
光强和氮肥互作对南方软米粳稻灌浆结实期碳氮代谢影响及其与产量品质间关系
2023-09-14 09:34:16陈心怡马中涛张明月魏海燕张洪程刘国栋李光彦许方甫
作物学报 2023年11期
陈心怡 朱 盈 马中涛 张明月 魏海燕 张洪程 刘国栋 胡 群 李光彦 许方甫
光强和氮肥互作对南方软米粳稻灌浆结实期碳氮代谢影响及其与产量品质间关系
陈心怡 朱 盈 马中涛 张明月 魏海燕*张洪程 刘国栋 胡 群 李光彦 许方甫
扬州大学江苏省作物遗传生理重点实验室 / 农业农村部长江流域稻作技术创新中心 / 江苏省粮食作物现代产业技术协同创新中心, 江苏扬州 225009
以南方软米粳稻南粳9108和扬农香28为材料, 设置2个光强处理和4种氮肥处理, 其中光强处理于结实期展开, 分为100%自然光照强度(L1)和50%自然光照强度(L2), 氮肥处理为生长中后期不施氮肥(N1), 分别于倒六叶(N2)、倒四叶(N3)、倒二叶(N4)期一次性施用氮肥, 研究了不同光照强度、氮肥施用时期及光氮互作条件下水稻结实期碳氮代谢差异及其对产量和品质的影响。结果表明, 结实期光强减弱, 剑叶净光合速率下降7.35%~42.36%、叶片中蔗糖磷酸合酶(SPS)和蔗糖合酶(SS)酶活性下降, 叶片C/N比下降3.98~6.49, 光合产物向籽粒输送减少, 籽粒中淀粉(包括直链淀粉)含量降低, 同时叶片中硝酸还原酶(NR)、谷氨酰胺合酶(GS)、谷氨酸合酶(GOGAT)活性增强, 植株含氮率提升, 蛋白质含量相对增加, 不利于产量和优良品质的形成。中后期施用氮肥后, 叶片中碳氮代谢关键酶活性显著提高、叶片的衰老减缓, 水稻的灌浆结实期延长, 有利于产量的提升。随中后期氮肥施用时期推迟, 氮代谢愈发旺盛, 籽粒中蛋白质含量相对明显增加, 导致淀粉与蛋白质、直链淀粉与蛋白质的比值均下降, 食味值下降。本试验条件下, 正常光照配合倒四叶施用氮肥处理(L1~N3)能够协同提高叶片碳氮代谢关键酶活性, 使得光合产物和含氮化合物以适宜的比例向籽粒输送, 最终籽粒中淀粉与蛋白质的比值在11.43~12.03之间, 直链淀粉与蛋白质的比值在1.34~1.50之间, 米饭的硬度低, 黏度、平衡性高, 食味好, 可同时获得高产和优质。
水稻; 光照; 氮肥; 碳氮代谢; 产量; 品质
我国水稻的种植面积在3000万公顷左右, 其中粳稻种植面积占水稻种植总面积的24.3%, 且主要作为口粮供给, 对保障国家粮食安全具有重要作用。“秦岭-淮河”以南的南方粳稻产区具有单产水平高、总产量大的特点。但与北方粳稻相比, 南方粳稻的稻米品质尤其是食味品质有待提高。近年来, 南方水稻育种家为了满足人们对于米饭“吃得好”的需求, 培育出了一批直链淀粉含量较低、食味较好的优质软米粳稻品种。
水稻籽粒胚乳淀粉和蛋白质合成和积累是影响水稻产量和品质形成的重要因素[1], 其含量高低、结构比例和分布状态都与植株体内的碳氮代谢过程密切相关[2-6]。水稻的碳氮代谢不仅取决于品种自身特性, 还受氮肥、光照等环境的影响。氮是水稻需求量最大的矿质元素, 也是植物体内许多化合物的重要组分。合理的N素有利于促进地下部根系和地上部器官的协同生长[7], 构建良好的植株形态, 影响叶片中叶绿体的结构, 提高植株光合作用强度, 增加颖花量[8-9], 延长同化物生产持续期, 促进植株对碳氮的同化和吸收能力。吕腾飞等[8]、吕川根等[10]、孙永健等[11]、从夕汉等[12]研究发现, 增施氮肥以及适当的氮肥后移, 能够增强水稻生长过程中的碳氮代谢能力, 协调蔗糖磷酸合酶(SPS, EC 2.4.1.14)、蔗糖合酶(SS, EC 2.4.1.13)、硝酸还原酶(NR, EC 1.7.99.4)、谷氨酰胺合酶(GS, EC 6.3.1.2)等碳氮代谢相关酶之间的活性, 进而促进植株对碳氮的同化和吸收能力, 实现水稻产量和氮肥利用率的同步提高。氮肥施用在影响植株碳氮代谢的同时还会改变籽粒淀粉和蛋白质的比例, 引起蛋白质含量增加而降低稻米的食味品质[13-14]。光照是影响水稻光合作用和生长发育的重要因素[15-16]。一般认为, 光强减弱会导致水稻产量降低, 且不同时期光强减弱对水稻产量的影响存在差异[17-19], 姜楠[20]认为抽穗后弱光对水稻产量影响最大。同时, 植株体内碳氮代谢也会因为光强减弱而变化。遮光后, 叶片中1,5-二磷酸核酮糖羧化酶/加氧酶(Rubisco, EC 4.1.1.39)活性及叶片中叶绿素/值下降[21-22], 叶片衰老速度减缓, 叶绿素含量增加, 光合速率下降, 碳代谢能力减弱。此外, 水稻的根系活力及根干重受遮光影响显著降低, 氮素吸收能力下降[23], 但遮光条件下的光以蓝紫光为主, 促进含氮化合物的合成, 提高植株含氮量, 增强植株的氮同化能力[24-27]。尽管前人关于水稻碳氮代谢已有一定研究, 但主要集中在环境因素对常规水稻碳氮代谢的影响或水稻产量和氮肥吸收利用对碳代谢或氮代谢单要素的响应上。而关于碳氮代谢对南方优质食味水稻的新类型—软米水稻产量、品质综合影响的研究尚有所缺乏, 为此, 本研究以优质食味软米粳稻南粳9108和扬农香28为材料, 设置中后期不同时期施用氮肥、结实期不同光强处理, 研究不同光、氮及光氮互作条件下南方软米粳稻结实期碳氮代谢差异及其这种差异对软米粳稻产量和品质的影响, 以期为南方软米粳稻优质丰产栽培提供一定的理论依据。
1 材料与方法
1.1 试验地点及供试材料
