
三江源地区人工克隆植物群落生物多样性对初级生产力的影响及机制
2023-09-25 03:42:08刘增辉卢素锦王雨欣张春辉尹鑫
草业学报 2023年9期
刘增辉,卢素锦,王雨欣,张春辉,尹鑫
(1. 青海大学生态环境工程学院,青海 西宁 810016;2. 青海大学省部共建三江源生态与高原农牧业国家重点实验室,青海 西宁 810016)
近几十年来,生物多样性丧失越来越严重,由此带来的生态系统功能和服务能力的变化一直是生态学家们关注的热点[1-3],其中最基本的生态系统功能生产力被关注的最多[4-5]。最早的时候,生态学家认为生物多样性和生产力之间是一种单峰关系[6],但是这种关系很快受到了质疑[7]。随着研究的推进,越来越多其他的模式被发现,例如,生物多样性与生产力之间的正相关关系[8-9]、负相关关系[10]、无显著关系[11]、U 型关系[12]等。虽然有多种因素会引起这种关系的变化(例如,空间范围[13-14]、营养网络[15]、气候变暖和大气氮沉降[16]等),但是这些影响因素都容易在试验中操作。有一种因素最不易与物种丰富度区分,也最难被研究,那就是蕴含在物种组成中的物种特性。在一些研究中,物种丰富度很小的改变可能伴随着物种组成变化的两个极端(巨大变化或者几乎没有变化)[17-18]。物种特性能够决定物种的种内和种间相互作用强度,这能够改变多样性-生产力关系[19],甚至能够越过生物多样性直接主导群落的生产力[20]。因此,有一些研究开始关注关键物种、优势物种、稀有物种等在这个关系中发挥的重要作用[20-22],以及在这些物种构建的群落中生物多样性-生产力关系呈现的状态[9]。例如,如果群落包含具备功能性优势和竞争性优势的物种,那么这个群落很有可能呈现的是负的多样性-生态系统功能关系[23]。此外,一些关键物种的冗余功能可能补偿生物多样性丢失带来的生态系统功能的减少,使得多样性与生态系统功能之间并未呈现任何关系[24]。对于稀少物种,有些研究漠视它的存在,选择直接在数据分析中移除[25],虽然它在群落功能中发挥着重要作用[26]。然而,以往的研究极少关注另外一类特殊的物种,这些物种天然就具备快速占据地上地下空间以及快速获取资源的能力,这就是能够进行克隆繁殖的植物。这类植物在不同生态系统中占据不同的地位[27]。这类植物中,有禾本科等潜在优势物种,有鹅绒委陵菜(Potentilla anserine)等群落构建先锋物种,也有豆科等固氮植物。无性繁殖能力使这些物种的种群扩张摆脱了种子库的限制,进而能够使得这些物种更加适应恶劣的环境[28],并在群落中占据更大的比例[29],并且这类植物能够更快更强地在群落中发挥自己的生态功能,这是单纯由种子繁殖的植物在相同时间内无法做到的。然而这类植物构建的群体中生物多样性-生态系统功能关系如何变化,现有的研究却非常稀少。
当前的研究认为生物多样性效应的出现主要依赖于两种机制:互补效应和选择效应[30]。Fox[31]对该机制进行了进一步的拓展,把选择效应划分为特征依赖型互补效应和优势度效应,并把原先的互补效应更名为非特征依赖性互补效应,由此两种机制转变成为3 种。非特征依赖性互补效应能够发挥作用主要是因为助长作用或者物种间更充分、互补地利用各种资源,这两种因素都能够促使物种更多的群落产生更高的生产力。其次,选择效应能够发挥作用是因为物种越多的群落包含高产物种的可能性越大,群落有更高生产力的可能性也就越大[32]。如果这个高产物种生物量的增加并不是以其他低产物种生物量降低为代价,那么特征依赖型互补效应就发挥了作用;如果是以其他低产物种生物量降低为代价,那么优势度效应就发挥了作用。对克隆植物来说,在合适的条件下植物以母株为中心向四周扩散[33],其相连的分株之间可以转移并共享获得的资源(例如光合产物、水分等)[34],这种特殊的空间繁殖策略不仅有利于其适应当地恶劣的环境,而且还可以改善周围生境并有助于其他植物的生存和繁殖[28],即它对其他物种的生存存在助长作用。