
我国特有植物菱叶凤仙花的繁育系统及其与同属植物的比较
2023-09-25 07:59郭银巧
西华师范大学学报(自然科学版) 2023年5期
林 祎,郭银巧,王 琼
(1.西华师范大学 a.生命科学学院,b.西南野生动植物资源保护教育部重点实验室,四川 南充 637009;2.中国农业出版社,北京 100125)
全世界凤仙花属(Impatiens)物种超过1 000种,中国目前记载有352种,其中特有种273种,约占全世界的1/4,大部分在西南地区,其中四川分布119种[1]。该属植物色泽艳丽,花型独特,着生方式和部位多样,普遍具有较高的观赏价值,也吸引了各类型的传粉者,被誉为“双子叶的兰花”[2]。植物繁育系统是指影响后代遗传组成的所有有性特征,包括花形态特征、开放式样、花部寿命、传粉者种类、访问频率和交配系统,其中交配系统是核心,指居群个体间的交配式样,即自交和异交状态的总和[3]。远缘异交的成本较高,自交会导致育性衰退,因此很多植物倾向于选择异交与自交动态共存的混合式交配机制,这利于植物种群在各种环境中生存扩展,是植物对时空、传粉媒介、环境及自身的一种折中的适应机制[3]。
凤仙花属植物的观赏性、入侵性等特性与其繁育系统密切相关。一般异交系花朵的观赏性大于自交系花朵,比如小花凤仙花(I.parviflora)传粉者少,具有高度自交的特性,其花朵色淡且小,而卡佩凤仙花(I.capensis)异交系花朵对花部特征的能量投入比同种自交系花朵高出100倍,其色泽艳丽,花型更大[4]。在物种入侵方面,一些自交性较强的凤仙花入侵力更强,在入侵早期可以快速产生大量自交后代顺利度过“瓶颈期”,因此对入侵种的繁殖性状和繁育系统进行分析,可以探明其种群扩散和分布趋势,并以此制定相关治理措施[5]。凤仙花繁育系统对物种保护亦有重要意义,比如I.grandis因生殖力低下和环境变化,在自然界面临生存威胁,研究这类濒危物种的繁育系统,可以探究其致濒机制,从而制定相应的保护计划[6]。在病虫害防治方面,有研究发现异交机制可能会溶解凤仙花的抗性基因,导致病毒感染性增强[7]。总之,研究凤仙花繁育系统可以为该属植物的适应性、物种形成机制、多样性的保护维持、外来入侵防控和濒危植物保护等研究提供重要的基础资料。
目前国内外对凤仙花繁育系统的研究众多,但迄今尚未发现对菱叶凤仙花(I.rhombifolia)繁育系统以及凤仙花属整体的繁育系统的研究报道。菱叶凤仙花模式产地为峨眉山,为四川特有种,本文通过测定其田间种群中的花部特征、花期寿命、传粉者种类及行为、花粉胚珠比(P/O值)、杂交指数(OCI),并结合人工授粉实验研究该植物的繁育系统,以此对该种进行更好的选种培育。另外,人们普遍将凤仙花以异交为主的观点作为共识[8],但Vervoort等[5]在2011年发现了高度自交的特殊种,因此本文期望通过收集并整理国内外已有的凤仙花繁育系统信息,验证大部分凤仙花兼性异交的结论,并从生境、传粉者等方面解释其产生的原因,以期更好了解凤仙花的生活史和进化史,探明凤仙花属植物对环境的适应性,对其引种驯化、野生资源开发提供参考。