试验于2019—2020年在扬州大学试验农场(32°39′N, 119°42′E)进行, 土壤类型为沙壤土, 地力较为良好, 土壤0~20 cm耕层有机质含量为28.3 g kg–1、碱解氮177.6 mg kg–1、速效磷52.8 mg kg–1、速效钾202 mg kg–1。供试品种为优质食味软米粳稻南粳9108和扬农香28, 均属迟熟中粳软米粳稻品种。
1.2 试验设计
本研究采用裂区设计, 品种为主区, 结实期光照强度和氮素水平为裂区。如表1所示, 结实期光照强度设置2个处理: 100%自然光照强度(L1)和50%自然光照强度(L2), 其中, L2处理采用透光率为50%的黑色遮阳网在水稻抽穗至成熟期进行遮光处理。光强处理用照度计(LI-250型; LI-COR, 美国)于08:00、12:00和16:00测量其网下透射率分别为52%、48%和51%, 平均50%。试验各小区基蘖肥用量一致,基肥、分蘖肥各施纯氮94.5 kg hm–2, 中后期设置4种氮肥处理: 不施氮肥(N1), 分别为于倒六叶(N2)、倒四叶(N3)、倒二叶(N4)一次性施用氮肥, 其中氮肥用量为纯氮81 kg hm–2, 每处理重复2次, 小区面积30 m2。氮(N)∶磷(P5O2)∶钾(K2O)比例为2∶1∶2, 磷肥一次性基施, 钾肥分别于耕翻前、拔节期等量施入。氮肥来源为尿素(N含量为46%), 磷肥来源为过磷酸钙(P2O5含量为12%), 钾肥来源为氯化钾(K2O含量为60%)。每个氮肥处理大区之间筑35~40 cm土埂并用塑料薄膜包埂, 单独排灌。水分管理及病虫草害防治等相关的栽培措施均按照优质高产栽培要求实施。其中, 2019年和2020年试验分别于5月25日和5月22日播种。
所用育秧盘为毯苗机插专用硬盘, 育秧期间肥水管理遵循旱育秧原则, 2年分别于6月13日和6月15日人工模拟毯苗插秧机移栽, 秧龄分别为19 d和24 d, 栽插行株距为30 cm × 12 cm, 每穴4株苗。
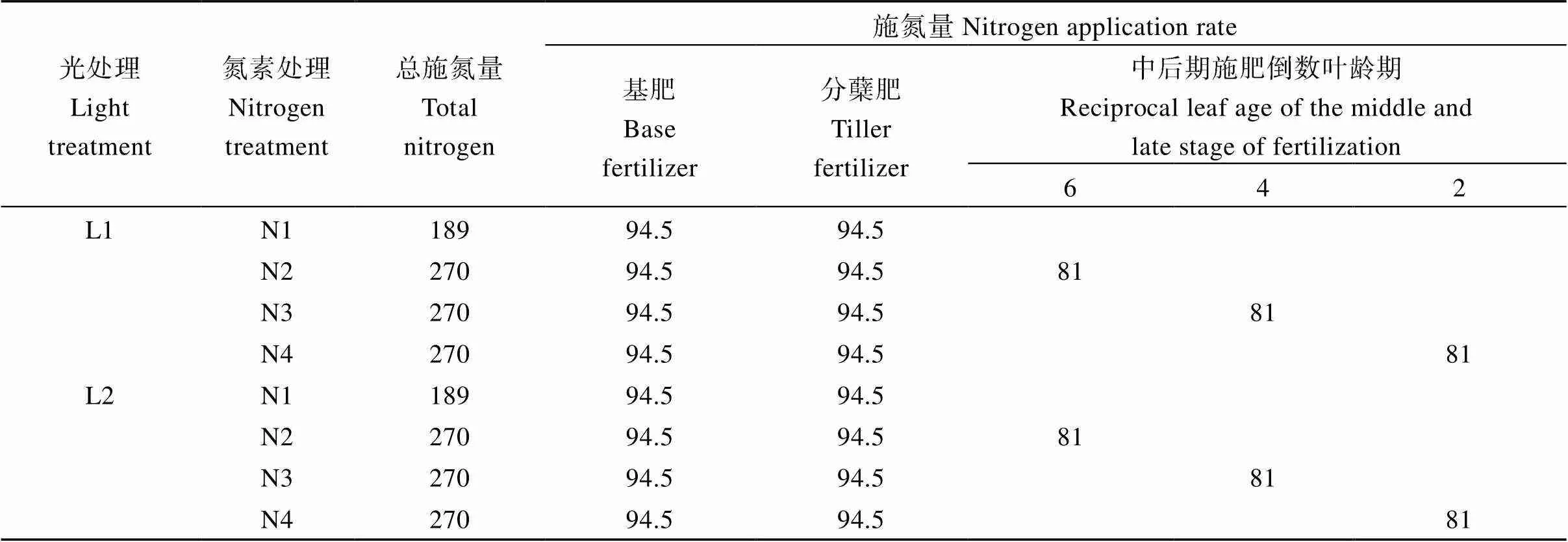
表1 不同光强处理下氮肥用量
1.2 测定内容与方法
1.2.1 叶片光合特性的测定 自开花至花后42 d,每隔7 d选取标记的同日开花的植株, 使用LI- 6400XT 便携式光合仪测定剑叶的净光合速率, 测定时各处理均选取剑叶的相同位置。
1.2.2 叶面积指数 分别于拔节期、抽穗期和成熟期, 根据各小区的每穴平均茎蘖数取具有代表性的植株3穴, 测定植株绿叶面积, 进而计算出叶面积指数。
1.2.3 碳、氮代谢相关酶活性 自开花至花后42 d, 每隔7 d取标记叶片液氮中速冻后置于–70℃超低温冰箱中保存。蔗糖成酶、蔗糖磷酸合酶、硝酸还原酶、谷氨酰胺合酶、谷氨酸合酶活性的测定使用上海酶联生物科技有限公司提供的相关试剂盒并按照相应方法进行测定。
1.2.4 可溶性糖及淀粉含量 自开花至花后42 d,每隔7 d取标记植株的叶片和籽粒105℃杀青0.5 h, 80℃烘干至恒重后粉碎并过100目筛, 称取0.1 g样品至10 mL离心管, 加入8 mL配制的80%乙醇溶液80℃水浴30 min, 冷却后离心, 重复3次, 合并上清液用于测定可溶性糖, 残渣用于测定淀粉, 采用蒽酮比色法测定可溶性糖和淀粉含量[28]。
1.2.5 高效叶片SPAD值的测定 高效叶片指水稻植株有效茎蘖抽穗后上3张叶片。自开花至花后42 d, 每隔7 d选取标记的同日开花的植株, 采用叶绿素仪SPAD-502仪(日本柯尼卡美能公司)测定标记植株高效叶片的SPAD值, 取其平均值。
1.2.6 含氮率 自开花至花后42 d, 每隔7 d选取标记植株3穴, 将所取植株样品于105℃杀青30 min, 80℃烘至恒重后测定干物质重。样品粉碎后采用H2SO4-H2O2消化, 半微量凯氏定氮法测定植株含氮率。
1.2.7 产量及其结构 成熟期时, 每小区随机选取5个点(每点15穴), 调查有效穗数, 按各小区每穴平均穗数取5穴以考查每穗粒数、结实率、千粒重(取1000粒称重, 重复3次, 误差不超过0.05 g)。每小区割取100穴, 脱粒、去杂后晾晒至含水量14.5%时测定籽粒重量, 计算产量。
1.2.8 糙米直链淀粉、总淀粉含量 取标记的同日开花的稻穗, 穗上籽粒出糙后粉碎, 购买 Megazyme 试剂盒并参照相应方法测定直链淀粉和总淀粉含量。
1.2.9 籽粒中蛋白质含量 成熟期取标记穗, 穗上籽粒出糙后粉碎, 籽粒中蛋白质含量采用凯氏定氮法测定(Kjeltec 8200, Foss, Hillerod, 丹麦)。
1.2.10 稻米品质 参照国家标准《GB/T17891- 1999优质稻谷》和《GB/T17891-2017优质稻谷》方法, 测定出糙率、出精率、整精米率、垩白粒率、垩白大小、垩白度。采用米饭食味计(STA1A, 日本佐竹公司)测定米饭的食味参数。
1.3 数据计算与统计分析
叶面积衰减率=叶面积衰减率(LAI d–1)= (LAI2–LAI1)/(t2–t1), 式中, LAI1和LAI2分别为抽穗期和成熟期测定的叶面积指数, t1和t2分别为抽穗期和成熟期测定的时间; C/N[29]=可用性糖的积累量(可溶性总糖+淀粉)/氮素的积累量; 氮素积累量(kg hm–2)=干物质积累量×含氮率。