此外,它在生长季的任何时候都可以通过无性繁殖扩大自己的种群,再加上不同克隆植物占据的生态位不完全相同,这些特性可以促使这些植物更加充分地占据空余生态位。不管是对其他植物的助长作用还是群落生态位得到更充分地互补性利用都能够增强非特征依赖性互补效应来促进多样性-生产力关系的正线性相关关系。其次,资源的多寡也可能影响植物的无性繁殖个体数,进而改变群落的丰度。丰度的改变可能使物种之间的相互作用方式由助长向竞争转变[35],或者发生竞争强度的改变[36]。这些变化能够促使特征依赖型互补效应或优势度效应发挥作用,进而改变生物多样性-生产力关系强度甚至方向。尽管克隆植物可能造成多样性-生产力关系的改变,但是作用机制是上述3 种的哪一种,目前尚不清楚。
虽然上述3 种机制的强弱可以通过数值展现出来,但是挖掘这些机制变化背后的原因却需要对群落中植物性状特征进行检测[37-38]。植物性状差异可以反映不同物种之间资源利用的差异[39],且在草原生态系统中对植物生长限制最多的是氮元素和光资源[40]。例如,植物对光的截获能力与其高度(height)、比叶面积(specific leaf area, SLA)、叶片氮磷含量密切相关[41]。越高的植物截取的光资源越多,光竞争能力越强,越能够压制或者竞争排除比它低的植物,导致特征依赖型互补效应或者优势度效应增大[42]。当植物有较高的SLA 和叶片氮磷含量时,它们即使在弱光环境中也能获得足够的光资源[43],这能够促进群落光资源的互补性利用,进而带来非特征依赖性互补效应的增强。与之类似,不同物种的高度差异越大,它们对垂直空间的利用越充分[44];同等高度物种的植株个体较多,它们对水平空间的利用越充分。这些导致的一个结果就是整个群落的空间利用越完全,光资源的获取也越多,也能够带来非特征依赖性互补效应的增强。基于此,本研究关注了植物特征的变化,以此了解3 种机制背后的变化机理。
在青藏高原高寒草地生态系统中,克隆植物物种数量高于非克隆植物,占植物总种数的53.09%,藏嵩草(Kobresia tibetica)草甸和沼泽化帕米尔薹草(Carex pamirensis)草甸克隆植物占比更是达到了67%和63%。另外,高寒草地克隆植物的相对重要值也显著高于非克隆植物[29]。为了更好地了解克隆物种间的相互作用方式,为退化草地生态系统的修复提供可行性建议,本研究在青藏高原腹地“三江源”区选取样地,选择3 种克隆植物[垂穗披碱草(Elymus nutans)、花苜蓿(Medicago ruthenica)和鹅绒委陵菜]构建多样性梯度,以此探究克隆植物多样性和物种组成与生产力的关系及相关机制。
1 材料与方法
1.1 研究地点和数据收集
本研究的试验样地位于青海省果洛州玛沁县大武滩(海拔3958 m,100°14′ E,34°29′ N),地处素有“中华水塔”之称的青藏高原腹地“三江源”区。属于大陆高寒气候,雨热同季,雨量稳定,无四季之分,仅有冷暖季之别,冷季漫长,干燥而寒冷,暖季短暂,湿润而凉爽; 温度年差较小而日差较大,年平均气温为-0.2 ℃,1 月(最冷月)平均气温-12.5 ℃,7 月(最热月)平均气温10.1 ℃;夏秋季短,雨量集中,年降水量为420~560 mm,主要集中在5-9 月;牧草年平均生长期150 d,地上生物量从5 月开始增加,7 月底至8 月初达到最大,10 月植物开始衰老。建群种主要为矮蒿草(Kobresia humilis),主要伴生种为高山嵩草(Kobresia pygmaea)、垂穗披碱草、早熟禾(Poa annua)、细叶亚菊(Ajania tenuifolia)等。主要土壤类型为高寒草甸土,年平均无霜期26 d,年平均日照时数2500 h。
本研究选取了三江源地区常见的3 种克隆植物:禾本科垂穗披碱草(En)、豆科花苜蓿(Mr)和杂类草鹅绒委陵菜(Pa),这些植物在高度和叶形上具有不同的表型特征,垂穗披碱草高度(800~1200 mm)占据绝对优势且叶子稀疏扁平[(60~180) mm×(3~5) mm],能够通过分蘖进行无性繁殖;鹅绒委陵菜高度(150~250 mm)相对较低但叶子浓密(11~25 对)且呈椭圆形[(10~25) mm×(5~10) mm],能够通过块状根和匍匐茎进行无性繁殖;花苜蓿高度(200~700 mm)适中且叶子呈羽状三出复叶[(6.0~25.0) mm×(1.5~12.0) mm],能够通过根部的不定芽进行无性繁殖。
