
云曼红豆杉生长和次生代谢产物对光照胁迫的响应
2023-09-14 09:44孙志鹏李亚平余雨红唐淑琴蒲尚饶
西北农业学报 2023年9期
王 刚,孙志鹏,李亚平,余雨红,唐淑琴,刘 茜,郭 骐,蒲尚饶,王 刚
(1.四川农业大学 林学院,成都 611130;2.国家林业草原红豆杉西南工程技术研究中心,成都 611130;3.四川省林业科学研究院,成都 610081; 4.四川农业大学 风景园林学院, 成都 611130)
光照作为重要的环境因子,不仅对植物的生长发育具有重要的作用,而且影响植物的次生代谢过程,有利于药用植物次生代谢产物累积的调控[1-5]。红豆杉是一种具有重要经济价值的珍贵木本植物[6],被列为中国《主要林木育种科技创新规划(2016-2025年)》中珍贵树种育种工程的特色树种,是当今工业化生产抗癌药物紫杉醇及其衍生物(多西紫杉醇、卡巴他赛等)的唯一原料和世界第一抗癌植物,其紫杉醇全球市场规模预计将在2020年至2025年的预测期内年增长率为8.2%[7-11]。随着紫杉醇市场的巨大需求,同时为了更好地保护野生红豆杉资源[12-13],红豆杉被大量人工栽培和开发利用,但不同光照条件对药用植物红豆杉的品质差异较大[14-15]。因此,关于红豆杉栽培的研究受到广大科研人员的普遍关注。邓哲等[16]在研究不同土壤含水率对太行红豆杉生长及相关品质的试验中发现,80%~90%和60%~70%的土壤含水量分别有利于太行红豆杉的生长和多糖量的积累。孙志鹏等[17]通过测定不同海拔、不同季节和不同采收期的云南红豆杉(Taxusyunnanensis)枝叶中10-去乙酰基巴卡 亭Ⅲ(10-DAB)含量及枝叶生物量,结果表明,生长年限、生物量、季节和海拔均显著影响云南红豆杉的单株10-DAB累积量。此外,已有研究[18-20]分别以曼地亚红豆杉(Taxusmedia)和南方红豆杉(Taxuswallichianavar.mairei)为研究对象,研究不同处理方式对红豆杉的生长生理和主要次生代谢物积累的影响。目前,虽然国内外学者在营养元素、树龄等方面对红豆杉生长和次生代谢产物的影响进行了相关研究,但缺少在光照强度和采收期方面的探讨,尤其是光照对云曼红豆杉的生长和次生代谢产物的影响尚鲜见报道。
云曼红豆杉(Taxusmadia×T.yunnanensis‘Yunman’)良种是近年来选育的新品种,苗木(树龄<4a)枝叶中紫杉醇、10-DAB含量高于我国现有红豆杉,是生产天然紫杉醇和优质10-DAB的最好原料之一。因此,通过评价光照条件和采收时间对云曼红豆杉生长和主要次生代谢产物积累的影响,探究云曼红豆杉生长性状及其紫杉醇和10-DAB累积对不同光照环境的适应机制,为云曼红豆杉营建短周期优质药用原料林确立最佳光照条件及采收期提供科学依据。