用Microsoft Excel 2016进行数据的录入和计算,运用SPSS软件进行统计分析, 用新复极差法SSR进行多重比较(<0.05), 产量及品质2年结果趋势一致, 年度间差异未达显著水平, 取2年平均值进行分析, 其余对测定年度数据进行分析; 以Origin 2018软件作图。
2 结果与分析
2.1 不同光、氮条件下南方优质粳稻碳代谢的差异
2.1.1 剑叶净光合速率 50%光强处理下水稻花后剑叶的净光合速率显著降低, 供试2个品种的变化基本一致, 降幅在7.35%~42.36% (表2)。中后期施用氮肥后, 花后2个软米粳稻品种的净光合速率显著提高, 并且升幅在正常光照强度处理下会随着中后期氮肥施用时期的推迟逐渐增加, 与N1处理相比, N2、N3、N4分别增加了2.90%~7.96%、5.61%~ 14.37%、8.84%~25.09%, 而在50%光强处理下升幅则随着中后期氮肥施用时期的推迟逐渐减少。2种光强处理下, 2个品种的净光合速率均以L1N4处理最高。除开花期外, 光强、氮素及其互作效应对剑叶净光合速率的影响均达到了显著水平。
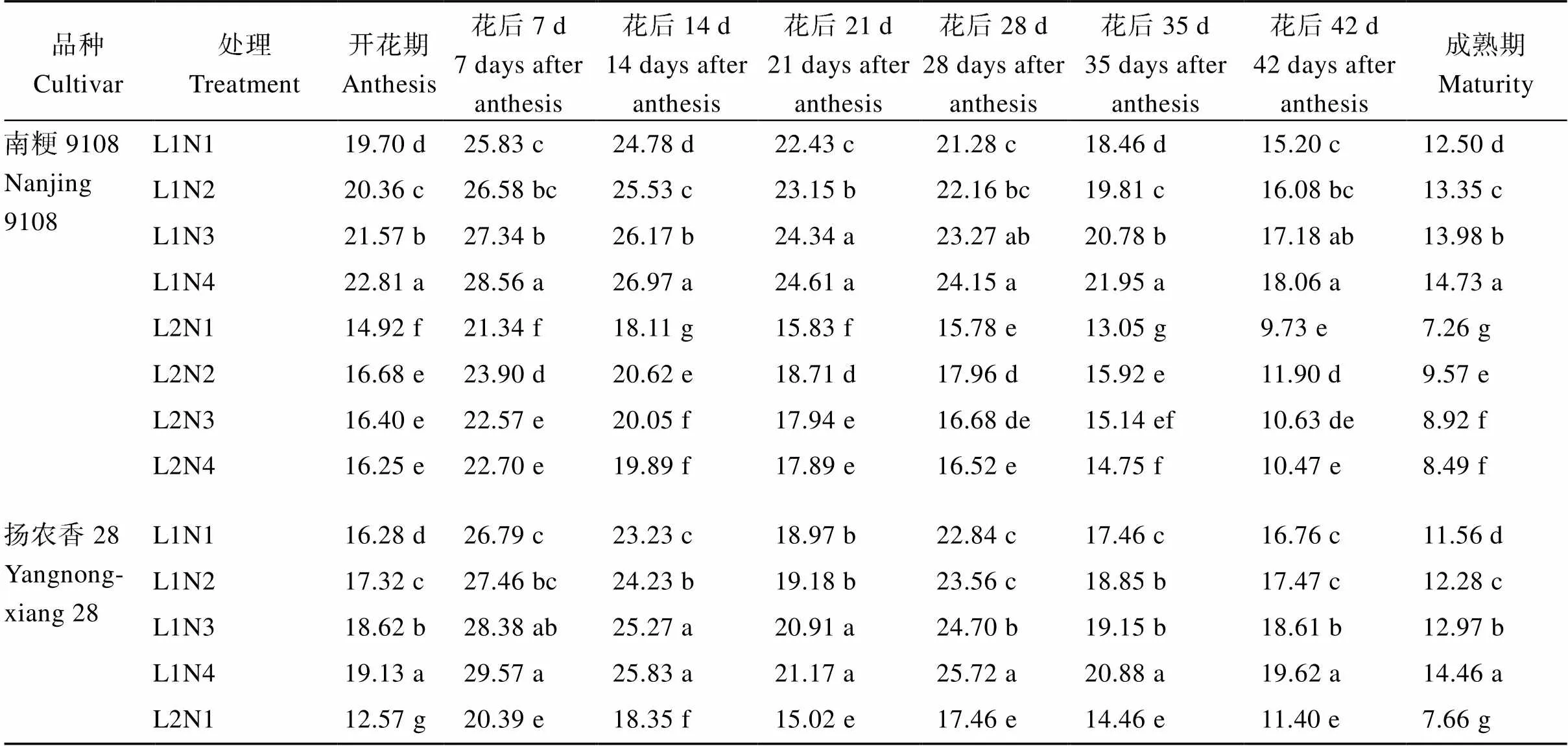
表2 不同光、氮条件下南方软米粳稻花后剑叶净光合速率的差异
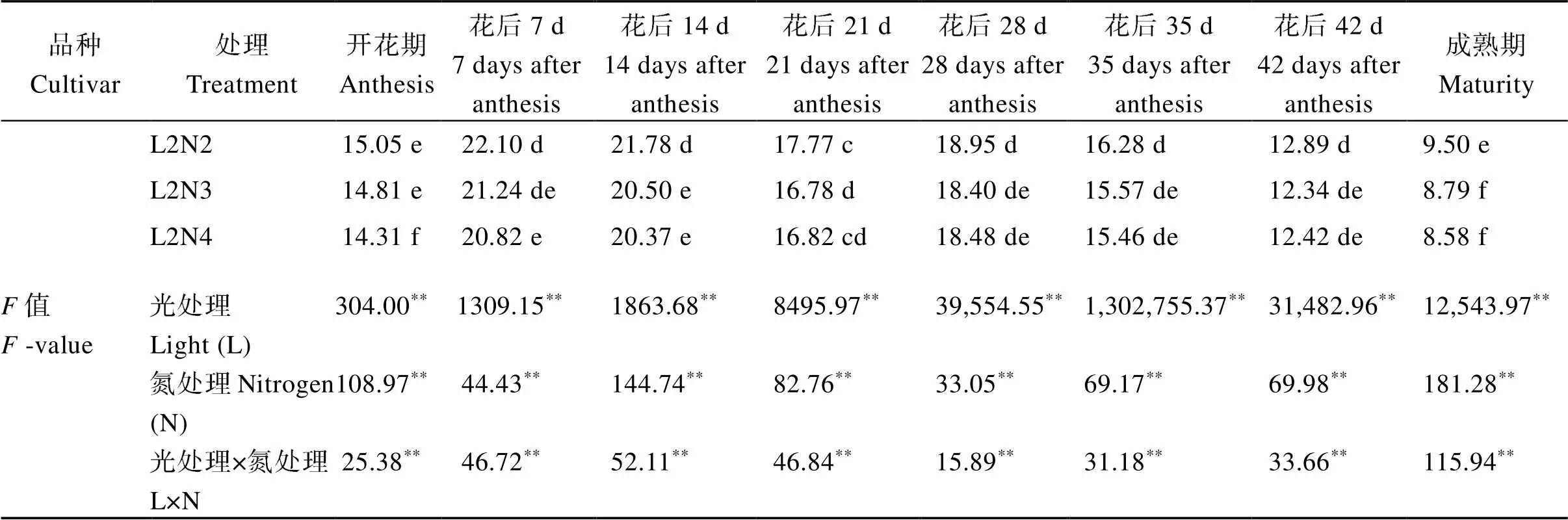
(续表2)
标以不同小写字母的值分别表示同一品种不同处理在0.05概率水平差异显著。*和**分别表示在0.05和0.01概率水平差异显著。
Different lowercase letters indicate that there are significant difference in the 0.05 probability level in different treatments of the same variety.*and**indicate significant differences at the 0.05 and 0.01 probability levels, respectively.
2.1.2 叶面积指数及叶面积衰减率 50%光强处理下水稻拔节期和抽穗期的叶面积指数在处理间差异不显著(表3), 成熟期叶面积指数显著提高, 供试2个品种的变化基本一致, 升幅在7.89%~29.98%。中后期施用氮肥后, 2个品种叶面积指数显著提高, 结实期叶面积衰减率降低。随着中后期氮肥施用时期的推迟, 2个品种拔节期叶面积指数逐渐下降, 抽穗期叶面积指数先增后减, 并以N3处理最大, 成熟期叶面积指数在不同光强处理下表现不一致, 在正常光强处理表现为先升后降, 在50%光强处理下则逐渐下降。2个品种结实期叶面积衰减率在正常光强处理下随中后期氮肥施用时期的推迟逐渐降低, 在50%光强处理下表现为先增后减。

表3 不同光、氮条件下南方软米粳稻叶面积指数及叶面积衰减率的差异
标以不同小写字母的值分别表示同一品种不同处理在0.05概率水平差异显著。*和**分别表示在0.05和0.01概率水平差异显著, NS表示差异不显著。
Different lowercase letters indicate that there are significant difference in the 0.05 probability level in different treatments of the same variety.*and**indicate significant differences at the 0.05 and 0.01 probability levels, respectively, and NS indicates no significant differences.
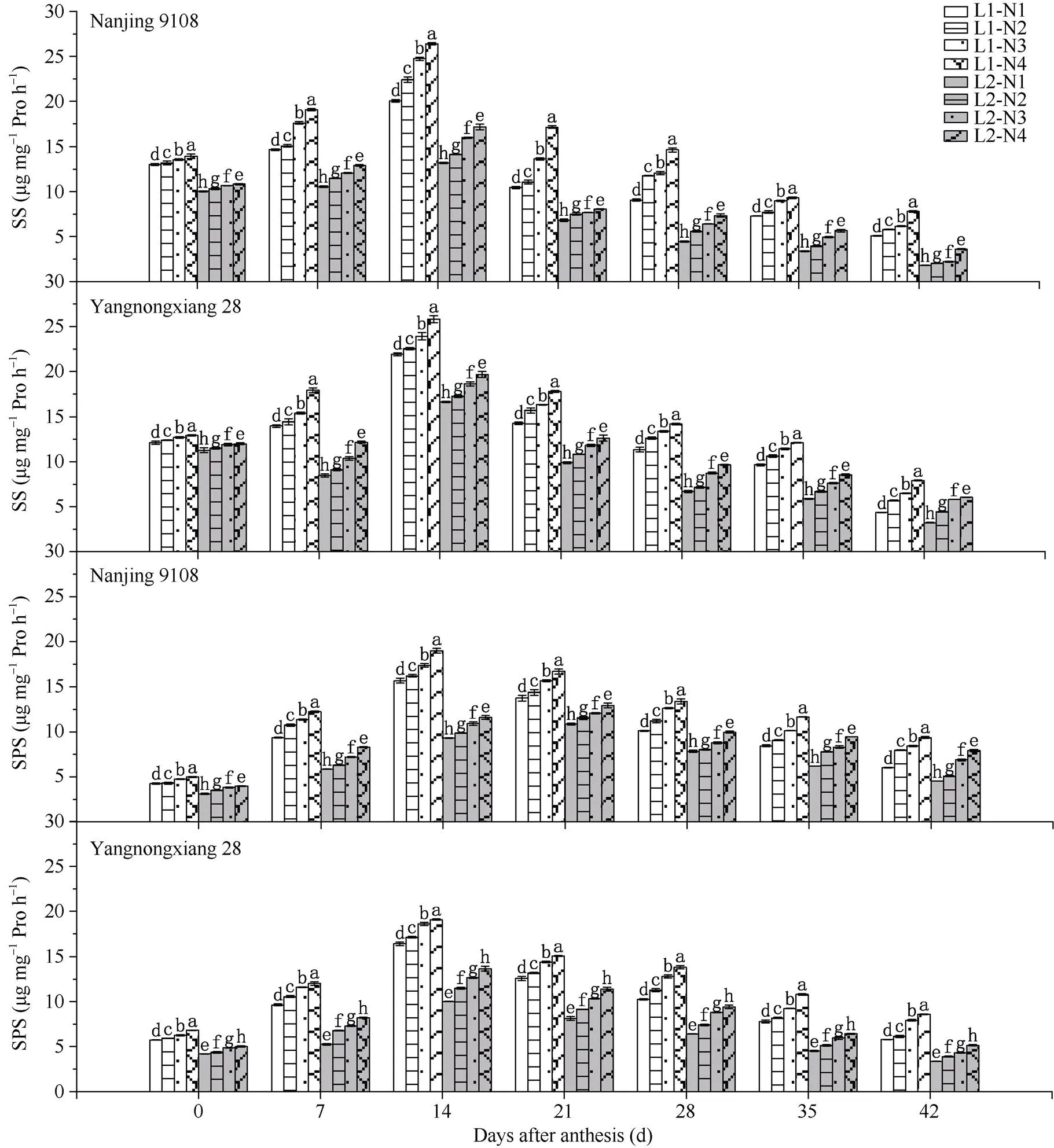
图1 不同光强、氮肥条件下南方软米粳稻花后叶片中蔗糖合酶(SS)、蔗糖磷酸合酶(SPS)的差异
图中“L1, L2”分别指结实期100%自然光照强度和50%自然光照强度, “N2, N3, N4”分别指中后期于倒六叶, 倒四叶, 倒二叶施用氮肥, “N1”指中后期不施氮肥处理。图中所标注的字母为蔗糖合酶和蔗糖磷酸合酶活性的显著性, 不同字母表示在0.05概率水平差异显著。
“L1, L2” in the figure refer to 100% natural light intensity and 50% natural light intensity at grain-filling stage, respectively. “N2, N3, and N4” refer to applying nitrogen fertilizer to 6 leaves, 4 leaves, and 2 leaves at middle and late stage, respectively. “N1” refers to not applying nitrogen fertilizer at middle and late stage. Different lowercase letters indicate significantly different at the 0.05 probability level for the activity of sucrose synthetase and sucrose phosphate synthetase.