本研究构建了6 种不同的物种组合:花苜蓿单播(Mr)、鹅绒委陵菜单播(Pa)、垂穗披碱草单播(En)、花苜蓿与垂穗披碱草混播(En+Mr)、鹅绒委陵菜与垂穗披碱草混播(En+Pa)、3 个物种的混播(En+Pa+Mr),每种组合设置3 个重复,总共有18 个2 m×3 m 的小区。然后将不同物种组合随机分配到小区中,并且用0.5 m 的缓冲带隔离。在每年8 月下旬,在每个小区内设置50 cm×50 cm 的样方,测定物种的盖度、多度和植株个数(母株与子株数量之和,禾本科植物为母体与分蘖子体之和),在样方内齐地面剪切植物样品。将植物样品按物种分类,在80 ℃干燥48 h 至恒重,称重得到物种生物量数据用于后续分析,样方内所有物种地上生物量之和表征群落生产力(g·m-2)。
在两个生长季内,采集样方植物样品的同时,也对与植物生长和资源利用策略相关的植物功能性状进行测量。在样方内分别统计各物种的株高(height)和植株个数(individuals,植株包含En 的分蘖数、Mr 的母株与子株、Pa 的母株与子株)。此外,在每个小区样方外的区域随机挑选30 株植株,采集足够的叶片,用于测定比叶面积(SLA)和叶片氮含量(leaf nitrogen content,LNC)。在每个试验小区,分物种计算各样方植株个数,测定样方中每个物种最高的3 株植株的高度,利用叶面积仪(LI-COR,Lincoln,美国)测定鲜叶的叶面积,然后将鲜叶样品放入烘箱内(80 ℃,48 h)烘干至恒重,称重得到各物种叶干重,以此计算各物种比叶面积(叶面积与叶干质量之比)。研磨烘干后的叶片,取0.30~0.50 g 过0.25 mm 筛的植物样品,用H2SO4-混合加速剂-蒸馏法[45]得到植物叶片氮含量消煮液,最后用Skalar San++(SA9101D,荷兰)测定叶片氮含量。
2016 年之前研究样地一直用于牦牛和藏羊的冬季放牧(11 月30 日-1 月15 日),这是该地区常见的冬季放牧,放牧发生在牧草休眠后并且清除所有现存凋落物。2017 和2018 年对样地进行5 次犁翻,清除样地现有植被并且尽量减少土壤种子库。2019 年6-7 月,3 个物种(2018 年在该样地种群获取)以每个样地2400 株的总密度进行播种,随后测量种子发芽率,以确保每个群落中的幼苗比例相同。在生长季,定期进行人工清除杂草。数据收集时间为2020-2021 年。
1.2 数据计算
用净多样性效应(net biodiversity effects,NE)来评估混播是否增产,NE 可在混播中观察生物量和预期生物量之间的差异[32],也是3 个附加成分的总和,计算公式为:
TICE 为非特征依赖性互补效应(trait-independent complementary effect),计算公式为:
TDCE 为特征依赖型互补效应(trait-dependent complementary effect),计算公式为:
DE 为优势度效应(dominance effect),计算公式为:
式中:DE为正时,单播中高产的物种在混播中的相对产量增加,但是以牺牲单播中低产的物种在混播中的相对产量为代价;DE为负时,单播中低产物种在混播中的相对产量增加,但是以单播高产物种的相对产量降低为代价。
本研究利用单播和混播生物量的自然对数响应比(natural logarithm response ratio,LNRR)估算某个物种的超产现象,以此来评估该物种承受竞争强度的强弱,计算公式为:
式中:Oi是混播中物种i的生物量,Mi是单播中物种i的生物量,Pi是种子中物种i的初始比例。当LNRR>0 时,与单播相比,一个物种在混播中产量过高(该物种的种间竞争强度小于种内竞争强度);当LNRR<0 时,该物种在混播中产量不足(该物种的种间竞争强度大于种内竞争强度)。
