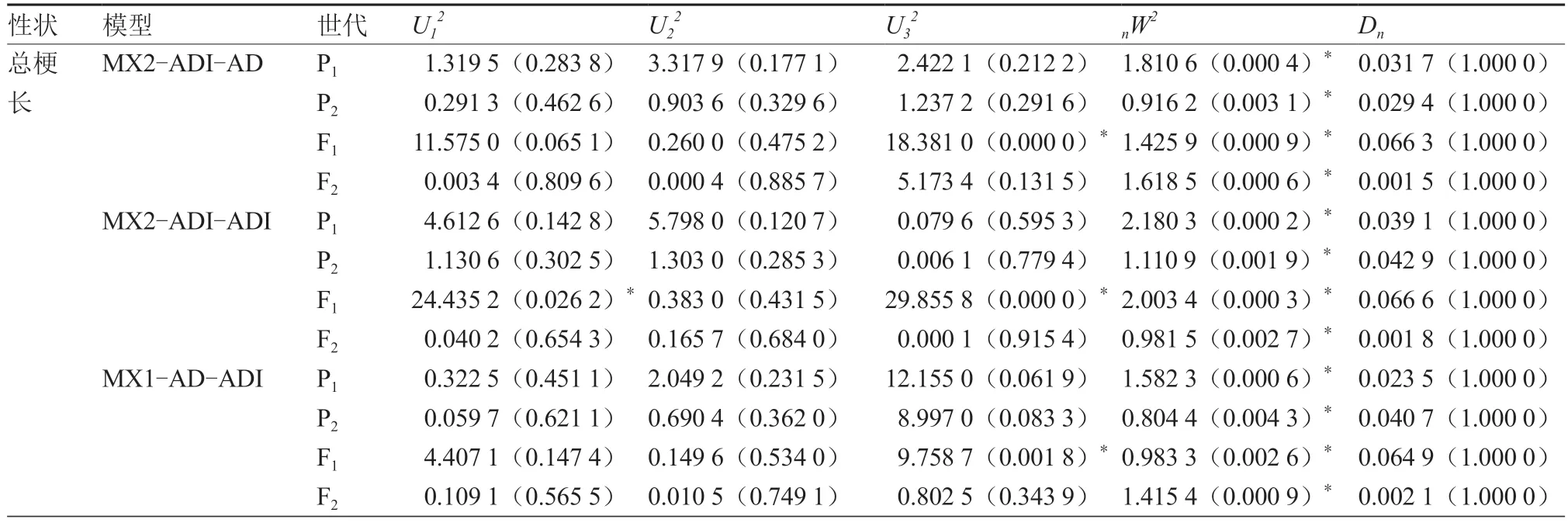
表5 番茄第3 花序备选模型适合性检验
番茄第4 花序总梗长的MX2-ADI-ADI 和MX1-AD-ADI 模型差异显著统计数量均为5,前者的AIC 值较小,MX2-EEAD-AD 模型差异显著统计数量为8,应排除(表6),故MX2-ADI-ADI为第4 花序总梗长的最优遗传模型。第4 花序前梗长的MX2-ADI-ADI 和2MG-ADI 模型差异显著统计数量均为4,前者的AIC 值最小,MX1-AD-ADI模型的差异显著统计数量为5,应排除,故将MX2-ADI-ADI 作为第4 花序前梗长的最优遗传模型。第4 花序后梗长的MX2-ADI-ADI 和2MG-ADI模型差异显著统计数量均为7,前者的AIC 值较小,MX2-ADI-AD 模型差异显著统计数量为9,应排除,故第4 花序后梗长的最优遗传模型为MX2-ADI-ADI。综上,第4 花序的总梗长、前梗长和后梗长的最优遗传模型均为MX2-ADI-ADI。
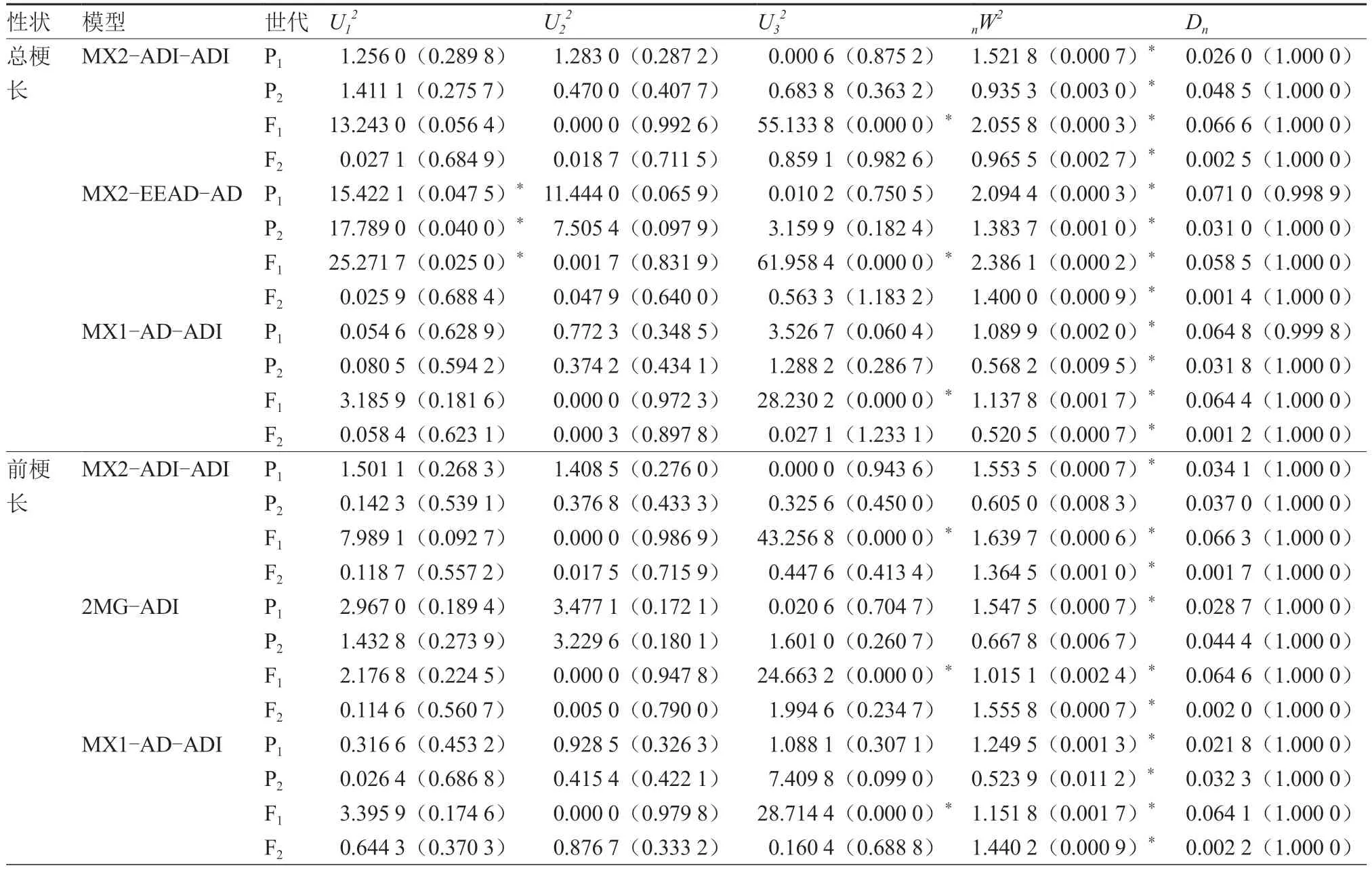
表6 番茄第4 花序备选模型适合性检验
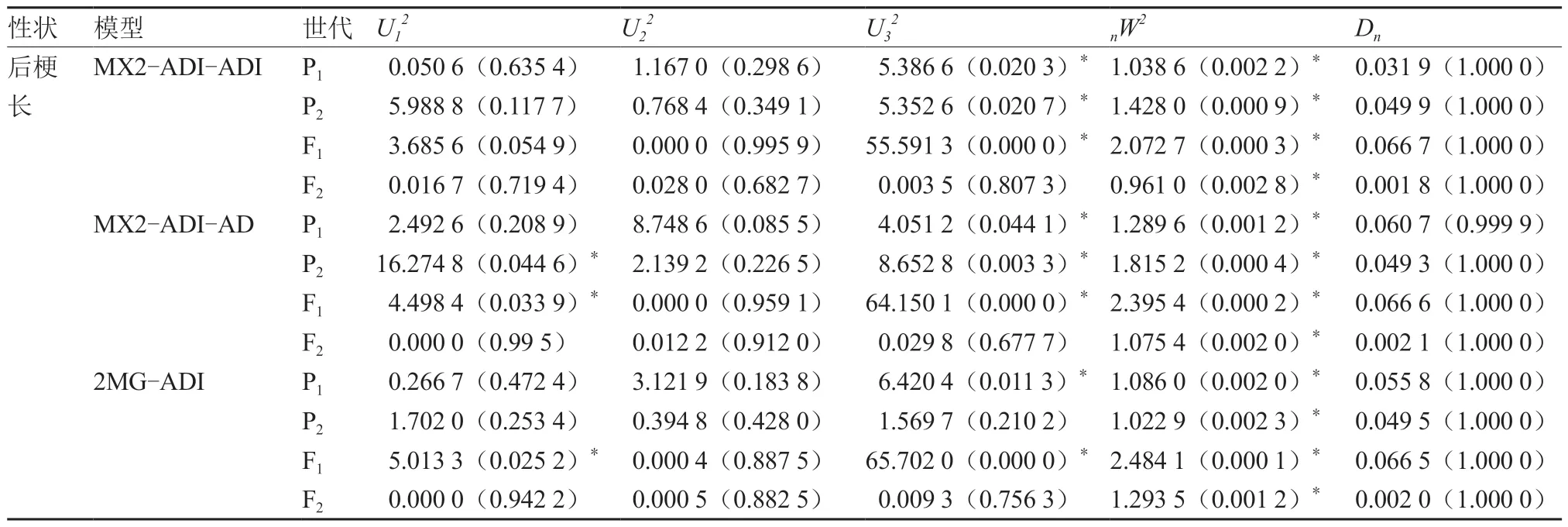
续表
