
长期放牧干扰降低了短花针茅荒漠草原植物多样性
2023-09-25 03:42:08赵敏赵坤王赟博殷国梅刘思博闫宝龙孟卫军吕世杰韩国栋
草业学报 2023年9期
赵敏,赵坤,王赟博,殷国梅,刘思博,闫宝龙,3,孟卫军,吕世杰,韩国栋
(1. 内蒙古农业大学草原与资源环境学院,内蒙古 呼和浩特 010018;2. 内蒙古自治区农牧业科学院,内蒙古 呼和浩特 010031;3. 内蒙古民族大学农学院,内蒙古 通辽 028043;4. 鄂尔多斯市农牧业局综合保障中心,内蒙古 鄂尔多斯 017010)
草地覆盖了陆地40%的表面积(除南极洲和格陵兰岛)[1],是具有生态、经济和社会价值的重要自然资源[2]。草地给各种有蹄类家畜提供各种各样美味的食物、给人类提供各种户外活动和多彩的自然风光,同时也是生物多样性重要的保护内容[3-5]。草地植物多样性受环境影响,具有尺度依赖性[6]。在全球及区域尺度上,地带性气候决定了植物多样性的分布格局[7],在景观及更小尺度上,非地带性因素是影响草地植物多样性的主导因素[8]。放牧作为一种普遍的草地利用方式,本质上是各种有蹄类家畜通过选择性采食植物地上器官(主要是叶片)、践踏、排便和排尿改变着草地植物物种组成、土壤的理化性质和土壤微生物活性等。大量的研究表明,过度放牧是草地植物多样性下降的主要原因[9-13]。
草地植物多样性对放牧的响应在过去几十年已被广泛研究,提出了几个流行的假说,例如,“适度干扰假说”认为,适度放牧能够维持最高的生物多样性[14]。放牧史说认为,放牧草地的植物多样性不仅取决于放牧强度,还取决于放牧周期,长期放牧会提高草地群落结构的稳定性[15]。目前,很多研究表明,禁牧是提高植物多样性最简单、经济、高效的措施[16-20]。但是,也有研究表明,随着放牧强度的减弱,物种多样性并没有显著改变。甚至有研究报道,高放牧强度对植物多样性有积极影响[21-22]。植物多样性对不同放牧强度响应的差异可能是研究的草地类型、放牧家畜种类和数量、恢复时间的差异造成的。其次,放牧强度严格来说属于定性指标,多样性的测度指标也很多,对放牧的敏感性有较大差异[23]。因此,进一步明确采取何种恢复草地植物多样性的方式仍然是当前研究的课题。
约占内蒙古草地总面积10.7%的内蒙古荒漠草原是亚洲中部特有的一种最干旱的草地类型[24],由于其植被稀少,利用形式单一,草地生态系统极易受外界环境影响[25]。近些年来,牧民为追求更高的经济效益,不断增加牲畜存栏数量,造成草地植物多样性锐减,退化严重[24-25]。Huston[26]认为,局地草地生态系统经常只有少部分的物种,而适度放牧却导致了更多物种的共存。过度放牧导致适口性好的物种被过度消耗,降低了其繁殖能力[27-28],只有更耐牧或避牧的物种能够生存,致使物种丰富度衰退[29]。高强度的放牧会增加植物的死亡率,降低植物多样性[30]。长期禁牧,凋落物富集,密集型丛生禾草对其他物种的强烈竞争,冠层扩张,可利用间隙的占领,使得其他物种很难定居和繁殖,不利于植物多样性的提高[31]。相反,适度放牧使栖息地更异质化,提供了更多的空隙给其他物种定居[32-33]。本研究探讨长期围封下不同放牧强度(重度放牧、中度放牧、轻度放牧和禁牧)植物多样性的恢复状况。假设:1)不同放牧强度下植物多样性是有差异的,适度放牧下的植物多样性最高;2)重度放牧下草地群落中的优势物种,在中度放牧、轻度放牧和禁牧草地群落中都会出现,并且具有较高的优势度。目的:1)明确长期不同放牧强度条件各个群落物种组成和功能群组成;2)明确长期不同放牧强度条件下植物多样性变化。