1.3 数据处理
本研究数据计算均为3 个重复的平均值±标准误。使用带交互作用的双因素方差分析方法(Two-way ANOVA)评估了物种组成和年份对地上生物量、NE、TICE、TDCE、DE 和各功能性状的影响,接着评估了物种丰富度和年份对以上变量的影响,最后利用LSD 法对以上结果进行多重比较;然后,用t检验评估了各混播的NE、TICE、TDCE、DE 以及各物种的LNRR 值是否与零存在显著差异。方差分析、t检验和作图都在R 4.1.0 软件中进行[46],方差分析结果的多重比较使用“agricolae”软件包来完成,作图使用“ggplot 2”软件包来完成。
2 结果与分析
2.1 三江源地区克隆植物群落物种丰富度和物种组成对地上生物量的影响
在试验的第一年,随着物种丰富度的增加,地上生物量呈先上升后下降的趋势(P<0.05, 图1a),而不同物种组成对地上生物量的影响并不显著(P>0.05, 图1b)。在第二年,3 个物种混播的地上生物量提升较快,与两物种混播之间无显著差异(P>0.05, 图1a);花苜蓿(Mr)和鹅绒委陵菜(Pa)单播的生物量显著低于其他几种物种组成(P<0.05, 图1b)。在播种第二年两物种混播和3 个物种混播的生物量显著增加(P<0.05),且两者之间的地上生物量无显著差异(P>0.05, 图1b)。

图1 物种丰富度和物种组成对克隆植物群落生物量的影响Fig.1 The effects of species composition and species richness on aboveground biomass in clonal plant community
2.2 物种组成和物种丰富度对NE、TICE、TDCE、DE 和LNRR 的影响
研究结果表明:在第一年,不管是物种丰富度还是物种组成在净多样性效应(NE)上都没有显著差异(P>0.05, 表1 和表2),但是两物种混播的NE 显著大于零(En+Pa: t=4.22,P<0.05; En+Mr: t=12.93,P<0.01; 图2a),而3 个物种混播的NE 并没有显著大于零(En+Mr+Pa: t=2.10,P>0.05; 图2b);在研究的第二年,3 个物种混播的NE 有较快增长,虽然在物种组成上2 个两物种混播和3 个物种混播之间没有显著差异(P>0.05, 图2a),但在物种丰富度上3 个物种混播的NE 显著大于两物种混播(P<0.05; 图2b)。不论是物种丰富度还是物种组成都不存在非特征依赖性互补效应(TICE)上的显著差异,并且除了试验第一年的En+Pa 混播组合(2020: t=2.27,P=0.08),其余所有物种丰富度和物种组成的TICE 都显著大于零(P<0.05, 图2c, d)。不同的物种组成和物种丰富度之间特征依赖型互补效应(TDCE)都不存在显著差异(P>0.05, 图2e, f),而在第二年的En+Pa 混播和物种丰富度为2 时TDCE 显著大于零(t=3.11,P=0.04; t=3.19,P=0.012)。优势度效应(DE)在不同的物种组成和物种丰富度之间不存在显著差异,并且也没有显著大于或小于零(图2g, h)。另外,年份对NE、TICE、TDCE 和DE 的影响都是显著的(P<0.05, 表1 和表2),且在试验的第二年这些指标均显著增大(P<0.05, 图2)。
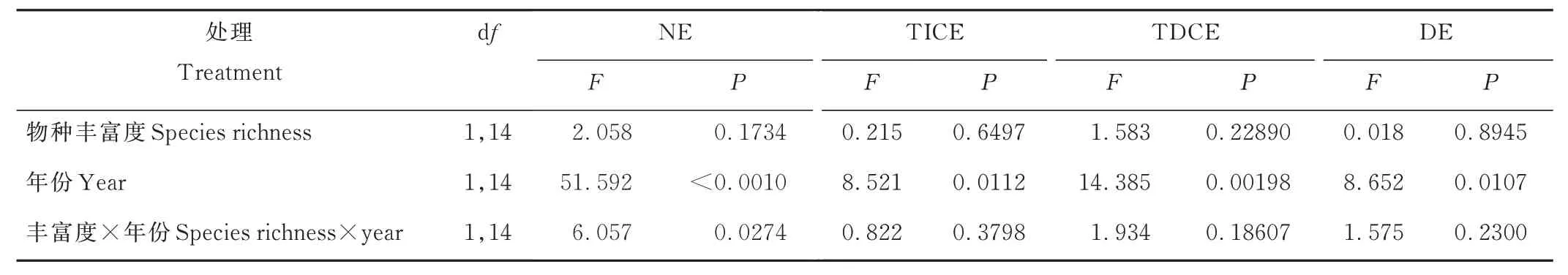
表1 物种丰富度和年份对NE、TICE、TDCE、DE 的影响Table 1 The effects of species richness and year on NE, TICE, TDCE and DE

表2 物种组成和年份对NE、TICE、TDCE、DE 的影响Table 2 The effects of species composition and year on NE, TICE, TDCE and DE