2.2.2 番茄花序长度遗传参数估计 根据番茄第2、3 花序总梗长的最优遗传模型MX2-ADI-AD,第4 花序的最适模型MX2-ADI-ADI,通过SEA 软件得出第2、3、4 花序总梗长的最优遗传模型的一阶和二阶遗传参数估计值(表7)。第2、3、4 花序总梗长均由两对加性-显性-上位性主基因和多基因共同控制,主基因遗传率为46.61%~66.36%,多基因遗传率均为0。第2 花序的显性度为(ha/da:-1.00;hb/db:-0.99),表明第1 对主基因是负向完全显性,第2 对主基因接近负向完全显性,上位效应相差不大,主基因遗传率为46.61%。第3花序2 对主基因的加性效应均为正向,两对主基因的显性效应相等且均为正向增效,在上位效应中,加性×显性效应较大,主基因遗传率为53.19%。第4 花序总梗长的加性效应和显性效应均为负向增效,在上位效应中,显性×显性互作较为明显,主基因遗传率为66.36%。说明花序总梗长在一定程度上受主基因控制。
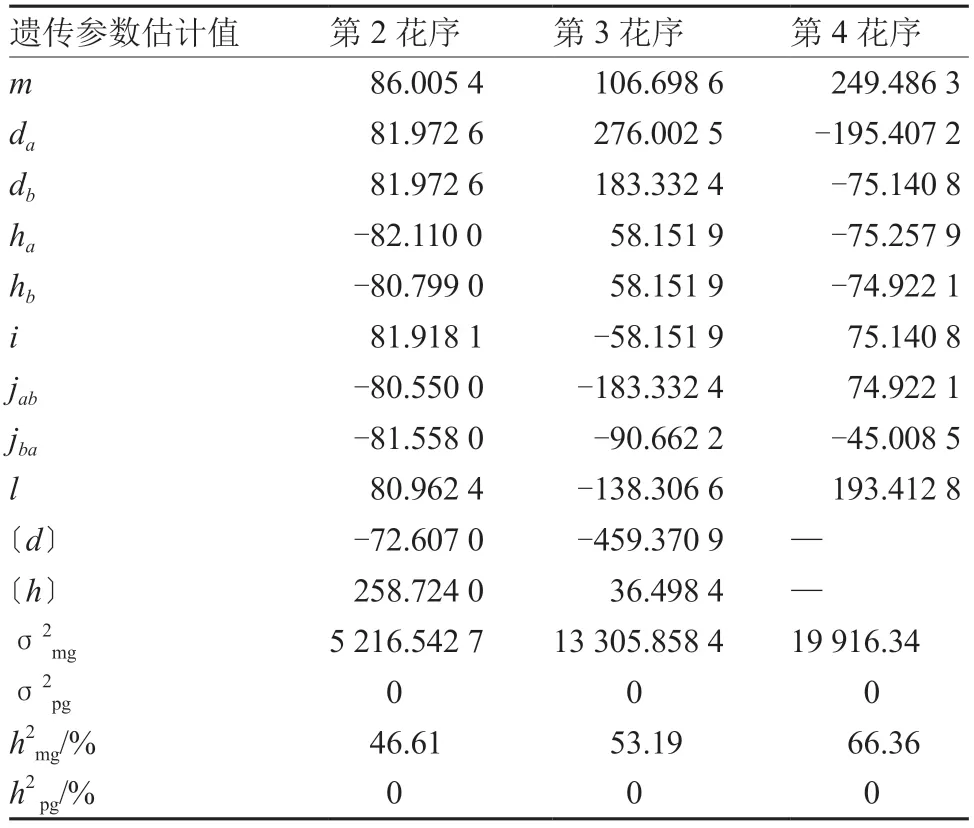
表7 番茄花序总梗长遗传参数估计值
第2、3 花序前梗长的最优遗传模型均为MX2-ADI-AD,第4 花序前梗长的最优遗传模型为MX2-ADI-ADI,对以上模型进一步分析,得出花序前梗长遗传参数估计值(表8)。