1 材料与方法
1.1 研究区概况
研究区位于内蒙古自治区农牧业科学院综合试验示范中心四子王基地(41°47′17″ N,111°53′46″ E,海拔1450 m)。该区属于典型中温大陆性气候,春季干旱多风,夏季炎热,冬季寒冷而多风。年均降水量为220 mm,主要集中在5-8 月,年均气温3.8 ℃,湿润度0.15~0.30,无霜期175 d。年蒸发量2300 mm,年均风速4~5 m·s-1,年日照时数3117.7 h,土壤类型为淡栗钙土。植被以强旱生禾草为主,属荒漠草原地带性植被,建群种是短花针茅(Stipa breviflora),优势种为无芒隐子草(Cleistogenes songorica)和冷蒿(Artemisia frigida),非优势种大约有20多种。
1.2 试验设计与取样方法
试验采用随机区组设计,2004 年将地势平坦,植被均匀的50 hm2天然草地进行围封并分为12 个小区,各个小区面积基本一致,为4.4 hm2,试验将12 个小区划分为3 个区组,每个区组4 个处理,分别为重度放牧(heavy grazing,HG)、中度放牧(moderate grazing,MG)、轻度放牧(light grazing,LG)和禁牧(no grazing,CK),用围栏隔开,对应载畜率为2.71、1.82、0.93、0 sheep·hm-2·a-1(图1)。试验选用当地成年2 岁羯羊,每年6-11 月为放牧期,每天早上6:00 放牧,晚上6:00 归牧。

图1 试验设计Fig.1 Experimental design
2017 年8 月中旬(生长高峰期)进行取样,在每个小区随机调查6 个1 m×1 m 样方(每个样方距离围栏5 m 以上,避免边际效应)。调查指标有各物种密度、频度、盖度,凋落物生物量等,共72 个样方。
1.3 功能群划分
植物生活型是植物对综合生境条件长期适应而在外貌上反映出来的植物类型,而群落的生活型功能群组成则是环境因子的综合反映。本研究按生活型划分为4 类功能群:多年生禾草(perennial grasses,PG);灌木、半灌木(shrub and semi-shrubs,SS);多年生杂类草(perennial forbs,PF);一、二年生植物(annual-biennial plants,AB)。
1.4 数据处理
1.4.1 重要值(important value,IV) 在草地生态系统中,植物重要值是物种或功能群在群落中的相对优势程度,能够反映出物种或功能群应对干扰所作出的变化响应。本研究采用经典的Curtis 计算公式,如下:
式中:RD(relative density)表示相对密度(某个物种的个体数与所有物种个体数的比值,%);RC(relative coverage)表示相对盖度(某个物种的盖度与所有物种盖度的比值,%);RF(relative frequency)表示相对频度(某个物种的频度与所有物种频度的比值,%)。