图2 物种丰富度或物种组成在不同年份内分别对净多样性效应、非特征依赖性互补效应、特征依赖型互补效应和优势度效应的影响Fig. 2 The effects of species richness or species composition on net biodiversity effects (NE), trait-independent complementary effects (TICE), trait-dependent complementary effects (TDCE) and dominance effects (DE) in different years
对自然对数响应比(LNRR)的结果分析发现,所有混播中垂穗披碱草(En)的LNRR 值都为正,且在2020 年的En+Mr 混播以及2021 年的3 个物种混播和En+Mr 混播中,En 的LNRR 值都显著大于零(P<0.05, 图3),表明与单播相比,混播中En 的表现更好。与之相反,Pa 的LNRR 值都为负,且在试验第二年的3 个物种混播中,其LNRR 显著小于零(P<0.05, 图3),表明混播中Pa 的表现比单播更差。与以上两个物种相比,Mr 在单播和混播中的表现差异是最小的(图3)。

图3 混播群落中各物种地上生物量的自然对数响应比(LNRR)在不同年份的表现Fig. 3 The values of natural logarithm response ratio (LNRR) of aboveground biomass about different species in all mixture of different years
2.3 物种组成和物种丰富度对植物性状的影响
与此同时,对比3 个物种在单播和混播中植物性状特征的变化,结果表明:从高度上来说,最高的物种En 没有显著的变化,Mr 的高度在所有混播类型中均显著提升(En+Mr: 33.63%; En+Mr+Pa: 6.60%; 图4a, c),Pa 的高度在3 个物种混播类型中显著下降(En+Mr+Pa: 21.74%; 图4c),这种变化使得3 种植物在地上空间上有着更加清晰的划分。从植株个数上来说,同单播相比,En 的植株个数在En+Pa 混播下显著增加了37.64%,在En+Mr 混播中显著增加了36.57%,在En+Mr+Pa 混播中显著增加了50.39%;Mr 在En+Mr 混播中下降了70.27%;Pa 在En+Pa 中下降了47.94%,在En+Mr+Pa 中下降了60.91%(全部:P<0.001; 图4d~f)。另外,同单播相比,在混播中Pa 的比叶面积值在En+Pa 和3 个物种混播中分别显著增加了29.77%和30.94%(P<0.05; 图4k, l),Pa 的叶片氮含量在3 个物种混播类型中显著增加了25.95%(P<0.001; 图4i),并且En 的叶片氮含量在En+Mr 混播类型中也显著增加了17.36%(P<0.05; 图4g)。

图4 不同播种方式(单播和混播)对植物性状的影响Fig.4 The effects of same plant species under different sowing mode (single and mixed sowing) on plant traits
3 讨论
本研究分析了物种丰富度对群落地上生物量的影响,发现在克隆植物群落中,地上生物量并非总是随着物种丰富度的提升而升高,并且这种关系会随着年份发生变化。尽管如此,在大多数情况下混播仍然比单播高产,这种净多样性效益主要来自TICE,也有一小部分来自随年份增强的TDCE 和DE。通过对植物特征进行分析发现,互补效应可能来自群落中各物种对地上垂直空间的更充分利用,并且处于底层的Pa 能够通过增加比叶面积和叶片氮含量来增强其光合作用以保证自身生物量。