2.1.3 碳代谢相关酶 50%光强处理下供试2个品种叶片中碳代谢相关的蔗糖合酶(SS)、蔗糖磷酸合酶(SPS)的活性均显著降低(图2)。中后期施用氮肥后, SS、SPS活性显著增强。随着中后期氮肥施用时期的推迟, SS、SPS的活性的升幅逐渐增加。光、氮共同处理下, 南粳9108和扬农香28叶片中SS、SPS活性均呈现L1N4>L1N3>L1N2>L1N1>L2N4> L2N3>L2N2>L2N1的趋势。

图2 不同光、氮条件下南方软米粳稻花后叶片中可溶性糖和淀粉含量的差异
处理同图1。图中所标注字母为叶片淀粉和可溶性糖含量的显著性, 不同字母表示在0.05概率水平差异显著。
Treatments are the same as those given in Fig. 1. Different lowercase letters indicate significantly different at the 0.05 probability level in the content of soluble sugar and starch in leaves.
2.1.4 叶片可溶性糖和淀粉含量 50%光强处理下供试2个品种水稻叶片中可溶性糖和淀粉含量均显著降低(图3)。中后期施用氮肥后, 叶片中可溶性糖和淀粉含量显著下降。随着中后期施肥时间的推迟, 叶片中可溶性糖和淀粉含量的降幅逐渐增大, 与N1处理相比, N2、N3、N4可溶性糖含量分别降低了1.51%~7.97%、4.43%~13.25%、9.24%~21.69%, 淀粉含量分别降低2.85%~10.36%、4.28%~17.15%、5.44%~23.04%。光、氮共同处理下, 南粳9108和扬农香28花后7~42 d叶片中可溶性糖和淀粉含量均呈现L1N1>L1N2>L1N3>L1N4>L2N1>L2N2>L2N3> L2N4的趋势。
2.1.5 籽粒可溶性糖和淀粉含量 50%光强处理下供试2个品种水稻籽粒中可溶性糖和淀粉含量均显著降低(图4)。中后期施用氮肥后, 籽粒中可溶性糖和淀粉含量均显著下降。随着中后期施肥时间的推迟, 籽粒中可溶性糖和淀粉含量的降幅逐渐增大, 与N1处理相比, N2、N3、N4可溶性糖含量分别降低了1.23%~9.64%、3.44%~13.47%、4.13%~18.81%, 淀粉含量分别降低了1.40%~7.18%、1.73%~12.30%、3.44%~16.92%。光、氮共同处理下, 南粳9108和扬农香28花后7~42 d籽粒中可溶性糖和淀粉含量均呈现L1N1>L1N2>L1N3>L1N4>L2N1>L2N2>L2N3> L2N4的趋势。
2.2 不同光、氮条件下南方优质粳稻氮代谢的差异
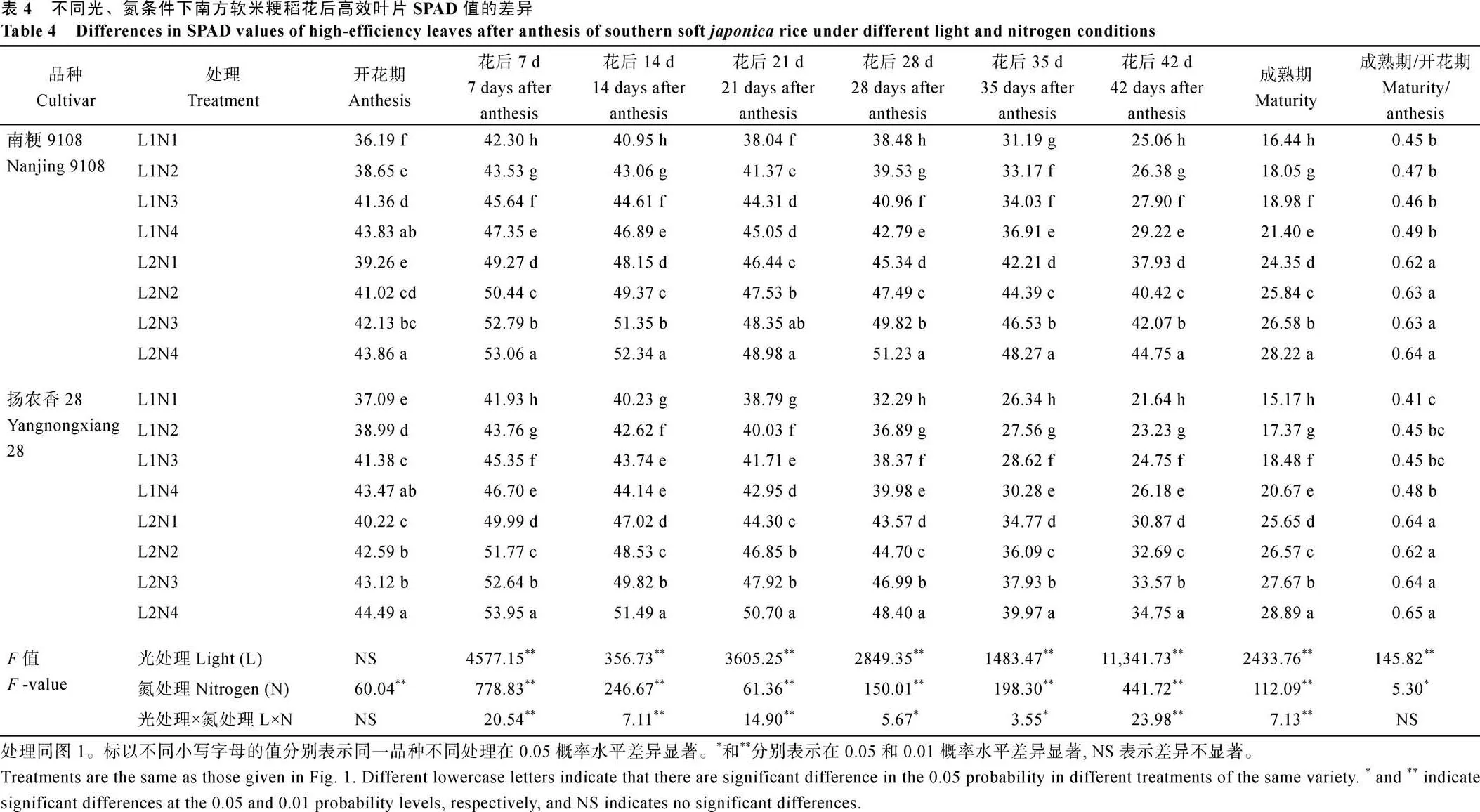