1 材料与方法
1.1 研究地点
峨眉山位于四川盆地西南边缘,是四川凤仙花属植物的分布中心之一。该地最高海拔3 099 m,复杂多样的地形气候孕育了丰富的动植物资源。海拔500~2 500 m有14种凤仙花分布,其中12种为我国特有种[9]。该地气候类型为亚热带湿润季风气候,植被类型为常绿阔叶林,土壤类型为黄壤。山区云雾较多,日照较少,雨量较为充沛;平均气温为17.2 ℃,最热月(7月)的平均温度为26.3 ℃,最冷月(1月)的平均气温为7 ℃;年均降水量是1 555.3 mm,全年降水量的70%~80%集中在5—9月。
1.2 研究对象
菱叶凤仙花分布海拔为380~1 850 m,在峨眉山、都江堰等地均有分布,多生于河岸石缝、林下、水沟边、山坡路旁等[10],属一年生草本植物,叶片菱形或近菱形,两面无毛,花黄色,花期在8—11月。该植物养护成本低,观赏价值高,在野外成丛分布,绿期长,覆盖度高,长势均匀,花朵明黄色,可用来点缀草坪,亦可丰富林下景观,在城市绿化中具有巨大的潜力。此外,因菱叶凤仙花具匍匐型生长的特点,还可将其开发为悬垂盆栽类,作为室内观赏植物[11]。因而,该植物是一种极具开发潜力的野生花卉。
1.3 研究方法
1.3.1 实验地设置
为更好地选育野生种质资源,于2021年3月将生长在峨眉山中下部(29°36′16″N,103°21′62″E,海拔932 m)的野生菱叶凤仙花幼苗60株(高约10 cm且长势相同)带土移栽至山腰(29°34′39″N,103°16′59″E,海拔1 241 m)。原生地点位于林缘和河边,移栽地点为农用地,周围有农田和人工种植的竹林、板栗树。移栽地中,沿水平方向设置4个小区,每个小区放置15盆(花盆底面直径25 cm,高30 cm),每盆中移栽1株,盆间隔10 cm。因凤仙花喜阴湿的习性,故在实验地上方覆盖了透光度26.83%±0.66%的遮阳网。
1.3.2 花部综合特征及开花物候
8月24日—9月28日,随机挑选25朵花蕾,吊牌标记,观察开花进程,记录花蕾期、雄花期、雌花期,同时观测群体花期寿命。另外选取108朵花蕾进行套袋,开放后对其传粉通道的长、宽、高,蜜距长度,花蜜体积等进行测定。
1.3.3 访花昆虫及其行为
9月12—19日,每天8∶00—16∶00约8个小时,观察40朵花上的传粉者种类、数量和访问频率,访问频率为1 h内昆虫对目标花朵的访问次数。9月22日开始选择20朵花蕾进行标记,每天早上7∶00进行套袋,下午17∶00去袋,统计其坐果和结籽情况,以此推测其夜间传粉者的存在与否。9月28日开始拍摄昆虫访花视频,描述其访花行为,并用剪映专业版软件逐帧分析视频,记录单花访问停留时间。
1.3.4 花粉胚珠比的测定