在试验的第一年,随着物种丰富度的增加,群落生产力呈先上升后下降的趋势,这与在人工群落里发现的生物多样性-群落生产力之间的正相关关系不一致[3,20],这也是本研究发现的克隆植物群落同其他人工群落明显不同的地方。然而,同很多研究一样[8-9,47],本研究发现生物多样性效应随着时间增强,且物种越多这种增强趋势越显著。例如,同第一年相比,在第二年两物种混播的生物量增加了47.06%,3 个物种混播的生物量增加了64.09%,这使得试验中3 个物种混播的生物量比两物种混播增加的更多,最终导致两物种与3 个物种混播的生物量在第一年的显著性差异消失了。按照该趋势,能够预期在试验的第3 年会出现生物多样性与地上生物量之间的正相关关系,但是目前无法做出这样的判断,因为很多研究都证明这种随时间增长的多样性效应,却得到了不同的结果。在Van 等[8]的研究中,两物种混播和4 物种混播的多样性效应分别在第4 和5 年就达到饱和状态;而在Roscher 等[9]的研究中,6 个物种混播的多样性效应在第2 年就高于9 个物种混播。对于达到饱和的年限和涉及的混播物种数量上都有很大的不同,这可能源于两个研究选择了不同的物种:前者选择了除豆科以外的其他3 个功能群物种,而后者选择的是4 个功能群中的杂类草。虽然本研究无法断言物种的这种差异完全由物种带来的,不过物种的不同多少都会与这些结果有关联。因此本研究在多年生且能够克隆繁殖的植物构建的群落中可能会在后续试验中发现多样性效应随着时间产生的另外一种变化趋势(即达到饱和状态)也就具备合理性。想要找到其中更普遍的规律,还需要更多的研究。在播种的第二年,本研究并没有发现两物种混播与3 个物种混播之间的生物量差异,产生这种现象的原因是在两物种混播中NE 增加了104.86%,在3 个物种混播中NE 增加了260.01%。NE 随着时间增长已经被很多研究报道过[48]。
与很多理论性和试验性研究一致[34,49],多样性效应主要来源于TICE,在两物种混播中91.22%的NE 是由TICE 提供的;在3 个物种混播中83.71%的NE 是由TICE 提供的。TICE 的值越高说明物种之间的生态位互补性利用越充分或者物种间的助长作用越强。带来较高的TICE 有很多原因,例如营养的互补性利用,光资源的获取、环境异质性、植物-土壤反馈、植物地上或地下性状差异[20,49-50]。本研究分析了植物的不同性状来判断TICE的来源,发现植物的一些性状在混播中发生了显著的变化。首先,作为最高的物种,En 在所有单播和混播中都没有出现显著的高度。然而,Mr 和Pa 虽然在单播中有着近似的高度,但是表现出完全不同的模式。Pa 在En+Pa混播中高度没有显著变化,但是在En+Mr+Pa 中高度显著降低了21.74%;Mr 在En+Mr 混播中高度显著提升了33.63%,在En+Mr+Pa 混播中高度显著提升了6.60%。在两个两物种混播中,不管Pa 和Mr 是否发生高度的变化,其仍未达到En 的1/2 高度,垂直空间上的物种分布明显。在3 个物种混播中,两个原本高度接近的物种一个显著降低,一个略微升高,这种变化降低了两个物种在垂直空间分布上的重叠。物种的这种高度变化会带来3 种植物在空间上更明显的分层,有助于它们更有效的利用地上垂直空间[9,44],这有助于混播中的物种更加高效地获取光资源,进而带来生物量的增长。除此之外,当对比单播与混播之间单个物种的差异时发现处于顶层的En 的个体数是增加的,而处于混播中底层的植物(Mr 或者Pa)都是显著降低的。与此同时,在混播中一些表现越来越好的物种也将拥有越来越大的密度,其他的一些物种则会降低它们在上层空间的使用,这种不同空间层的密度变化会带来群落累计盖度的增加[51],这种地上空间的互补性利用同样有助于它们的混播进行互补性的、高效的光资源获取[52]。以上两个特征的变化可能是本研究得到较高TICE 的主要原因。本研究赞同Gravel 等[43]的观点,即物种需要时间来发展它的生物学特征,进而允许它们去填充各种各样的生态位,使得这些生态位能够得到更加充分的运用。