1.4.2 植物多样性 草地植物多样性是群落水平的概念,它能反映应对干扰后,物种丰富程度、均匀程度以及优势度的变化,通常是用群落中的种群数量特征以及某个种群的个体数量与所有种的个体数量的相对比值来计算。本研究采用α 多样性指数,公式如下:
Patrick 丰富度指数(Patrick richness index,R):
Shannon-Wiener 多样性指数(Shannon-Wiener diversity index,H):
Simpson 优势度指数(Simpson dominance index,D):
Pielou 均匀度指数(Pielou evenness index,J):
式中:S代表物种数;N代表所有物种的多度;Pi=RD,表示某个物种的相对密度。
1.5 统计分析
不同放牧强度之间的所有植物、优势与非优势植物和生活型功能群的重要值以及植物多样性指数的差异显著性均采用单因素方差分析进行检验,显著性水平设为P<0.05。并用LSD 检验进行平均值之间的比较,显著性水平设为P<0.05。通过线性模型分析群落多样性和优势植物及物种、生活型功能群的动态变化,分析优势植物及物种、功能群和物种多样性之间的相关性(Pearson 相关系数)。相关数据制表、结果绘图在Excel 2010 和SigmaPlot 12.5 软件中操作,运用SPSS 19.0 软件进行分析。
2 结果与分析
2.1 不同放牧强度下植物组成和功能群重要值变化
2017 年8 月生长高峰期短花针茅荒漠草原调查结果可知(表1),样地植物由9 科,17 属,21 个物种组成。其中禁牧区包含9 科,17 属,20 个物种;轻度放牧区包含7 科,13 属,15 个物种;中度放牧区包含7 科,9 属,10 个物种;重度放牧区包括4 科,7 属,7 个物种。

表1 植物物种重要值在不同放牧强度下的差异Table 1 Differences in important values of plant species under different grazing intensities
对植物物种的重要值在不同放牧强度下的差异进行分析,冷蒿、短花针茅、无芒隐子草、银灰旋花是该区域的优势种。这4 种植物在不同的放牧强度下,其重要值较其他物种高,拥有绝对优势(表1)。但在不同放牧强度下优势种和非优势种的重要值有显著差异(图2)。随着放牧强度的增加,优势种的重要值逐渐增加,并且禁牧和轻度放牧显著低于中度放牧和重度放牧(P<0.05,图2A)。同时,优势种之间也有显著差异(表1)。短花针茅的重要值是重度放牧>中度放牧>轻度放牧>禁牧,并且重度放牧和中度放牧显著高于轻度放牧和禁牧(P<0.05);冷蒿的重要值是禁牧>轻度放牧>中度放牧>重度放牧,并且禁牧显著高于中度放牧和重度放牧(P<0.05),同时轻度放牧显著高于重度放牧(P<0.05);无芒隐子草的重要值是中度放牧>重度放牧>轻度放牧>禁牧,并且中度放牧显著高于禁牧(P<0.05);银灰旋花的重要值在各个放牧强度下无显著性差异(P>0.05)。相反,非优势种的重要值随着放牧强度的增加而降低,并且禁牧和轻度放牧显著高于中度放牧和重度放牧(P<0.05,图2B)。