第2、3 花序的2 对主基因加性效应均为正向,第4 花序的显性度为(ha/da:-0.46;hb/db:-0.98),说明第1 对主基因为负向部分显性,第2 对主基因接近负向完全显性。第2、3 花序的加性×显性互作效应(jba)较大,分别为-182.031 8 和-193.036 0,第4 花序的加性×加性效应(i)较大为60.256 9。第2、3、4 花序的主基因遗传率分别为53.72%、55.03%、62.53%,多基因遗传率均为0,表明花序前梗长主要由主基因控制。
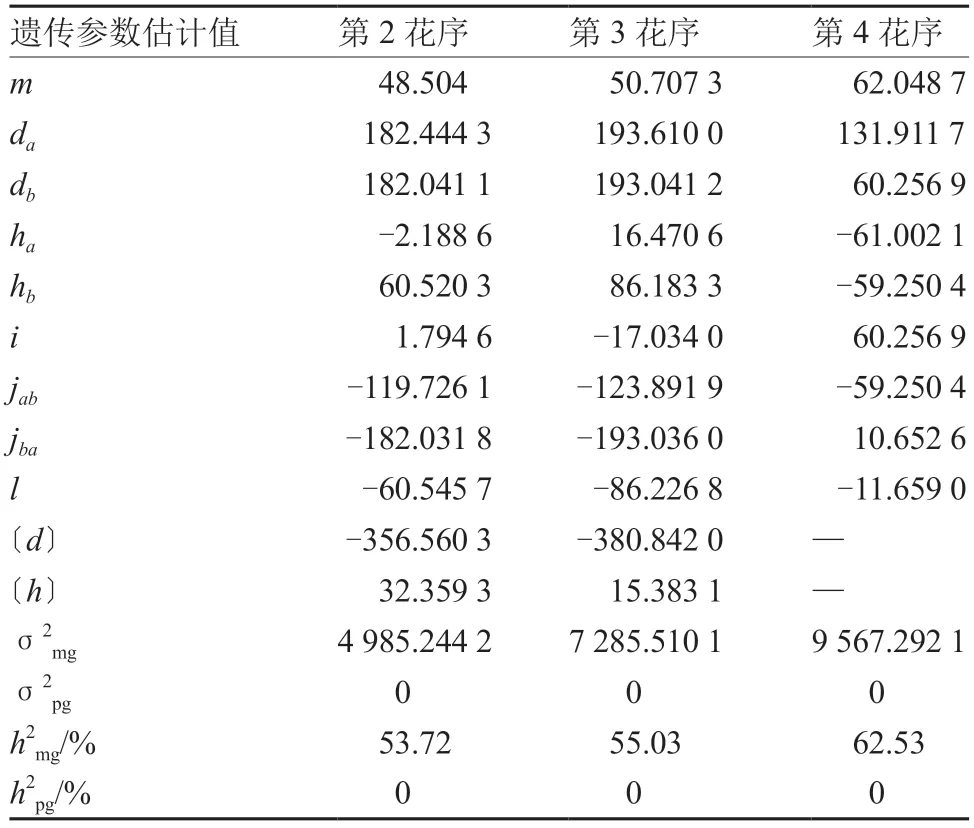
表8 番茄花序前梗长遗传参数估计值
第2、3 花序后梗长的最优遗传模型均为MX2-ADI-AD,第4 花序后梗长的最适遗传模型为MX2-ADI-ADI,对它们进行遗传参数的估计(表9)。第2、3 花序的2 对主基因加性效应均为正向,第1 对主基因的加性效应分别为106.865 7和89.008 0,大于第2 对主基因的69.900 4 和45.784 8,显性效应均为正向增效;在上位性效应中,第2 花序2 个主基因间加性×显性互作效应(jab)最强,其数值为-69.808 9,显性×显性互作效应(l)最弱,其数值为8.915 3。第3 花序的加性×加性互作最强,达到-45.813 0。