1 材料与方法
1.1 试验区域概况
试验地位于四川省都江堰市胥家镇(东经103°41′42″,北纬31°0′30″),土壤类型为黄壤土。地势平整,平均海拔680 m,属中亚热带湿润季风气候,四季分明,无极端天气,年均降水量1 243 mm,年均温17.1 ℃,年均日照时数1 045 h,无霜期269 d。
1.2 试验材料
选用2 a生云曼红豆杉实生苗,由国家林业草原红豆杉西南工程技术研究中心提供,平均地径2.59 mm,平均苗高22.64 cm,单株生物量平均值为3.76 g。
1.3 试验设计
2019年12月,将云曼红豆杉苗木移栽至地势平坦且土壤水分和温度等生境条件均匀一致的实验地,栽植株行距0.55 m×0.50 m。利用不同遮阳网处理(0层、1层、2层、3层)进行光照梯度设置,共分为4个光区域,分别为0%遮光、30%遮光、60%遮光和90%遮光,遮阳网设置光区域长12 m、宽2 m、离地面1 m。苗木生长期间养分、水分持续供应充足,按照常规技术管理,定期除草,避免光照条件与其他环境因子的交互作用影响。自2020年3月中旬起,每间隔3个月,分别在4块不同光区域中随机测量和采集长势均匀的2 a生云曼红豆杉苗木,3株为1个重复,重复5次,每组采样15株/次,4组共计60株/次。
1.4 测定项目及方法
苗木地径、苗高分别采用电子游标卡尺和钢卷尺测定,测定结果保留小数点后两位。生物量(干质量)测定用烘干箱70 ℃将样品烘干至恒重,然后1/100电子天平精确称量,采用HPLC法测定活性成分紫杉醇和10-DAB含量[17,21],累积量即生物量和含量的乘积。2020年7月下旬选择晴朗天气,采用LD-GH80便携式光合仪从 6:00 时到18:00 时每间隔2 h测定1次不同处理的云曼红豆杉叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、CO2浓度(Ci)等指标。每处理重复测定3次。
1.5 数据分析
采用Microsoft Excel 2017软件进行数据处理,运用SPSS 19.0软件进行方差分析,采用Duncan’s法进行显著性检验,使用Origin 8.0进行绘图。
2 结果与分析
2.1 遮光对云曼红豆杉苗木生长的的影响