2.2.1 花后高效叶片SPAD值 50%光强处理下水稻花后高效叶片的SPAD值显著升高(表4), 供试2个品种的变化基本一致, 升幅在1.5%~69.0%, 且成熟期与开花期的SPAD比值显著增加。中后期施用氮肥后, 2个品种花后高效叶片的SPAD值显著升高, 且升幅随着中后期氮肥施用时期的推迟逐渐增加, 与N1处理相比, N2、N3、N4分别增加了5.44%~ 7.0%、8.48%~12.65%、12.73%~18.85%。2个品种SPAD值均呈现L2N4处理最高, 相较于L1N1处理增加了17.25%~90.45%。
2.2.2 花后叶片、植株含氮率及N素积累 50%光强处理下供试2个品种花后植株及叶片的含氮率均显著升高(表5和表6); 中后期施用氮肥后, 植株及叶片的含氮率显著上升, 随中后期氮肥施用时期的推迟升幅逐渐增加, 与N1处理相比, N2、N3、N4处理植株含氮率分别增加了6.46%~9.25%、12.24%~ 19.59%、17.81%~30.33%, 叶片含氮率分别增加了6.45%~12.08%、9.83%~18.67%、16.04%~23.70%。此外, 50%光强处理下中后期施用氮肥对植株含N率的增加效应更加显著。2个品种花后植株及叶片的含氮率均以L2N4处理最高, 相较于L1N1处理, 植株含氮率增加了15.60%~73.40%, 叶片含氮率增加了24.53%~57.07%。抽穗至成熟期的氮素积累, 2个品种均在L1N3处理下积累量最高。光、氮及光氮互作对植株及叶片含氮率的影响在开花期以后均达到极显著水平, 对抽穗至成熟期的氮素积累的影响也达极显著水平。
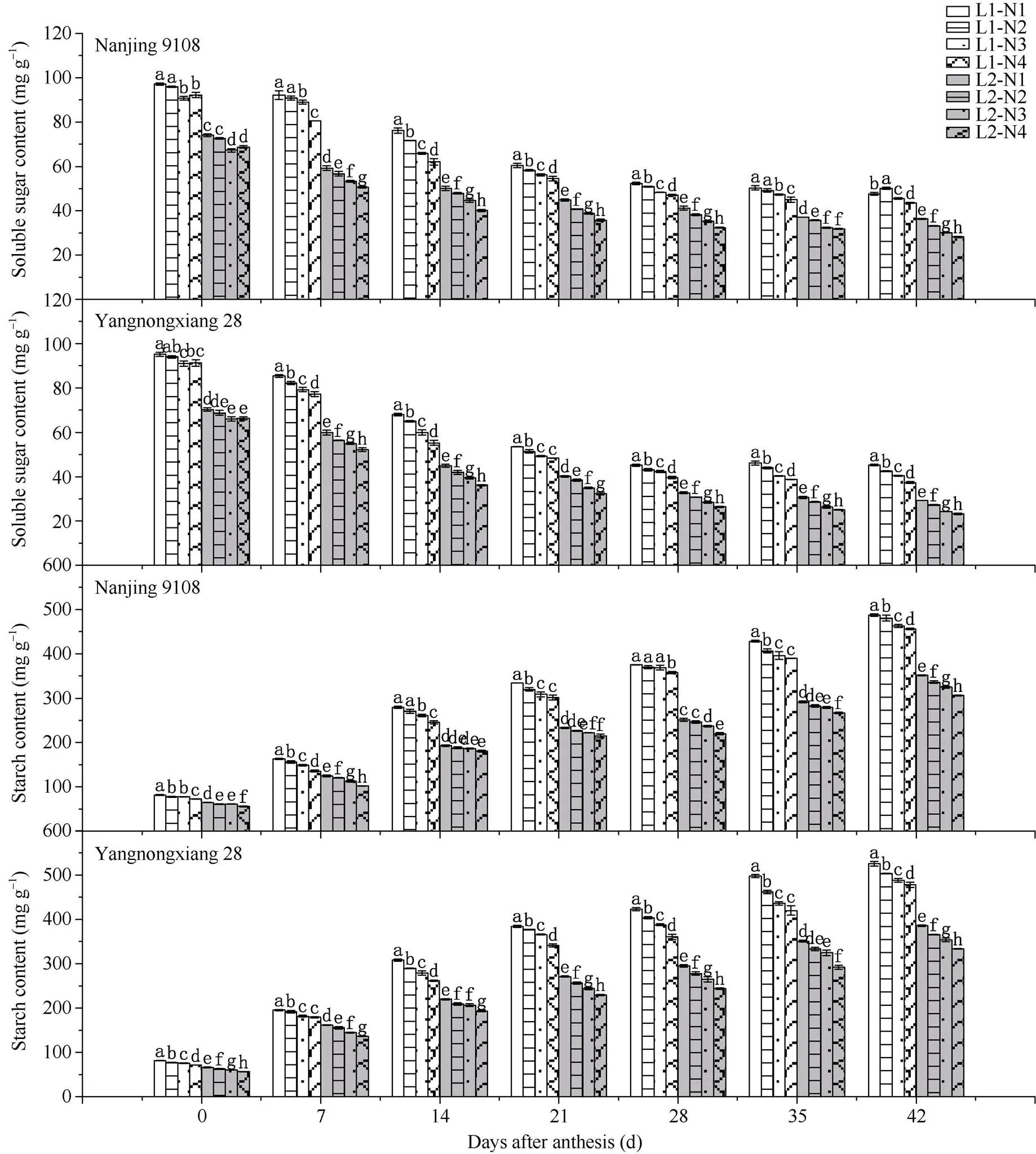
图3 不同光、氮条件下南方软米粳稻花后籽粒中可溶性糖和淀粉含量的差异
处理同图1。图中所标注字母为籽粒淀粉和可溶性糖含量的显著性, 不同字母表示在0.05概率水平差异显著。
Treatments are the same as those given in Fig. 1. Different lowercase letters indicate significantly different at the 0.05 probability level for the content of soluble sugar and starch in grains.
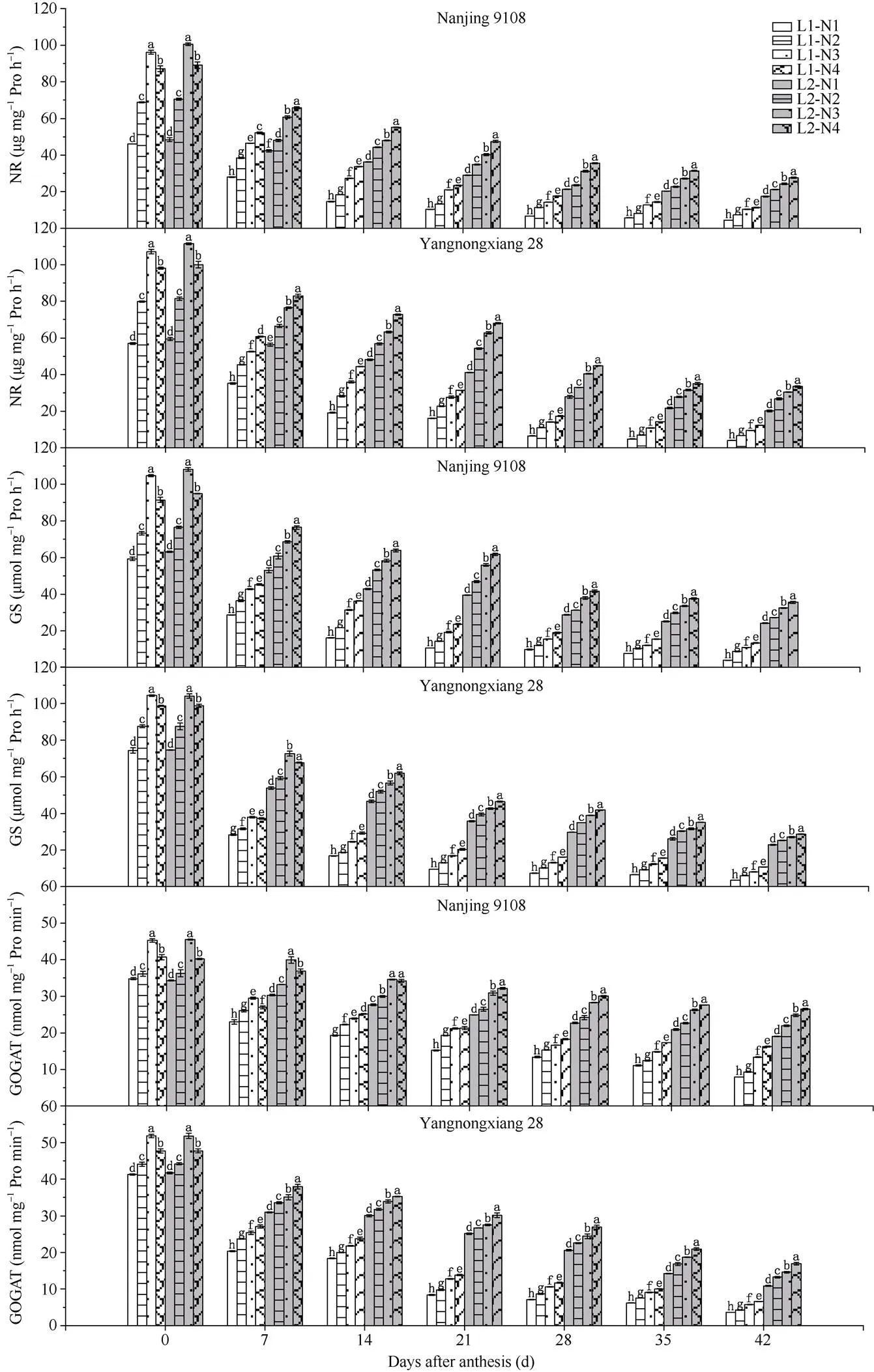
图4 不同光、氮条件下南方软米粳稻花后叶片中硝酸还原酶(NR)、谷氨酰胺合酶(GS)和谷氨酸合酶(GOGAT)的差异
处理同图1。图中所标注字母为硝酸还原酶、谷氨酰胺合酶和谷氨酸合酶的显著性, 不同字母表示在0.05概率水平差异显著。
Treatments are the same as those given in Fig. 1. Different lowercase letters indicate significantly different at the 0.05 probability level for activity of nitrate reductase, glutamine synthetase, and glutamate synthetase.