第4 花序的加性和显性效应均为负向,以第1 对主基因的加性效应为主。第2、3、4 花序的主基因遗传率分别为71.45%、77.08%和64.51%,多基因遗传率均为0,说明花序后梗长的遗传中主基因起较大作用。
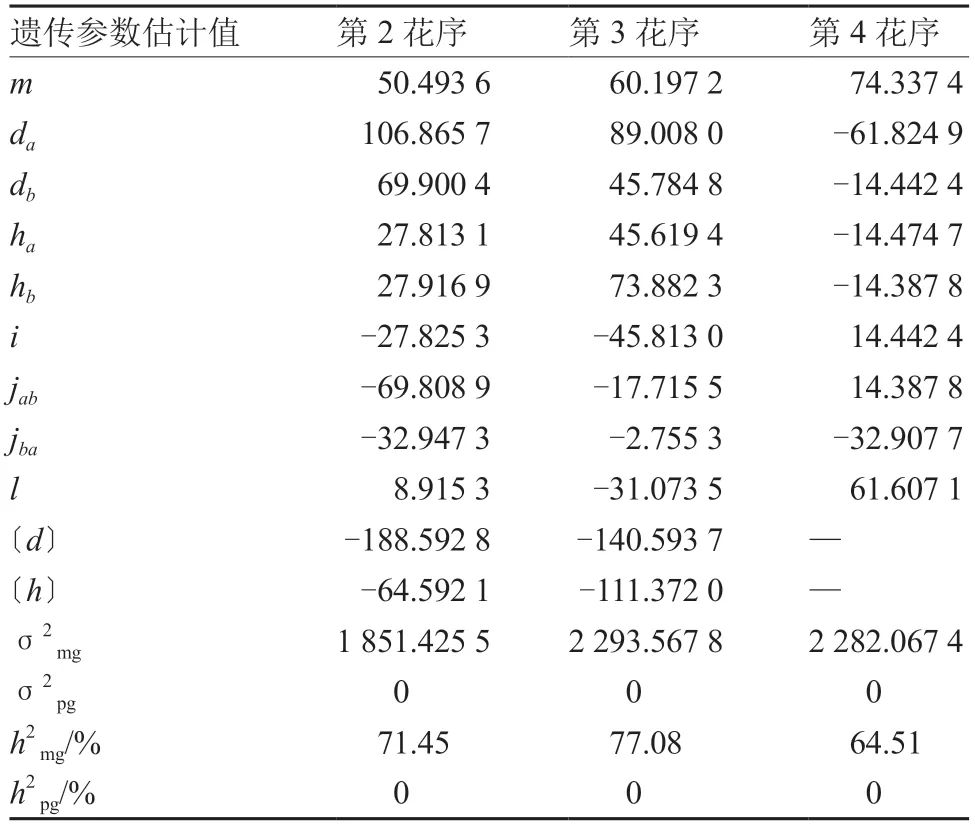
表9 番茄花序后梗长遗传参数估计值
3 讨论
盖钧镒等(2003)根据现代遗传学已有的研究基础,利用极大似然原理、最小二乘法、AIC 值准则、适合性检验等原理,总结出一套主基因 + 多基因的遗传体系用于植物数量性状的分析方法。在2013 年由曹锡文研发出SEA 软件包,界面简洁易操作(曹锡文 等,2013),受到了遗传研究者的广泛使用。目前该方法已应用于番茄萼片形态性状(王晶 等,2020)、茄子株高性状(杨锦坤 等,2019)及紫色果皮着色深度性状(刘娅萍,2020)、黄瓜果实空心性状(周赓 等,2021)及果刺大小性状(狄胜强 等,2018)、甜瓜果实糖组分含量性状(叶红霞 等,2019)、苦瓜种子大小和单粒质量性状(邹一超 等,2020)、枣杂交群体花表型主要性状(杨植 等,2023)等园艺作物主要性状的遗传分析中,表明SEA 软件包可明确相关性状的遗传作用,同时为相关基因定位提供参考。