由表1可知,遮光和生长时间对云曼红豆杉苗木地径、苗高及生物量存在不同的影响。从光照强度来看,随着遮光的增强,各时期不同生长指标均呈先升高后降低的趋势,总体上60%遮光高于30%遮光、90%遮光和0%遮光。其中3月,60%遮光的苗木地径、苗高、生物量依次是0%遮光的1.18倍、1.08倍、1.25倍,30%遮光的1.01倍、1.02倍、1.05倍,90%遮光的 1.07倍、1.06倍、1.21倍。从生长时期来看,随着时间的增加,各生长指标逐渐升高,其中在0%遮光下,苗木地径、苗高、生物量分别在3月增长0.13 mm、1.10 cm、0.32 g,6月增长1.33 mm、3.76 cm、3.85 g,9月增长0.98 mm、1.80 cm、2.13 g,12月增长0.35 mm、0.98 cm、0.58 g。12月与3月相比在0%遮光下,苗木地径、苗高、生物量分别增长 97.8%、27.5%、1.61倍,30%遮光下分别增长 131.1%、51.4%、200.4%,60%遮光下分别增长1.43倍、54.5%、2.07倍,90%遮光下分别增长97.0%、35.7%、2.18倍。由此可得,2 a生云曼红豆杉苗木在60%遮光生长较好,6-9月是其生长旺盛期,12月各生长指标达到最大。
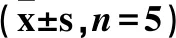
表1 不同遮光下苗木生长动态变化
2.2 遮光对云曼红豆杉苗木光合特性的影响
从图1可以看出,在不同遮光处理下,云曼红豆杉苗木的Pn在30%和90%遮光下日变化呈现“双峰型”曲线,有明显的“光午休”现象,受到了光抑制。其中:90%遮光下最明显,在8:00 时出现第1峰值为 7.96 μmol/(m2·s),之后随着光合有效辐射的增强,空气相对湿度逐渐下降,气温逐渐上升,Pn开始下降;在16:00时出现第2峰值为4.04 μmol/(m2·s),比第1峰值下降 49.25%。在60%遮光和0%遮光下,光合有效辐射相对较弱,未达到其光饱和点,苗木未发生“光午休”现象,其Pn日变化均为单峰曲线。对比云曼红豆杉苗木的Pn与Gs的日变化趋势,可以发现Pn与Gs有显著的正相关并且与其“光午休”对应,说明Pn对Gs具有反馈调节作用,当Gs增大时利于叶片的光合作用,相反,Gs减小不利于叶片的光合作用。不同遮光处理下Tr与Pn、Gs日变化趋势基本一致。此外,Ci在午间有所上升,Pn和Gs降低,Ci升高,这是一种非气孔 限制。
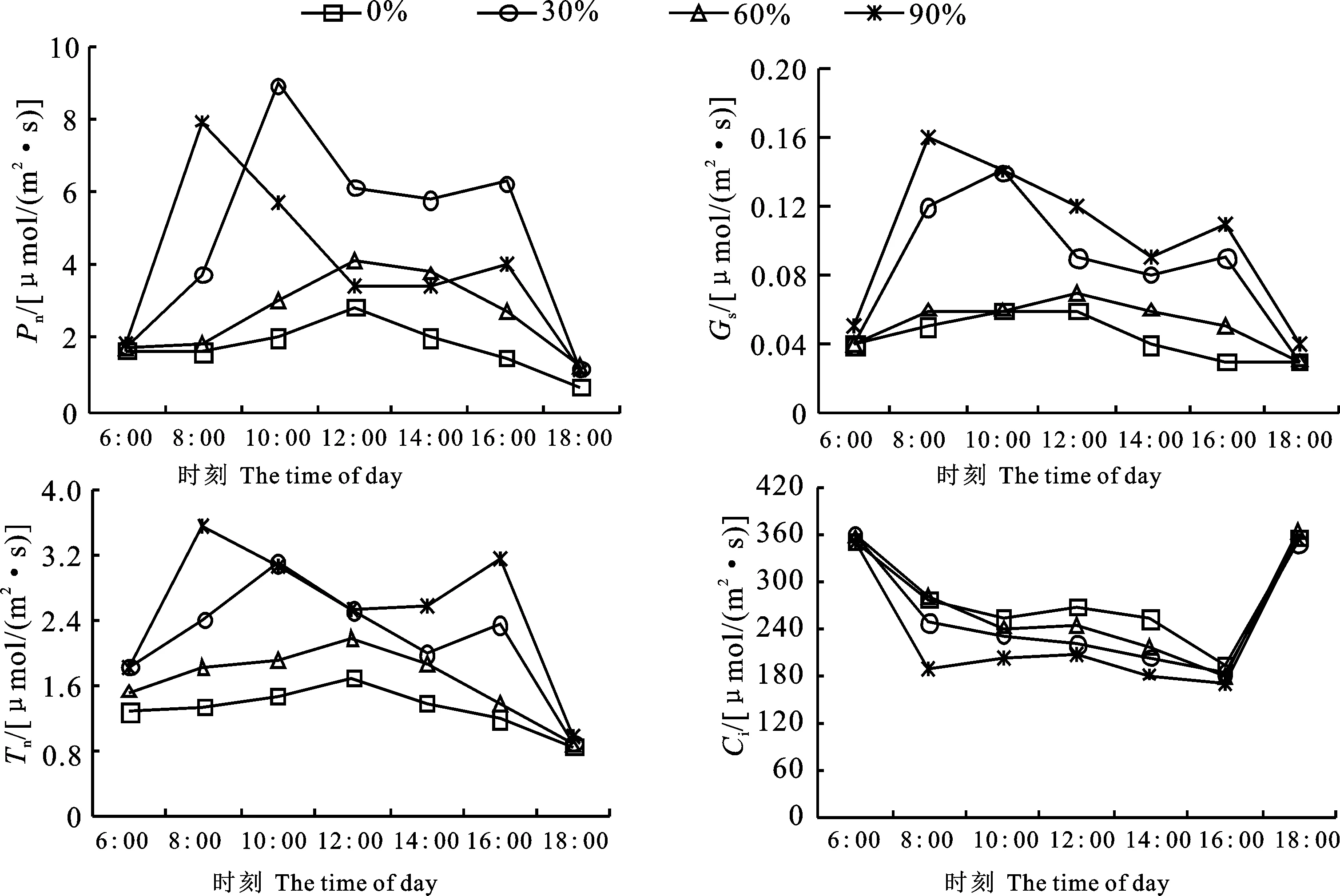
图1 不同遮光下苗木叶片光合参数日变化
2.3 遮光对云曼红豆杉苗木次生代谢产物的 影响