2.2.3 氮代谢相关酶 50%光强处理下供试2个品种叶片中氮代谢相关的硝酸还原酶(NR)、谷氨酰胺合酶(GS)、谷氨酸合酶(GOGAT) (EC 2.6.1.53)的活性均显著增强(图3)。中后期施用氮肥后, 2个品种叶片中NR、GS、GOGAT活性显著增强, 且随着中后期氮肥施用时期的推迟增幅逐渐增加。在光、氮共同处理下, 南粳9108和扬农香28的NR、GS、GOGAT活性在灌浆后期均表现为L2N4>L2N3> L2N2>L2N1>L1N4>L1N3>L1N2>L1N1。
2.2.4 叶片和籽粒的碳氮比 50%光强处理下水稻成熟期叶片和籽粒中的C/N均显著降低(图6), 供试2个品种的变化基本一致, 其中叶片中C/N下降40.43%~54.23%, 籽粒中C/N下降34.04%~38.16%。中后期施用氮肥后, 2个品种叶片和籽粒中的C/N下降, 下降幅度随着中后期氮肥施用时期的推迟逐渐增加, 与N1处理相比, N2、N3、N4处理下叶片中C/N分别降低了7.69%~13.45%、14.15%~22.66%、20.81%~26.31%, 籽粒中C/N分别降低了4.03%~ 5.39%、7.80%~10.89%、12.50%~14.55%。
2.3 不同光、氮条件对优质粳稻产量的影响
2.3.1 产量及产量结构 虽然50%光强处理下供试2个品种的每穗粒数和总颖花量无显著差异, 但结实率、千粒重显著下降, 产量降低(表7)。中后期施用氮肥后, 2个品种总颖花量和产量显著升高。随着中后期氮肥施用时期的推迟, 总颖花量呈现N3>N2>N4的趋势, 结实率在正常光照强度处理下随中后期氮肥施用时期的推迟逐渐上升, 产量则呈现先升后减的趋势, 而在50%光强处理下结实率和产量均随着中后期氮肥施用时期的推迟逐渐下降。光、氮及光氮互作对2个品种产量的影响均达极显著水平, 产量以L1N3处理最高, 遮光处理下, 产量随着中后期氮肥施用时期的推迟而逐渐下降, 表明中后期适当的提前施用氮肥可缓减光照不足导致的产量下降。光照强度对结实率和千粒重的影响达极显著水平, 氮素对产量构成因素均达极显著水平。
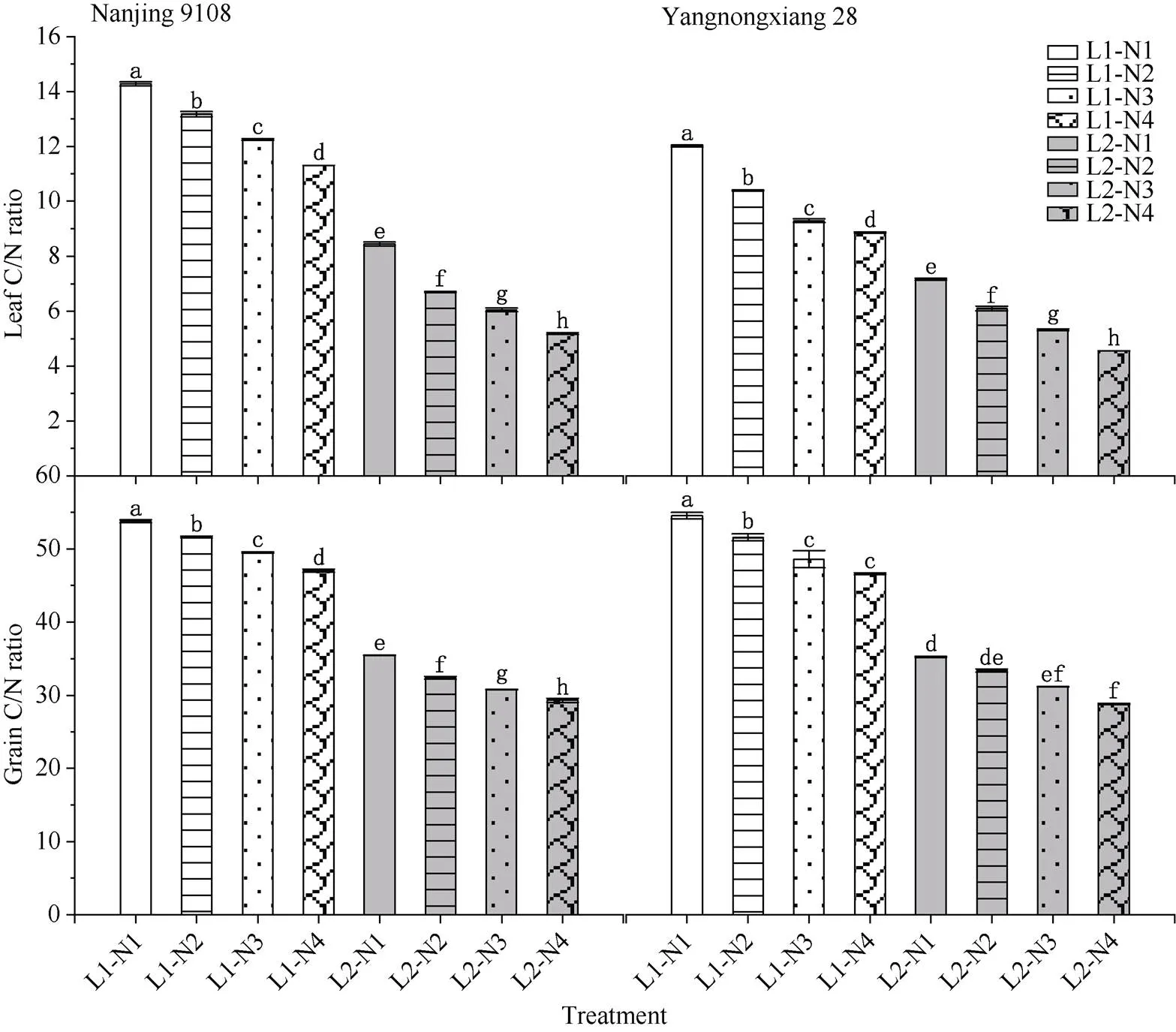
图5 不同光、氮条件下对南方软米粳稻叶片和籽粒C/N比的差异
处理同图1。图中所标注字母为叶片和籽粒中C/N的显著性, 不同字母表示在0.05概率水平差异显著。
Treatments are the same as those given in Fig. 1. Different lowercase letters indicate significantly different at the 0.05 probability level for C/N ratio of leaves and grains.
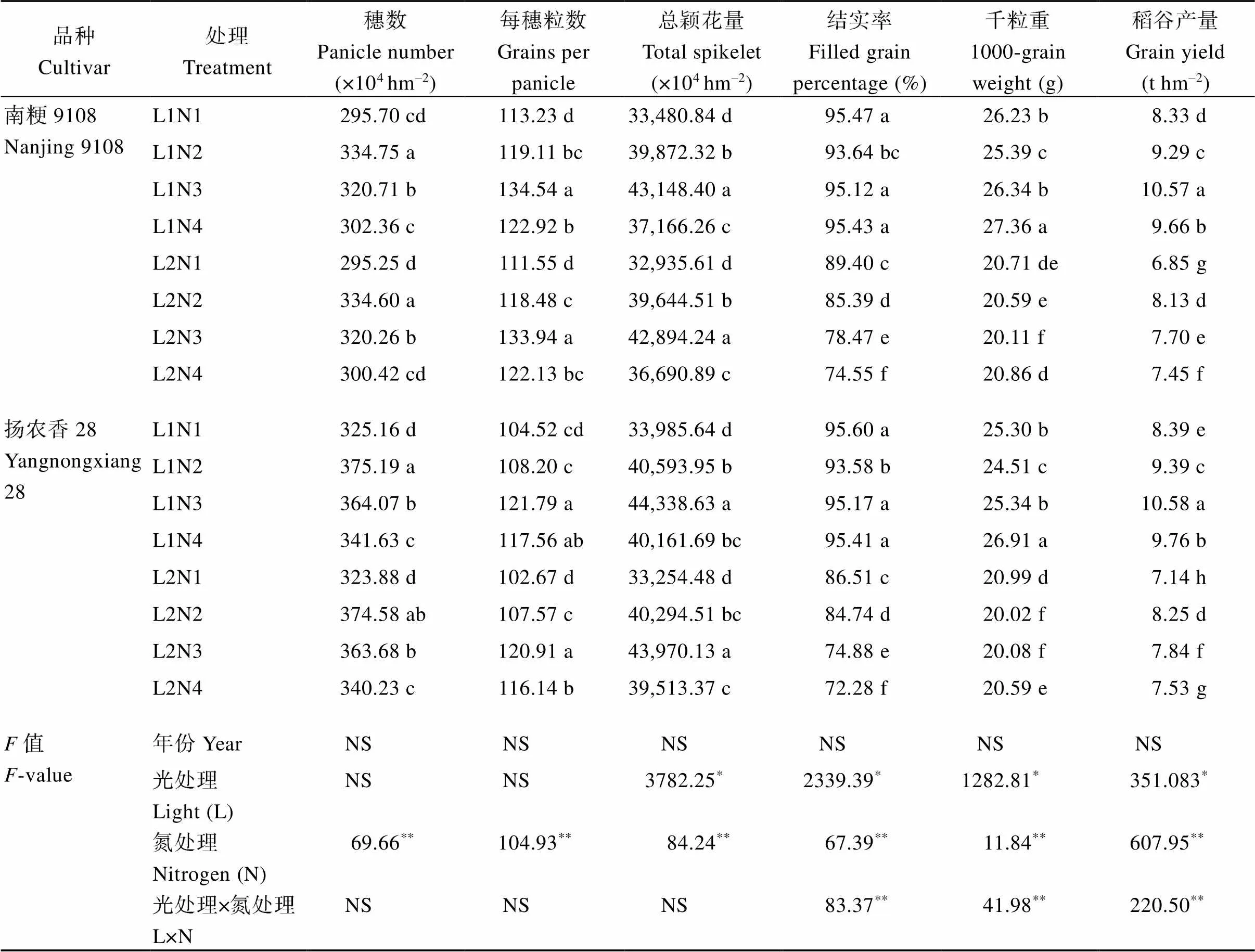
表7 不同光、氮条件下南方软米粳稻产量构成因素的差异
处理同图1。标以不同小写字母的值分别表示同一品种不同处理在0.05概率水平差异显著。*和**分别表示在0.05和0.01概率水平差异显著, NS表示差异不显著。
Treatments are the same as those given in Fig. 1. Different lowercase letters indicate that there are significant difference in the 0.05 probability level in different treatments of the same variety.*and**indicate significant differences at the 0.05 and 0.01 probability levels, respectively, and NS indicates no significant differences.