番茄花序长度影响番茄坐果率和机械化采收等,是影响产量的一个重要性状。本研究以花序后梗部较短的AC 和花序后梗部较长的LA1964 作为亲本,构建4 世代遗传群体,花序梗部长度的差异显著性在第2、3、4 花序上均有不同,说明番茄花序梗部长度是数量性状,环境对其有一定影响。对第2、3、4 花序的总梗长、前梗长和后梗长进行遗传分析,结果表明:第2 花序和第3 花序的总梗长、前梗长、后梗长符合MX2-ADI-AD 模型,第4 花序总梗长、前梗长、后梗长的最适遗传模型为MX2-ADI-ADI,说明番茄花序梗部长度均由2 对加性-显性-上位性主基因 + 多基因控制,不同之处在于是否存在多基因的上位性效应。初步判断是由于温度较高时,多基因存在上位性效应;温度适宜时,多基因没有上位性效应,或者遗传机理较为复杂,具体原因有待进一步探索。主基因效应的作用方式在不同花序之间,均有差异,证明番茄花序梗部长度遗传机理较为复杂。番茄第2、3、4 花序梗部长度的主基因遗传率为46.61%~77.08%,多基因遗传率均为0,说明番茄花序梗部长度的遗传以主基因贡献为主。基因位点的表型效应与环境的互作有关(向道权 等,2001),有研究表明甜瓜幼苗下胚轴长度因季节不同导致主基因的多种效应存在差异(李琼 等,2021),与本试验推测花序梗部长度多基因效应的作用方式存在差异的原因类似,均为环境因素。有研究报道,黄瓜花梗和花冠长度的多基因遗传率均为0(苗永美 等,2015),与本试验的花梗长度遗传分析结果相符。
因此,在明确番茄花序梗部长度的遗传效应后,可利用分子标记技术构建遗传连锁图谱,进行番茄花序梗部长度基因座位的探索,从而实现分子辅助育种,培育出优良的番茄品种,使其花序梗部长度有助于提高果实品质、产量和机械化采收率。
4 结论
番茄花序梗部长度是影响番茄产量的一个重要性状。本试验经遗传分析后得出的结论为:花序梗部长度符合2 对加性-显性-上位性主基因 + 多基因的遗传模型,主基因遗传率为46.61%~77.08%,多基因遗传率为0,表明主基因遗传在番茄花序梗部长度的遗传中起主导作用。
猜你喜欢
——植物学教材质疑(六)
生命科学研究(2022年1期)2022-03-14
消费导刊(2017年24期)2018-01-31
南风(2017年34期)2018-01-16
辽宁大学学报(哲学社会科学版)(2017年3期)2017-06-21
科学种养(2017年6期)2017-06-13
科学种养(2017年5期)2017-05-17
领导决策信息(2017年11期)2017-05-17
中国农资(2016年1期)2016-12-01
陕西画报(2015年3期)2015-03-11
中学语文(2015年27期)2015-03-01