2.3.1 对云曼红豆杉苗木紫杉醇含量及单株累积量的影响 由图2-A可知,遮光对云曼红豆杉苗木枝叶中紫杉醇含量存在极显著差异(P< 0.01),随着遮光的增强,紫杉醇含量降低。4个遮光条件下的云曼红豆杉苗木枝叶中紫杉醇含量为 0.020%~0.036%,6月,各遮光下的云曼红豆杉苗木枝叶中紫杉醇含量均表现为最高,分别比3月、9月和12月同遮光增加7.58%、 7.57%、12.70%,有显著性差异(P<0.05)。由图2-B可知,遮光对云曼红豆杉单株紫杉醇累积量影响差异极显著(P<0.01)。4个遮光下的云曼红豆杉苗木枝叶中单株紫杉醇累积量为 1.15~4.51 mg;各遮光条件下苗木枝叶中单株紫杉醇累积量最高值均出现在9月,其中60%遮光最高,是0%遮光条件下的1.36倍。从生长时期来看,各测定月份间枝叶单株紫杉醇累积量差异极显著,4个遮光条件下的云曼红豆杉苗木单株紫杉醇累积量在9月都出现最高值,其次分别为12月、6月和3月。由此可得,不同遮光强度对云曼红豆杉苗木紫杉醇的合成具有一定影响,6月全光照条件更有利于苗木枝叶紫杉醇的合成;60%遮光有利于苗木单株紫杉醇的积累,9月是紫杉醇利用的最佳采收时间。
*表示差异显著(P<0.05);**表示差异极显著(P<0.01)。下同
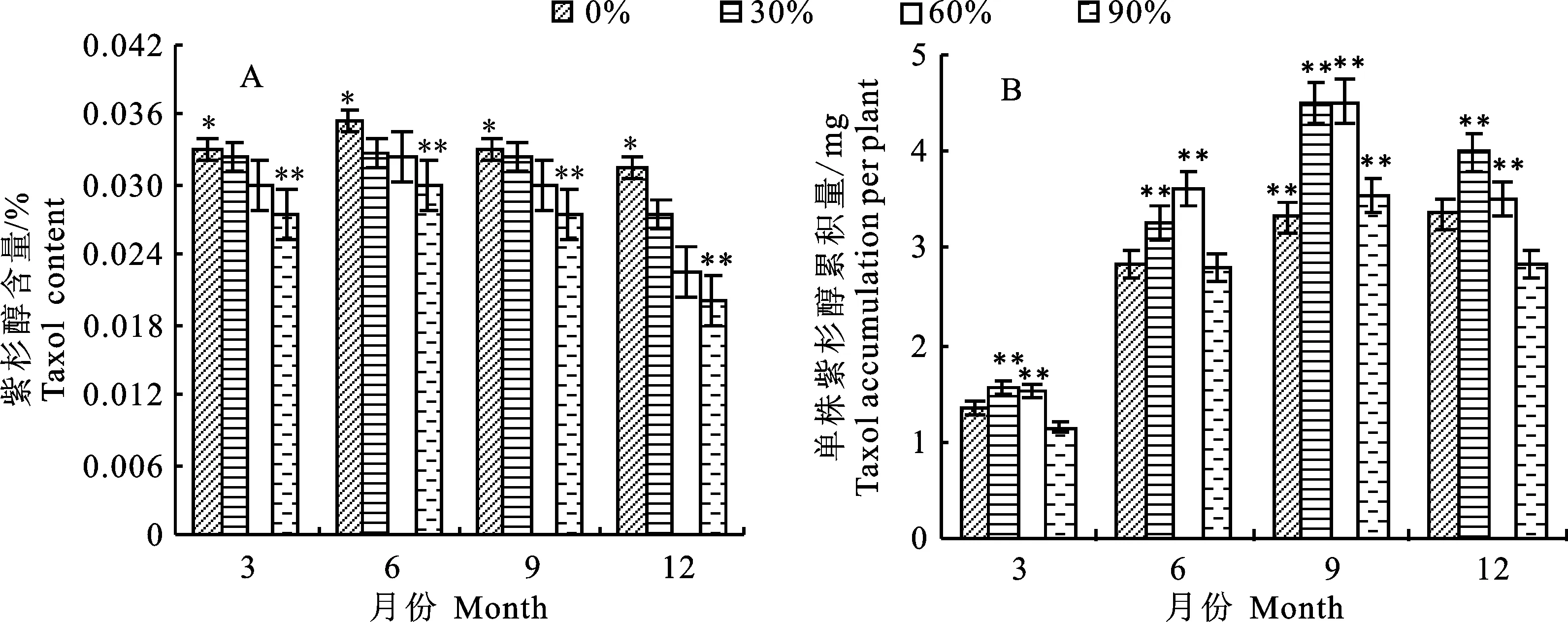
2.3.2 对云曼红豆杉苗木10-DAB含量及单株累积量的影响 由图3-A可知,遮光对云曼红豆杉苗木枝叶中10-DAB含量差异不显著,3月和9月,10-DAB含量随遮光增强而缓慢降低,6月和12月,10-DAB含量变化不明显。4个遮光条件下的云曼红豆杉苗木枝叶中10-DAB含量为 0.328%~0.383%,9月0%遮光达到峰值,分别是3月、6月和12月最低值的1.17倍、1.07倍和 1.08倍。由图3-B可知,遮光对云曼红豆杉苗木单株10-DAB累积量有显著影响。除3月外,60%遮光与0%遮光相比,单株10-DAB累积量存在极显著差异(P<0.01)。12月云曼红豆杉单株10-DAB累积量依次是:60%遮光>30%遮光>90%遮光>0%遮光,60%遮光是0%遮光下的1.46倍。从生长时间上看,6月、9月和12月与3月相比,苗木单株10-DAB累积量差异变化明显。分析相同遮光不同生长时期的单株10-DAB累积量可以发现:4个遮光条件的苗木单株10-DAB累积量均在12月出现较高峰值,其次分别为9月、6月和3月,这与苗木生长相似,均随着时间的增长而增加。由此可得,不同遮光强度对苗木枝叶10-DAB的合成具有一定影响,9月全光照条件更有利于苗木枝叶10-DAB的合成;60%遮光有利于苗木10-DAB的积累,12月是10-DAB利用的最佳采收时期。