9月1—5日,随机采集35朵花蕾,测P/O值:(1)用解剖针将花药和子房分离,将花药放入研钵内磨成匀浆,全部转移到离心管中,定容到4 mL。(2)将花粉溶液摇匀后,用微量加样器(Eppendorf Research Plus,0.1~2.5 μL,艾本德(上海)国际贸易有限公司)取1 μL溶液,用显微镜观察并统计花粉数,每朵花重复5次,取平均值并计算4 mL溶液中的花粉总数。(3)用解剖针解剖子房,统计单花胚珠数。(4)计算单花P/O值。据Cruden[12]的标准判断繁育系统:P/O值在2.7~5.4时,闭花受精;18.1~39.0时,专性自交;31.9~396.0时,兼性自交;244.7~2 588.0时,兼性异交;2 108.0~195 525.0时,专性异交。
1.3.5 杂交指数
2021年9月中旬,用数显游标卡尺(CD-15AX,精确度:0.01 mm,Mitutoyo Corporation)测108朵花传粉通道的长和宽,并观察雄蕊和柱头间的距离[13]。按照Dafni[14]的标准进行杂交系数判断:(1)花朵直径<1 mm记为0;1~2 mm记为1;2~6 mm记为2;>6 mm记为3。(2)雌、雄蕊在同一位置记为0;否则记为1。(3)雄蕊与雌蕊二者同时成熟或雌蕊先熟,记为0;雄蕊先熟,记为1。以上三者之和为OCI值。评判标准为:OCI=0时,繁育系统为闭花授精;OCI=1时,为专性自交;OCI=2时,为兼性自交;OCI=3时,为自交亲和,有时需要传粉者;OCI=4时,为异交为主且需要传粉者。
1.3.6 人工套袋授粉实验
套袋实验于2021年9月开始,先后选择约200朵花作5组处理(5个处理尽量在同一株):处理1,去雄套袋,即不授粉;处理2,不去雄套袋;处理3,人工自交,即去雄套袋,待花进入雌性期,授充足的自株花粉后套袋;处理4,人工异交,即去雄套袋,待花进入雌性期,授充足的异株花粉后套袋;处理5,自然授粉。待果实明显膨大和伸长时,统计各处理的坐果率、结籽率,并计算自交不亲和指数(ISI)和自动融合指数(SFI),其中人工自交与人工异交的坐果率比值为ISI值,ISI>1代表自交亲和,在0.2≤ISI≤1代表部分自交不亲和,ISI<0.2代表大部分自交不亲和,ISI=0代表完全自交不亲和。SFI指不去雄套袋与人工异交处理的坐果率比值,SFI>0.2代表允许自动自交和融合生殖[15]。
1.3.7 凤仙花属植物繁育系统资料收集以及数据分析
以Impatiens、breeding system等为关键词在百度学术、Happy Botanist、eFlora of India和中国植物志等数据库里搜集凤仙花属繁育系统信息。人工授粉实验数据经Excel预处理,用SPSS 26进行分析,用One Sample Kolmogorov-Smironv检验正态性,坐果率不满足正态分布,所以采用非参数独立样本检验比较不同处理组间的指标差异显著性;结籽率满足正态分布和方差齐性,采用单因素方差分析。所有数据以平均值±标准差表示。
2 结果与分析
2.1 花部综合特征及开花物候