2.3.2 干物质积累 50%光强处理下供试2个品种抽穗期干物质积累无显著差异(表8), 成熟期及花后干物质积累量显著降低。中后期施用氮肥后, 抽穗期、成熟期及花后干物质的积累量均显著提高, 且随着中后期肥施用时期的推迟, 抽穗期的干物质积累量先增后减, 成熟期及花后干物质积累量在正常光照强度处理下随中后期氮肥施用时期的推迟先增后减, 而在50%光强处理下则逐渐减少。光、氮及光氮互作对2个品种抽穗期、成熟期及花后干物质的积累量的影响均达极显著水平。
2.4 不同光照、氮素条件对优质粳稻品质的影响
2.4.1 加工和外观品质 50%光强处理下水稻的糙米率、精米率、整精米率均显著降低(表9), 2个供试品种的变化基本一致, 降幅在2.08%~11.51%, 垩白粒率和垩白度均显著升高; 在中后期施用氮肥后, 糙米率、精米率、整精米率、垩白粒率和垩白度均显著升高, 且升幅随中后期氮肥施用时期的推迟逐渐增加; 在50%光强处理下中后期推迟施用氮肥, 能够缓解由于光照强度不足造成的糙米率、精米率、整精米率下降, 但同时增加了籽粒垩白粒率和垩白度。

表8 不同光、氮条件下南方软米粳稻干物质积累的差异
处理同图1。标以不同小写字母的值分别表示同一品种不同处理在0.05概率水平差异显著。*和**分别表示在0.05和0.01概率水平差异显著, NS表示差异不显著。
Treatments are the same as those given in Fig. 1. Different lowercase letters indicate that there are significant difference in the 0.05 probability level in different treatments of the same variety.*and**indicate significant differences at the 0.05 and 0.01 probability levels, respectively, and NS indicates no significant differences.
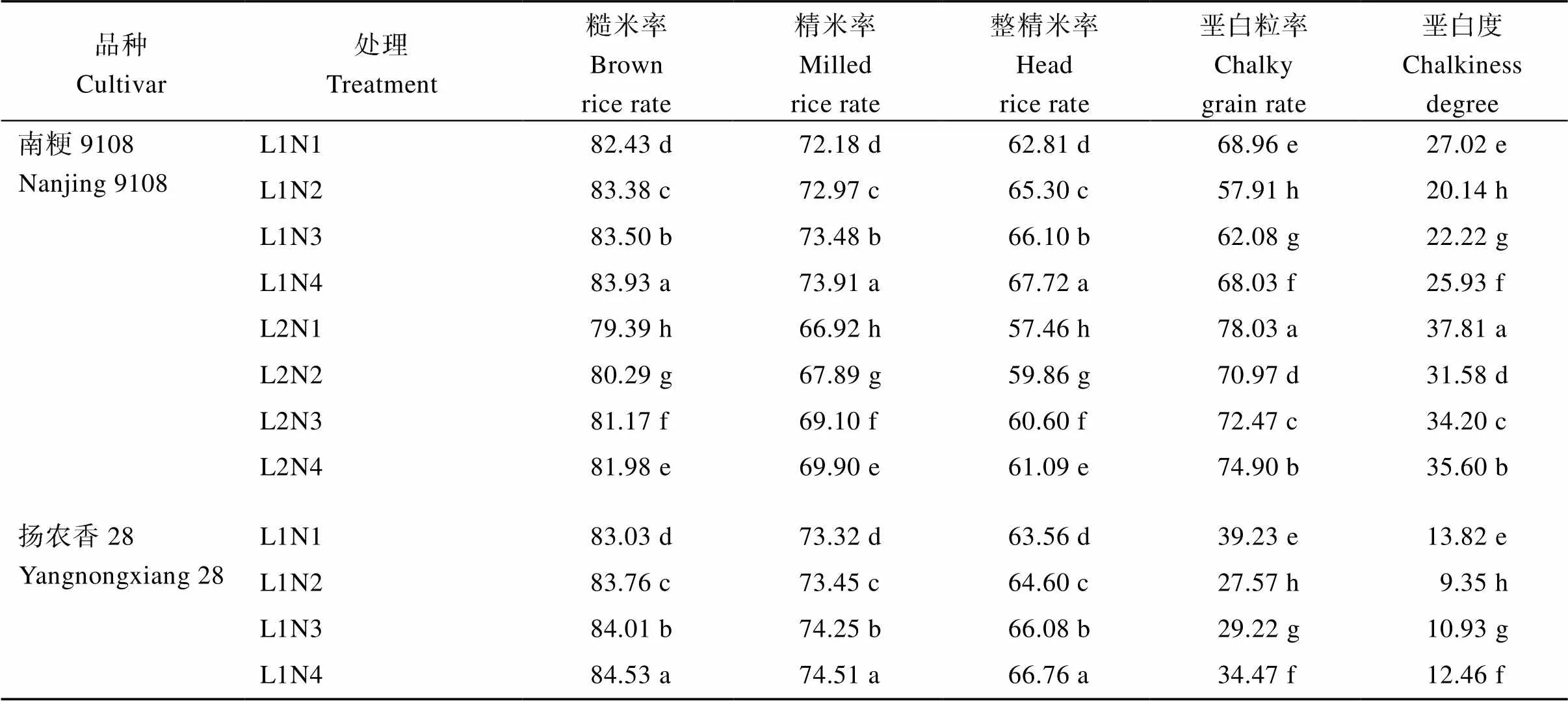
表9 不同光、氮条件对南方软米粳稻加工和外观品质的差异

(续表9)
处理同图1。标以不同小写字母的值分别表示同一品种不同处理在0.05概率水平差异显著。*和**分别表示在0.05和0.01概率水平差异显著, NS表示差异不显著。
Treatments are the same as those given in Fig. 1. Different lowercase letters indicate that there are significant difference in the 0.05 probability level in different treatments of the same variety.*and**indicate significant differences at the 0.05 and 0.01 probability levels, respectively, and NS indicates no significant differences.