图3 不同遮光下苗木10-DAB含量和单株累积量的动态变化
3 讨 论
3.1 不同光照下苗木生长节律的变化
不同遮光强度和生长时间对云曼红豆杉苗木的地径、苗高以及生物量积累存在显著影响。从遮光强度来看,总体60%遮光条件下地径、苗高、生物量最大,其增长量在不同季节依次高于30%遮光、90%遮光和0%遮光,这是由于适度遮光使到达地表的太阳总辐射量降低,同时冠层的温度、湿度及土壤等也会发生相应的变化[22];从生长时间来看,苗木的地径、苗高、生物量均随着时间的变化而变化,6月的3个生长指标的增长量在4个遮光下均大于其他月份,最佳光照的云曼红豆杉苗木地径增长达全年生长的53%,苗高增长占全年生长49%,生物量增长达全年生长的51%。其次为9月,生长最慢的是12月。这可能是因为夏季植物新陈代谢和光合作用增强,遮光降低了土壤中水分蒸腾及蒸发,从而为云曼红豆杉的生长发育提供适宜的光照水分条件。冬季气温较低,含水量减少,不利于植物的生长等原因[23]。本试验得出60%遮光适合2 a生云曼红豆杉生长,6月和9月是其生长旺盛期,这与彭晓邦[24]50%的光照适合商洛丹参生长吻合,与王磊等[25]关于须弥红豆杉在6-9月生长旺盛期的研究结果一致,可能原因是适度的遮光改善了空气相对湿度与二氧化碳浓度[26],使云曼红豆杉具有较高的相对生长率。
3.2 不同光照下苗木光合特性的变化
不同遮光强度和测定时间对云曼红豆杉的光合特性存在明显差异。30%遮光和90%遮光下的Pn日变化均呈现“双峰型”曲线,有明显的“光午休”现象。0%遮光条件下,其Pn日变化曲线为单峰曲线,光合有效辐射较弱,未达到其光饱和点,可能原因是强光、低湿、高温这种胁迫导致植物大量合成ABA等信号物质,植物为了自我保护关闭气孔,限制气体交换速率,从而Pn等光合参数降低[27];与30%遮光相比,60%遮光下云曼红豆杉各时刻叶片Pn、Tr、Gs均表现出不同程度的下降,但Ci升高,这可能是由于遮光条件下光照不足,叶片同化效率降低所致[28]。4个遮光处理下的云曼红豆杉Ci日变化在早晨和傍晚较高,但在中间时刻却相对较低,这正是云曼红豆杉通过光合作用利用CO2的重要体现。研究发现气孔限制是引起某一时段云曼红豆杉光合作用降低的原因,这与前人的研究结论基本一致[29-30],光照通过直接或间接作用影响植物光合作用[31-32]。随着光照强度增加,会导致光抑制的发生,同时会影响温度和空气相对湿度的变化。鉴于云曼红豆杉苗木生长对光照强度的响应特点,在造林时,宜进行林下种植或与其他树种进行混交栽植。因此,适度的遮光处理可降低温度和蒸腾量,减轻“光午休”现象,为云曼红豆杉快速生长创造良好的光照环境。