菱叶凤仙花具营养繁殖的能力,成簇丛生。1株有3~5朵单花,1个花梗上有2朵单花,常存在开放时差,花梗旁有1枚叶片伴生。单花两侧对称,花内包含繁殖器官,是典型的雄蕊先熟。花蕾黄绿色,蜜距逐渐由弯曲变直(图1a)。刚开花时,花药完整,不久后花药开裂呈散粉状态,为雄性期(图1b)。散粉后雄蕊脱落,进入雌性期(图1c)。花内含1枚雌蕊,雌蕊接收到花粉后很快受精坐果,果实膨大后开裂弹出种子(图1d)。该植物花色明黄,无明显气味,蜜距细直,内含花蜜。传粉通道长为8.41±1.68 mm (n=108),宽5.69±0.06 mm

(n=108),高6.84±1.04 mm(n=108),蜜距长度为18.14±1.46 mm(n=108),花蜜体积为2.19±1.42 μL(n=105)。该植物花期在8—11月,单花寿命为5.12±0.16 d(n=25),其中雄性期寿命为2.04±0.16 d(n=25),雌性期寿命为2.00±0.17 d(n=25)。
2.2 访花昆虫及其行为
菱叶凤仙花的主要访花者为三条熊蜂(Bombustrifasciatus)、中华蜜蜂(Apiscerana),其次为短头熊蜂(B.breviceps)、小豆长喙天蛾(Macroglossumstellatarum),以及少数蝴蝶等(图2)。三条熊蜂和中华蜜蜂为数量较多且明显的花粉携带者,前者体型较大,头部为花粉沉积位,具有明显的传粉行为,后者体型较小,大多为盗粉者,少数为次级盗蜜者。短头熊蜂为盗蜜者,直接破坏蜜距获取花蜜。小豆长喙天蛾为国内罕见的害虫,具有“采花不携粉,采蜜不酿蜜”之称,其出现可能与外来花卉引种有关,外形与蜂鸟相似,访花时身体呈飞行姿态悬停在花外,仅将长口器伸入花中取蜜,访花时间极短,据计测其访问过的雌蕊上的花粉落置数,发现其传粉效果不明显。三条熊蜂访花停留时间平均为1.57±0.17 s(n=68),中华蜜蜂的访花停留时间为2.73±0.29 s(n=81)。整个实验地中蜂类传粉者(三条熊蜂和中华蜜蜂合计)对标记花朵的访问频率为0.10±0.01次·h-1(n=119)。夜间开放传粉的花,其坐果率和结籽率分别为30%和38%,说明存在夜间访花者,其种类和传粉行为需进一步调查。

2.3 花粉胚珠比
菱叶凤仙花单花花粉数量约为140 434.29±5 892.15粒(n=35),单花胚珠数量约为12.60±0.33个(n=35),因此该植物P/O值约为11 442.67±558.28(n=35),据Cruden[12]的标准判断该植物繁育系统属于专性异交。
2.4 杂交指数
菱叶凤仙花的传粉通道宽平均为5.69 mm,加上旗瓣、翼瓣,花朵直径明显大于6 mm,计为3;幼雌蕊被花丝紧紧包被,只有雄蕊脱落后才露出来,是典型的雄蕊先熟,计为1;雌雄蕊无明显空间隔离,计为0。因此菱叶凤仙花的OCI为4,按Dafni[14]的标准判断该植物繁育系统主要为异交,需要传粉者。
2.5 人工授粉实验结果