图2 不同放牧强度下优势和非优势植物种的重要值Fig.2 Important value of dominant and non-dominant plant species in different grazing intensities
对群落植物功能群重要值在不同放牧强度下的差异进行分析(图3),在不同放牧强度长期处理下,多年生禾草随着放牧强度的增加,重要值增加,并且禁牧和轻度放牧显著低于中度放牧和重度放牧(图3A);多年生杂类草的重要值随着放牧强度的增加逐渐下降,禁牧显著高于中度放牧和重度放牧(图3B);灌木和半灌木随着放牧强度的增加,重要值逐渐减小,并且禁牧显著高于中度放牧和重度放牧,同时轻度放牧和中度放牧显著高于重度放牧(图3C);一、二年生植物的重要值在轻度放牧下最高,并且,轻度放牧和禁牧显著高于中度放牧和重度放牧(图3D)。

图3 不同放牧强度下植物功能群重要值Fig.3 Important value of plant functional groups in different grazing intensities
2.2 不同放牧强度下植物多样性差异
不同放牧强度下群落的植物多样性的研究结果显示,群落物种组成结构发生了显著变化(图4)。随着放牧强度的增加,Patrick 丰富度指数逐渐下降,禁牧显著高于轻度放牧、中度放牧和重度放牧,并且轻度放牧显著高于中度放牧和重度放牧(图4A);Shannon-Wiener 多样性指数随着放牧强度增加呈递减趋势,并且禁牧显著高于中度放牧和重度放牧(图4B);Pielou 均匀度指数在中度放牧下最高,并且显著高于重度放牧(图4C);Simpson 优势度指数随着放牧强度增加呈递减趋势,禁牧和轻度放牧显著高于重度放牧(图4D)。

图4 不同放牧强度下植物群落α 多样性指数Fig.4 α diversity index of plant community in different grazing intensities
2.3 群落组成物种、功能群和物种多样性的关系
对群落组成物种和植物多样性指数进行相关性分析发现:Patrick 丰富度指数与优势植物重要值呈极显著负相关(表2),说明优势植物重要值越大,群落物种种类越少,具体表现为冷蒿和银灰旋花的重要值与Patrick 丰富度指数呈极显著正相关,而短花针茅和无芒隐子草的重要值与Patrick 丰富度指数呈极显著负相关,说明冷蒿和银灰旋花的重要值越大,群落物种越丰富;相反,当短花针茅和无芒隐子草重要值增大时,群落的物种丰富度降低;群落的Shannon-Wiener 多样性指数与Patrick 丰富度指数的规律基本一致,只是与无芒隐子草相关性并不显著,但和短花针茅一样都表现为消极作用;Simpson 优势度指数和冷蒿的重要值呈极显著正相关,与短花针茅的重要值呈极显著负相关。优势植物短花针茅、冷蒿、无芒隐子草、银灰旋花的重要值与α 多样性指数相关性分析表明,短花针茅和冷蒿呈此消彼长的作用,而银灰旋花和无芒隐子草起过渡作用。

表2 优势植物重要值与植物物种α 多样性的相关关系Table 2 Correlation between important values of dominant plants and α diversity of plant species