3.3 不同光照下苗木次生代谢产物的变化
不同遮光强度和生长时间对云曼红豆杉苗木的枝叶紫杉醇、10-DAB含量和单株累积量差异显著:全光照条件更有利于苗木枝叶紫杉醇、10-DAB的合成,但不利于其单株累积量的提高;9月,虽然60%遮光下枝叶紫杉醇含量偏低,但生物量较大。因此,60%遮光条件下苗木单株紫杉醇累积量最高,是对应0%遮光条件下的1.36 倍;12月,60%的遮光有利于苗木单株10-DAB累积量的积累,是对应0%遮光条件下的1.46倍;从生长时间上看,4个不同遮光处理下苗木枝叶紫杉醇、10-DAB含量及单株累积量年变化趋势基本上为先升高后降低,夏秋两季含量偏高,冬春两季含量偏低,这与曾小珂[33]关于云南红豆杉紫杉烷类化合物的动态变化规律研究结果类似。根据药用林木的栽培目的,综合考虑遮光强度以及采收时间,60%遮光有利于苗木紫杉醇和10-DAB含量累积,9-12月是最佳采收季节。本试验的次生代谢产物含量变化规律与孙志鹏等[17]、冯永林等[21]的研究结论基本一致,但紫杉醇单株累积量在9月达到最高的结论与杨逢建等[34]认为南方红豆杉枝叶中紫杉醇产量在11月末期达到最高不一致,可能原因是研究对象和采样时间不同,另外,光照强度影响红豆杉紫杉醇的生物合成及相关酶基因的表达[35],这也是影响紫杉烷类化合物累积量的重要因素。
4 结 论
光照对植物生长生理和次生代谢产物具有重要的作用。文章以云曼红豆杉苗木在不同光照强度开展生长和次生代谢产物动态变化研究,总体来看,2 a生云曼红豆杉实生苗木最适光照强度为60%遮光,生长旺盛期为6-9月,最佳采收时期为9-12月。研究结果对于云曼红豆杉良种的高效栽培与利用具有重要意义和应用价值。本研究仅利用光照单因素处理对云曼红豆杉苗木开展生长和主要次生代谢产物分析,未来将瞄准光照与施肥、水分等多因素交互作用对其生长代谢影响,开展更系统、深入的研究,揭示云曼红豆杉对不同环境的响应,为高品质云曼红豆杉培育管理和林下复合种植提供科学依据。
猜你喜欢
当代陕西(2020年17期)2020-10-28
西江月(2018年5期)2018-06-08
中成药(2018年3期)2018-05-07
中成药(2017年7期)2017-11-22
实用口腔医学杂志(2017年6期)2017-09-19
中外医疗(2016年15期)2016-12-01
广西林业科学(2016年4期)2016-03-16
哈尔滨医药(2015年2期)2015-12-01
党员生活(2015年6期)2015-07-16
传奇故事(破茧成蝶)(2015年1期)2015-02-28