如表1所示:不去雄套袋和去雄套袋后的坐果率分别为5%和0,表明菱叶凤仙花具有低比率的自动自交能力,不存在无融合生殖现象。相比套袋处理,人工自交、人工异交和自然授粉显著提高了坐果率(P<0.001),说明缺乏传粉者等媒介时该种无法正常结实。人工异交和人工自交处理后坐果率无显著差异,表明该植物自交亲和性较高。相较于自然授粉,人工授粉并未显著提高菱叶凤仙花的坐果率,可能与实验地传粉者较丰富有关。ISI值为0.86,SFI值为0.05,显示菱叶凤仙花部分自交亲和,几乎无自动自交和无融合生殖的现象。除了去雄套袋组,所有组的结籽率差异均不显著(F=0.289,P=0.833)。

表1 菱叶凤仙花人工授粉实验结果
2.6 凤仙花属植物的繁育系统
搜集的36种凤仙花在5大凤仙花热点地区均有分布,其中印度11种,非洲2种,东南亚4种,喜马拉雅以东12种以及欧美洲7种。有27种凤仙花明确记载了生长在林缘、溪流边等潮湿荫蔽的微生境,因此即使36个种的宏观生境不同,但相似的微生境使得它们仍具有一定的可比性。 所有36种凤仙花都是需要传粉者的异交植物,其中30%属于高度异交或专性异交,自交亲和的占70%,自交亲和中能够自动自交的占24%,只有小花凤仙花具有高度自交性,传粉类群中仅有1种盗粉者。凤仙花的传粉者种类丰富,有熊蜂类、蜜蜂类、天蛾类、蝶类、蝇类、鸟类等,盗蜜者类群主要包括短头熊蜂、考氏无垫蜂(Amegillacaldwelli)、灰熊蜂(B.grahami)等。蜜蜂类传粉者在各热点分布区均有记载,为最广泛的传粉者,熊蜂类在欧美、日本、中国有分布,天蛾类和蝶类分布在印度、中国,鸟类在欧美、非洲,盗蜜者主要在中国、日本、东南亚分布。凤仙花种间的自然坐果率变化范围较大(8%~90%),这与当地传粉环境以及植物繁殖特性有关。平均坐果率为48.59%±0.05%,低于平均坐果率的凤仙花占据多数(56.52%),说明自然条件下野生凤仙花仍然普遍存在传粉限制,且10种明确记载为自交不亲和的凤仙花的平均坐果率仅有25.58%,说明凤仙花的自交亲和性对其自然结实具有重要作用(表2)。
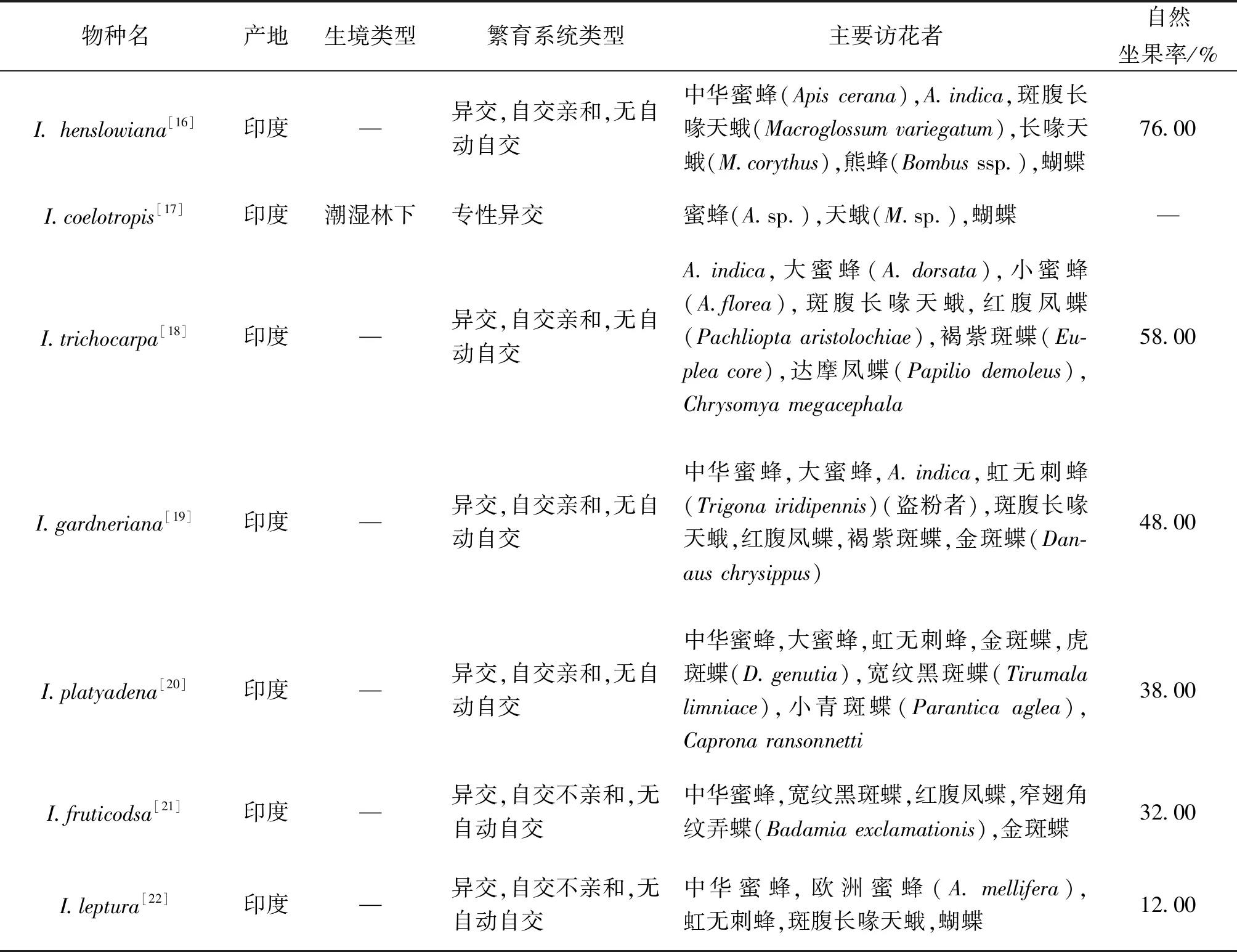
表2 凤仙花属植物的繁育系统比较
3 讨 论
3.1 花部特征和访花昆虫
花部特征包括开花式样和单花特征。菱叶凤仙花成簇丛生以及同株多花的特性可以提高种群对传粉者的整体吸引力。同花梗的两朵单花存在开放时差,这可能是为了避免传粉者的连续访花性,提高异交率。该种花色鲜艳具有花蜜报酬,符合虫媒花特征。三条熊蜂为泛化传粉者,体型较大,而菱叶凤仙花的传粉通道较短(8.41 mm),使得花粉落置于其头部,而同城分布的其他同属植物的花粉一般落置于其背部,这样的位置分化在一定程度上缓解同域物种之间的花粉竞争[38]。该植物单花寿命为5 d左右,花性分配比(雌性期/雄性期)为0.98,高于侧穗凤仙花(I.lateristachys)和匍匐凤仙花(I.reptans)[8],说明菱叶凤仙花的雄性投入低于部分同属植物,授粉程度、传粉者访问频率等均会影响该指标[33]。