植物功能群的重要值与物种多样性相关分析显示:多年生禾草的重要值和Patrick 丰富度指数 、Shannon-Wiener 多样性指数、Simpson 优势度指数呈极显著负相关,说明当禾本科植物重要值增大,群落物种组成更单一;相反,多年生杂类草、灌木、半灌木和一、二年生植物的重要值和Patrick 丰富度指数、Shannon-Wiener 多样性指数、Simpson 优势度指数呈显著或极显著正相关(表3)。Pielou 均匀度指数和优势物种、功能群的重要值相关性均未达到显著水平(表2 和表3),可能是由于家畜采食具有随机性。
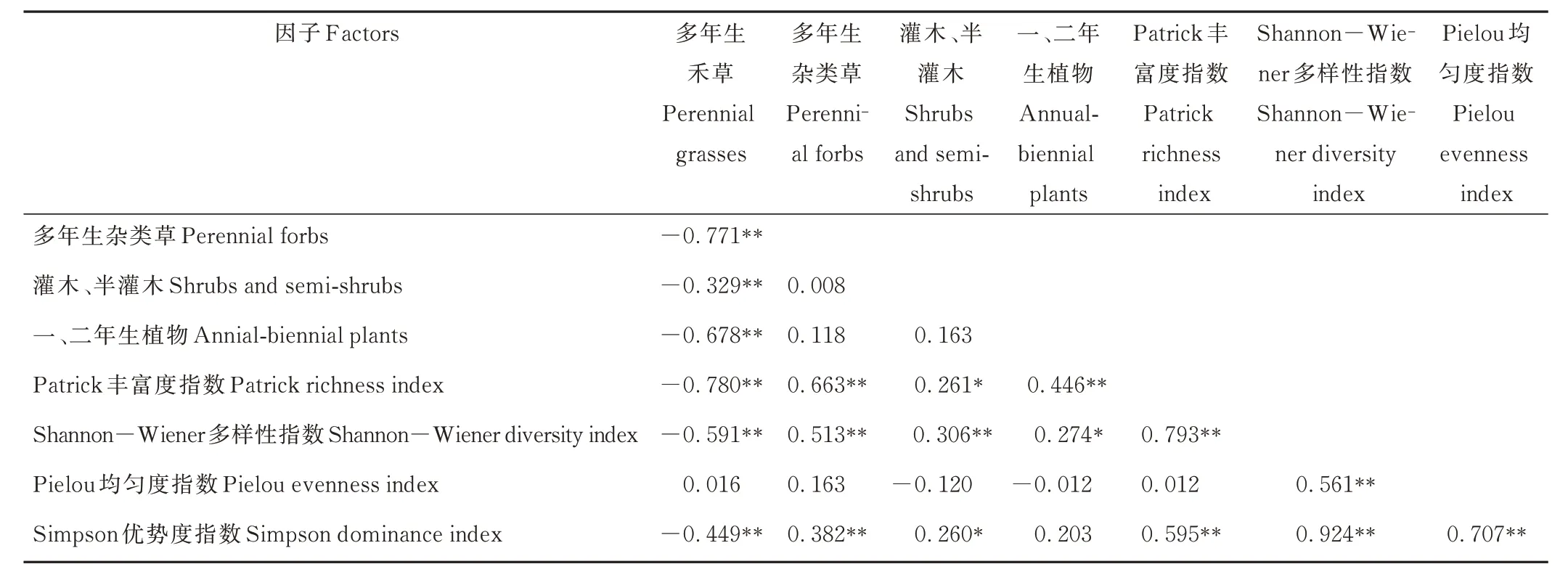
表3 植物功能群重要值与物种多样性的相关关系Table 3 Correlation between important values of plant functional groups and species diversity
3 讨论
3.1 放牧对群落物种组成的影响
草地群落中往往只有几个有竞争力的物种长期共存[26]。放牧改变了群落物种组成和物种相对优势度/重要值[33]。在本研究中,优势物种短花针茅、冷蒿、无芒隐子草维持着稳定的群落结构。随着放牧强度的增加,优势种的重要值增大,非优势种的重要值下降。首先,这几种优势植物都是家畜喜食的物种,但是当它们共存时,家畜就有选择性采食的行为[34-35],其次,从生长和生殖角度,它们应对放牧干扰的响应不同[32]。本研究表明在生长高峰期8 月,家畜优势植物采食率为冷蒿>无芒隐子草>短花针茅;冷蒿属于冷季植物,短花针茅和无芒隐子草属于暖季植物。有研究表明暖季植物往往比冷季植物生长滞后[36],可能在生长高峰期前,家畜主要采食的对象是冷蒿,因此,高峰期短花针茅和无芒隐子草的优势度变大;也有研究表明,植物生殖分配对放牧干扰非常敏感,草原植物在面对放牧干扰时会权衡它们的繁殖策略[37],大量研究表明,随着放牧强度增加,植物有性生殖能力减弱[37-40],随后主要以营养生殖为主[41],杜利霞等[40]的研究表明,冷蒿有性生殖分配减少,生殖构件显著下降,生殖枝数和种子产量降低,甚至在重度放牧中有性生殖几乎消失;白永飞等[42]的研究表明,放牧通过家畜践踏使短花针茅株丛破碎化,随着放牧强度增大,大株丛形成若干小株丛,后脱离母株通过“分丛”进行营养繁殖,使种群密度增大。王亚婷[43]指出,随着放牧强度增大,短花针茅叶片硬度增大,降低了适口性,可能这也是抵抗家畜采食的一种生存方式。家畜长期对冷蒿的偏食性,使得被偏食的植物休憩补偿时间不足,因此不利于其繁殖,相反给短花针茅和无芒隐子草提供了更多的生态位,也是放牧强度增加,短花针茅和无芒隐子草重要值增加的原因[43],虽然随着放牧强度的增加,优势植物冷蒿、无芒隐子草和短花针茅都表现出避牧的行为[33],植株矮小化和生殖策略转变,但是家畜长期选择性采食可能是优势物种重要值变化的根本原因,其次是优势物种的形态学特征,导致了应对家畜践踏的承受程度不同,短花针茅呈现密集型丛生生长,无芒隐子草呈现疏丛生长,丛生生长的植物基部表面积较大,抗践踏能力强,储存碳水化合物的能力较强,当被采食后,更容易再生。冷蒿虽然匍匐生长,当构件被践踏破坏后,残枝可以营养繁殖,但是相比于短花针茅和无芒隐子草,需要的资源更多。
3.2 放牧对群落功能群的影响