与此同时,本研究发现在所有混播中Pa 的SLA 值有明显的提升,且在3 个物种混播中叶片氮含量达到了显著的提升。这两个指标都与光合作用效率有着正向联系[53],说明底层物种光合效率的提升也为较高的TICE 做出贡献。本研究还发现,在En+Mr 混播中,En 的叶片氮含量显著提升,这可能与Mr 的固氮作用有关[40]。然而,本研究并没有在3 个物种混播中发现豆科植物的显著影响。其原因可能是豆科并非总是在正的多样性-生产力关系中发挥积极的作用[54],并且豆科植物的作用强度也有强弱之分[55]。可能本研究使用的豆科植物固氮作用较弱且植株在两物种混播中的密度高于3 个物种混播,因此只在两物种混播中略微显现出来。
显著大于零的LNRR 值说明该物种在混播中的表现要强于单播,即该物种的种内竞争强于种间竞争[20],这能够带来该物种在混播中优势的增加,进而带来TDCE 和DE 的改变。例如,当En 的优势增加不是以Pa 优势的降低为代价时,TDCE 增加;当以Pa 优势的降低为代价时,DE 增加。在2020 年的结果中,各个混播中各物种的LNRR 绝大部分都与零无显著差异,预示着各物种的优势没有发生大的改变,与该年份TDCE 和DE 基本接近于零的结果一致。在2021 年的结果中,En 的LNRR 值基本上都显著大于零(En+Pa 中En 的LNRR 虽然不显著,但是也在显著性的边缘:t=2.3574,P=0.07);Pa 的LNRR 值与零无差异。这个结果说明En 在混播中的优势随时间增加,这种增加在一些混播中并不一定以Pa 或者Mr 优势度降低为代价,因此TDCE 增加;在另外一些混播中,以Pa 优势度的降低为代价,导致DE 的增加。换句话说,TDCE 和DE 随着物种丰富度和时间的增加而增强。这与其他研究结果一致[50],但也与一些研究不同[38,56]。有的研究发现选择效应(即本研究中的TDCE+DE)在试验开始时为正,但是随着时间逐渐减弱[38];另一些研究发现选择效应并没有在群落中起到任何作用或起到微弱的作用,产生不同结果的原因可能与选择效应的来源有关。选择效应的增大主要源于混播中某个物种具备一些优势性状,能够比伴生的其他物种更高效地吸收和利用有限的资源,进而在混播中获得更高的优势[37],反之亦是如此。因此,如果物种不具备优势性状或者环境中不存在有限资源都可能观察不到选择效应的变化。在试验中出现以上结果可能是因为En 获取资源的能力比较强且群落中的资源比较有限,并且这两者都随着时间和物种丰富度增强。
4 结论
克隆植物作为草地生态系统的重要组成部分,在多样性-生产力关系中的表现可能会影响到整个生态系统这个关系的呈现。本研究表明,在克隆植物构建的群落中,物种丰富度对生产力的正向显著影响主要来自地上垂直空间的分层和个体密度的变化带来的互补性光资源利用。在这个过程中,优势植物优势度的不断增加也带来了TDCE 和DE 对生物多样性效应的加成作用。遗憾的是,本研究并没有在自然群落中研究克隆植物对这种关系的影响,未来这方面的研究能够深化对多样性-生态系统功能关系的理解,对于应对全球生物多样性丢失所带来的生态系统功能和服务的损失有着重要的意义。
猜你喜欢
科学技术与工程(2022年34期)2023-01-14 10:27:54
现代畜牧科技(2021年11期)2021-12-21 06:11:24
现代畜牧科技(2021年7期)2021-07-28 06:40:30
今日农业(2020年19期)2020-12-14 14:16:52
西藏农业科技(2019年1期)2019-07-25 00:37:00
杂草学报(2015年2期)2016-01-04 14:57:54
中国科技信息(2015年2期)2015-11-16 08:18:32
汽车文摘(2014年12期)2014-12-15 22:25:34
天津科技(2014年6期)2014-08-08 01:00:31
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