菱叶凤仙花主要访花昆虫为三条熊蜂、中华蜜蜂、小豆长喙天蛾、蝴蝶等,盗蜜者为短头熊蜂,存在夜间传粉者。与国内其他凤仙花研究相比,菱叶凤仙花的传粉者种类较为丰富,可能是由于本研究地点位于峨眉山山腰,为低海拔至高海拔的过渡带,传粉者多样性较高。因为不同海拔传粉动物一般不同,比如熊蜂因具有较强的体温调节机制常现于高海拔,而蜜蜂常见于低海拔地区[39]。蜂类访花者访花时间较为固定,8∶00至17∶00为主要访花时段。三条熊蜂的访花停留时间短于中华蜜蜂,可能是由于前者的口器(19.67 mm,往年观测数据)偏长于蜜距(18.14 mm),导致取蜜难度较低,花平台较小难以支撑熊蜂较重的体型,加之菱叶凤仙花的花蜜体积偏低(2.19 μL),所以其停留时间较短。而中华蜜蜂因其口器较短,难以获取花蜜,且大多还存在盗粉行为,所以停留时间较长。小豆长喙天蛾作为外来物种,其出现可能与峨眉山发达的旅游业有关。在印度、湖南等均有天蛾在夜间访花的现象[6,25],而本研究仅在日间观察过天蛾访菱叶凤仙花,但通过夜间开放授粉实验可知菱叶凤仙花也可能存在夜间访花者。蝴蝶常出现在阳光充足的晴天,然而蝴蝶和天蛾常常不被认为是有效传粉者[34]。经前期调查,菱叶凤仙花原生地和引种地的传粉者类群差异不大,但引种地短头熊蜂盗蜜者更多,虫害现象更严重,生存环境更严峻,野生种群的繁育系统待进一步调查,但因本文研究目的重在引种驯化野生菱叶凤仙花,因此对田间种群中的传粉者进行调查对后续研究更具实际意义。
3.2 花粉胚珠比、杂交指数和繁育系统实验
P/O和OCI是测定植物异交率的间接方法。菱叶凤仙花的P/O结果显示该种为专性异交,而OCI和人工授粉实验结果显示,该植物为兼性异交,因此依据P/O值判断为专性异交的结果并不完全准确。不去雄套袋实验显示该植物具有低比率的自动自交能力,这可能是因为菱叶凤仙花雌蕊在花蕾期就已经具有可授性,有自动自交的趋势[30],也可能是因为其他媒介(如蚂蚁)形成的误差。人工授粉并未显著提高自然坐果率,可能与传粉者较丰富有关,比如Sreekala[25]研究发现,夜间传粉者对某些凤仙花自然坐果具有重要作用。菱叶凤仙花鲜艳的花部特征、极高的P/O值表示其为典型的异交植物,但异交受环境的影响极大,其荫蔽的生境和峨眉山多变的天气可能会导致传粉者限制,因此该植物自交亲和。Escaravage等[40]对高山植物传粉研究发现,高山昆虫多样性低,传粉者少,因此很多异交植物有向自交进化的趋势。
3.3 凤仙花属植物繁育系统
植物繁育系统的广义范畴包括生殖、生殖前传粉、传粉者行为以及无性克隆等,对植物繁育系统的了解是认识植物生活史的前提,也是其他相关研究的基本背景知识[3,41]。由于性别系统的不同,植物常常表现出多样化的繁育系统类型。对36种凤仙花属植物繁育系统的分析发现,大部分凤仙花为混合式交配系统,它们大多生活在潮湿荫蔽的环境,各种类之间自然坐果率和传粉者种类的变化较大。凤仙花的混合式交配系统可能与其生境类型有关。在野外荫蔽环境中,大多凤仙花可能会面临传粉限制,因为传粉者更偏爱在阳光充分的环境下访花,蜜蜂活动会受温度调控,其视觉反应也与光敏感度和紫外光反射有关[39,42]。当传粉者稀少时,凤仙花通过高雄性投入提升花部吸引力来吸引传粉者,因此大多野生凤仙花颜色鲜艳、造型奇丽,比如Trunschke等[43]对细距舌唇兰(Platantherametabifolia)林间种群和草地种群的研究发现,林间种群的植物更高、花观赏性更强、传粉限制更严重。自然坐果率较大的变异性可能与时空异质性、传粉者多样性、植物自身交配系统特性等因素相关。