在干旱半干旱草地,植物生长高峰期,随着长期放牧强度的增加,多年生禾草的优势度增加,灌木、半灌木和多年生杂类草优势度降低[36,44],本研究结果与其一致。原因是长期放牧,群落组成与结构简单化,家畜的选择性采食使多年生禾草逐渐占优势,此外,长期放牧的累积效应,使得群落发生演替,多年生禾草替代了杂类草的生态位[45]。而备受争议的是短命的一、二年生植物,随着放牧强度增加,一、二年生植物优势度增加[33]、减小[36]、无变化[44]。本研究结果与Zhang 等[36]的一致。增加的原因可能是一、二年生植物繁殖速度快,更容易占据由于放牧践踏、粪尿造成的小斑块和草丛空隙[33];在本研究中发现轻度放牧一、二年生植物优势度最高,原因可能与其一致,但是长期放牧,中度和重度放牧区一、二年生植物急剧下降,可能是被家畜大量采食所引起。同时一、二年生植物生长繁殖很大程度上依赖于水分的调节[36],在干旱半干旱草地,降水往往波动较大,很有可能在正常的生长高峰期,出现干旱化,导致一、二年生植物无法生长。
3.3 长期放牧对植物多样性的影响
很多研究表明适度放牧干扰会增加群落植物多样性,验证了中度干扰假说,但是局限于短期放牧干扰[33]。在本研究初期,适度放牧确实使植物多样性有所提高[46-47],那是因为适口性好的优势植物冷蒿足以维持家畜的采食,同时,家畜分泌的唾液也会刺激被采食植物的再生长[33],因此适度放牧给多年生杂类草和一、二年生植物提供了更多的生存空间,植物多样性提高。但是随着放牧周期增加,Zhang 等[36]、韩梦琪[44]的研究结果均显示,长期放牧导致植物多样性线性降低,验证了Milchunas 等[48]的观点,即半干旱草地植物多样性与放牧的响应呈负相关,由此看来,植物多样性对长期放牧的响应主要取决于资源的可利用性[49]。从群落的物种组成来看,本研究发现,多年生禾草和多年生杂类草的相对优势度在不同的放牧强度处理下占绝对优势,随着放牧强度增加,多年生禾草的相对优势度增加,多年生杂类草的相对优势度降低,植物丰富度指数、多样性指数和优势度指数在下降。而多年生禾草和多年生杂类草中主要植物就是短花针茅、无芒隐子草和冷蒿,因此优势植物之间的相对优势决定了植物多样性的高低。
4 结论
随着放牧强度的增加,短花针茅荒漠草原植物生长高峰期长期禁牧、轻度放牧、中度放牧和重度放牧植物多样性呈下降的变化趋势。植物多样性的高低和优势植物的相对优势度密切相关。禁牧是恢复退化的荒漠草原植物多样性的理想途径之一。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
草业科学(2021年9期)2021-11-03 02:42:16
食品安全导刊(2021年20期)2021-08-30 06:40:50
草业科学(2021年6期)2021-07-28 07:19:50
草地学报(2021年6期)2021-07-05 03:50:08
畜牧与饲料科学(2021年3期)2021-06-23 09:38:20
电脑报(2020年12期)2020-06-30 19:56:42
癌症进展(2016年9期)2016-08-22 11:33:10
水生生物学报(2015年1期)2015-02-28 16:01:05
声屏世界(2014年6期)2014-02-28 15:18:07