混合式交配系统既是植物在适宜环境下提高后代遗传多样性和适合度的机制,也是恶劣环境中的生殖保障,凤仙花大多为一年生植物,很难形成永久的种子库,须靠异交扩展种群,自交维持种群[30,44]。凤仙花中有不少维持异交的机制,比如雄蕊先熟特性、异交系植株提高开花质量减少开花数量的特性等[45]。异交还可以提高胚珠受精率[46],产生的种子更健康,传播距离更远[45,47]。但如果仅依靠异交,凤仙花很难维系种群。比如本文搜集的10种自交不亲和的凤仙花,其平均坐果率仅为25.58%,这说明自交亲和性对凤仙花自然结实具有重要作用。凤仙花的自交机制包括自动自交、同株异花授粉等。部分凤仙花的雌蕊在花蕾期就有很强可授性,具有自动自交的趋势,虽然该属具有严格的雄蕊先熟异交机制,但同株异花授粉依然会引发自交,维持混合式交配系统[3]。另外,传粉者的泛化性常引起异交结实率的低下,因此自交也是植物应对花粉竞争的重要机制[3]。总之,植物混合式交配系统是动态的,生理机制隐性的自交不亲和机制指出,当异花花粉大量存在时,植物会极大限制同株花粉的生殖率,促进异交,而当传粉资源缺乏时,植物会加大雄性投入或者启动自动自交和营养繁殖作为繁殖补偿机制[3]。因此,混合式交配机制可能是凤仙花植物面对多变的环境影响形成的生殖策略,自交和异交的交配机制是其长期演化形成的有效繁殖机制。
综上,大部分凤仙花属植物为混合式交配系统,以异交为主,自交亲和,与环境影响密切相关,故在该属植物的传粉生态学研究、入侵种防控和新品种选育方面需要考虑其繁育系统的影响。然而本文研究内容仍有不足之处,比如在凤仙花属整体繁育系统的研究中,不同文献中的年份、实验方法和判断标准均可能对结果产生误差。在菱叶凤仙花繁育系统研究中,花粉活力、柱头可授性等指标尚待研究,且目前研究主要针对菱叶凤仙花的人工种群,其野外自然种群的情况还需要进一步调查。
致谢:感谢卢焱茜同学为野外实验工作提供的帮助!
猜你喜欢
中国蜂业(2023年2期)2023-04-04
经济林研究(2022年2期)2022-07-20
今日农业(2021年6期)2021-06-09
基层中医药(2020年10期)2020-11-27
爱你(2019年17期)2019-11-14
爱你·健康读本(2019年5期)2019-06-06
儿童故事画报·发现号趣味百科(2017年10期)2018-03-13
儿童故事画报·发现号趣味百科(2017年9期)2018-03-13
儿童故事画报·发现号趣味百科(2016年4期)2016-06-24
中国民族医药杂志(2016年2期)2016-05-14