2.4.2 营养和蒸煮食味品质 50%光强处理下供试2个品种的蛋白质含量(PC)均显著提高(表10), 总淀粉含量(TSC)、直链淀粉含量(AC)及AC/PC、TSC/PC均显著降低。中后期施用氮肥后, 2个品种的蛋白质含量显著提高, 总淀粉含量、直链淀粉含量、AC/PC及TSC/PC均显著下降, 随着中后期氮肥施用时期的推迟, 蛋白质含量的升幅、总淀粉含量、直链淀粉含量的降幅逐渐增加, AC/PC、TSC/PC逐渐降低。50%光强处理下中后期推迟施用氮肥与正常光照强度处理下相一致, 2个品种的蛋白质含量显著提高、总淀粉含量、直链淀粉含量、AC/PC及TSC/PC显著下降。
50%光强处理下供试2个品种的硬度较高, 但外观、黏度、平衡度以及食味值均较低。中后期施用氮肥后, 2个品种的硬度增加, 外观、黏度、平衡度以及食味值降低, 且随着中后期氮肥施用时期的推迟, 除硬度呈递增趋势外, 其余指标逐渐递减。在光、氮共同作用下, 食味值在L1N1处理下最优, 此时2个品种籽粒中AC/PC、TSC/PC均最高。
3 讨论
3.1 碳氮代谢对水稻产量的影响
光照强度影响水稻碳氮代谢进而影响产量。本研究中结实期遮光, 光强仅为自然光的50%, 水稻花后剑叶净光合速率下降7.35%~42.36%、同时叶片中蔗糖磷酸合酶(SPS)和蔗糖合酶(SS)酶活性下降, 碳代谢能力减弱, 不利于叶片中可溶性糖及淀粉合成, 导致叶片C/N比下降3.98~6.49, 水稻花后干物质生产能力大幅降低。水稻碳代谢减弱, 无法为叶片等营养器官中的氮素同化为氨基酸和蛋白质并向籽粒中转运的过程提供足够的碳骨架, 导致氮素在水稻各器官中的分配比例改变[30-31], 更多的氮素滞留于叶片[26,32], 来维持叶片中较高的硝酸还原酶(NR)、谷氨酰胺合酶(GS)、谷氨酸合酶(GOGAT)活性同时让叶片长期持绿[33-34], 保持较高的SPAD值, 以致于水稻贪青迟熟[18,30], 籽粒灌浆不充分, 籽粒C/N比下降17.44~19.28, 结实率和千粒重显著降低[35], 产量下降。
氮素在调节植株氮代谢的同时也在间接影响水稻碳代谢, 进而共同影响水稻产量。中后期施用氮肥能够促进根系的生长发育, 维持根系活力[36], 加快氮素吸收, 从而提升地上部植株及叶片的氮素含量。氮素能调控叶绿素合成与分解, 充足的氮素会减缓叶绿素等光合色素的分解[37], 延缓叶片的衰老, 延长水稻的灌浆结实期。同时中后期氮肥的施用使得植株绿叶面积增大、叶片中NR、GS、GOGAT酶活性增强, 同时叶片保持较高的SPAD值, 在正常光照条件下, 叶片的光合生产效率提升[38-40], 叶片中SPS、SS酶活性也随之增强。水稻分蘖的生长受碳氮代谢的直接影响, 碳代谢增强, 更多的糖类经韧皮部转运到分蘖芽中积累, 促进分蘖生长[38,41-42]。因此水稻群体在中后期施用氮肥后群体颖花量增加, 抽穗后光合生产持续稳定在较高水平, 加以灌浆结实期延长, 有更多的氮素和光合产物向穗部源源不断地输送[43-44], 籽粒灌浆充分[45], 产量增加。本研究中随着中后期施肥时期的推迟, 抽穗后剑叶净光合速率、叶绿素含量及碳氮代谢关键酶活性均逐渐升高, N3 (倒四叶施用氮肥)处理下, 叶片碳代谢能力增强, 促进颖花分化, 形成大穗, 拉动植株对氮素的吸收, 从而形成高产。
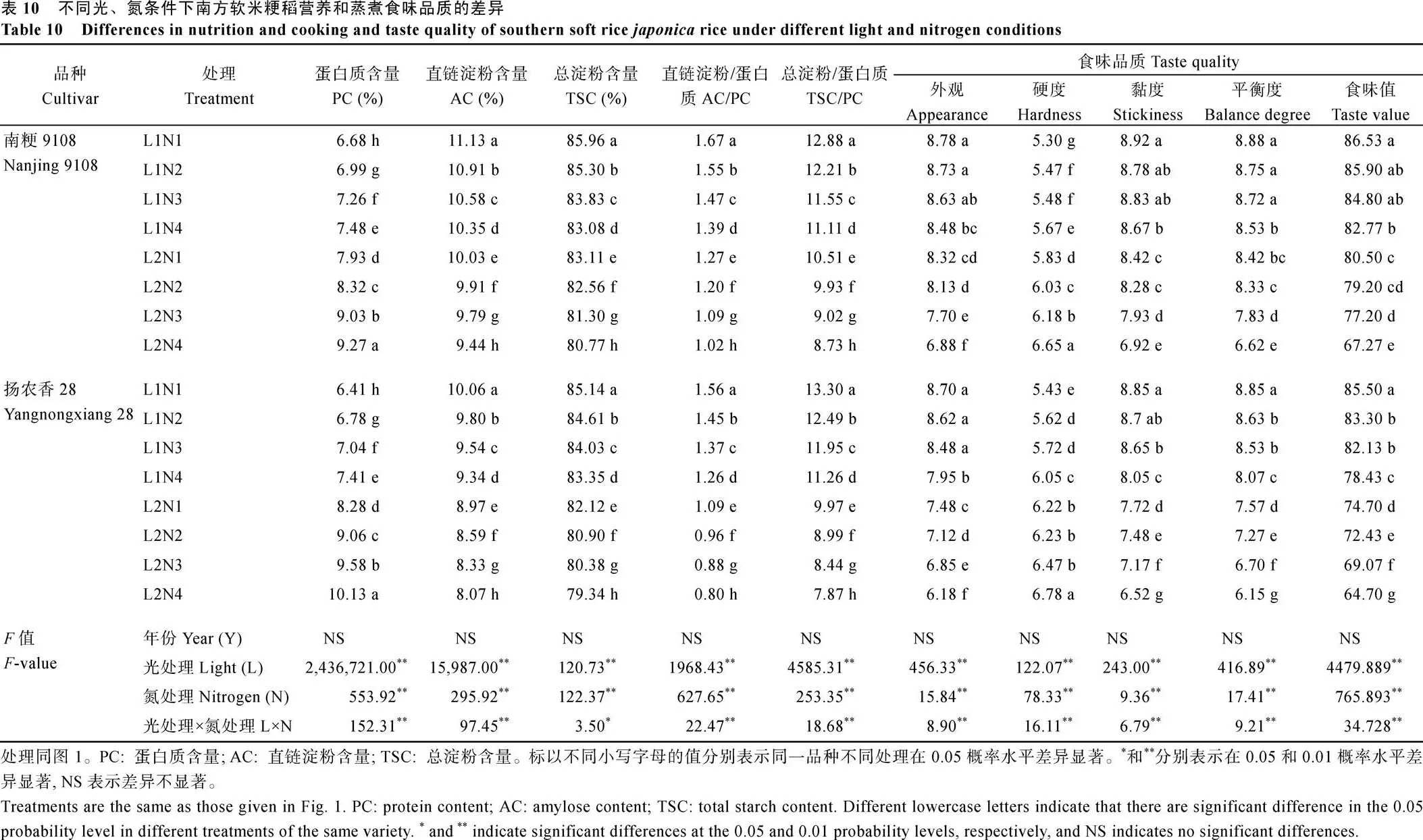
50%光强条件下中后期施用氮肥, 使得原本相对较高的氮代谢因为施用了氮肥而更加旺盛, 叶片C/N比进一步降低, 氮素在叶片中大量积累, 直至成熟期, 叶色因氮含量高而保持浓绿, 成熟期叶片与抽穗期叶片的SPAD比值相较于正常光照强度上升了0.16~0.17, 叶片中充足的氮素营养因缺少光照条件而不能充分用于促进光合产物的生产并向籽粒的输送而滞留于叶片, 难以实现结实阶段碳氮代谢平衡而获得高产。
3.2 碳氮代谢对水稻品质的影响
水稻源器官生产的光合同化物以蔗糖的形式经韧皮部运输到籽粒的过程中也伴随着植株体内包括功能叶片中的氮素转化为氨基酸向籽粒的协同输送,最终在籽粒中形成一定比例的碳氮代谢产物——淀粉和蛋白质, 从而决定稻米的品质[46-50]。50%光强条件下, 水稻叶片的净光合速率下降, 叶片中的光合产物减少, 同化物分配发生紊乱, 胚乳淀粉合成受限, 籽粒中淀粉含量和直链淀粉含量均降低[51], 淀粉体排列疏松, 间隙增大, 籽粒的垩白率和垩白度增加[24,51-52]。光强减弱后碳代谢的减弱伴随着氮代谢的相对增强, 本研究中遮光后光强仅为自然光强的50%, 且可利用光是以蓝紫光为主的散射光。蓝紫光能促进氮素的合成, 导致植株含氮量增加[32], 进而促进蛋白质的积累, 使得成熟期籽粒蛋白质含量上升, C/N比下降。过多的蛋白质与直链淀粉形成复合物, 增加了淀粉粒的刚性, 阻止大米在糊化过程中淀粉粒的膨胀和崩解, 促进淀粉回生, 最终导致米饭硬度增加, 黏度、平衡性降低, 食味品质变差[53-54]。
氮素在调节碳氮代谢的同时也调控稻米品质。中后期施用氮肥后, 叶片中氮代谢相较于碳代谢更加旺盛, 花后更多的含氮化合物转运至籽粒中, 籽粒C/N下降, 蛋白质含量相对增加[55]。蛋白质含量增加后更好的填充了因为淀粉充实不完整而造成的空隙[56], 胚乳结构愈加坚硬紧致, 抗压性增强, 加工和外观品质得到改善, 同时蛋白质含量增加, 限制稻米蒸煮过程中的吸水, 食味品质变差。随着中后期氮肥施用时期的推迟, 籽粒C/N进一步下降, 蛋白质和淀粉的比例发生改变, 籽粒中蛋白质含量更高, 淀粉和直链淀粉含量则下降趋势。碳氮代谢强度与碳氮代谢产物的积累密切相关[57], 适宜的碳氮代谢产物比例有助于形成良好的食味品质, 本研究中N2处理在中后期施用氮肥处理中籽粒C/N、淀粉/蛋白质及直链淀粉/蛋白质的比值最高, 食味品质最优。
50%光强条件下中后期施用氮肥, 碳代谢减弱的同时氮代谢相对旺盛, 籽粒中C/N进一步下降, 淀粉与蛋白质、直链淀粉与蛋白质的比值分别在7.87~9.93和0.80~1.20之间, 同正常光强条件施用氮肥相比分别下降了2.28~3.51、0.35~0.49, 米饭的硬度增加, 黏度、平衡性降低, 食味值降低[53]。
4 结论
水稻结实期平衡的碳氮代谢是水稻获得高产优质的基础。光强减弱, 叶片净光合速率和叶片中碳代谢相关酶活性下降, 不能为氮代谢的进行提供充足的碳源和能量, 中后期适期施用氮肥能为水稻的生长提供充足的氮素营养, 叶片延缓衰老, 叶片中氮代谢相关酶活性增强, 在提升水稻氮代谢的同时又能够进一步提高水稻叶片的净光合速率, 增强碳代谢能力, 促进籽粒中碳氮代谢产物的积累。本试验条件下, 正常光照强度配合倒四叶施用氮肥处理(L1~N3)能够协同提高叶片碳氮代谢关键酶活性, 使得光合产物和含氮化合物以适宜的比例向籽粒输送, 最终籽粒中淀粉与蛋白质的比值在11.55~11.95之间, 直链淀粉与蛋白质的比值在1.37~1.47之间, 米饭的硬度低, 黏度、平衡性高, 食味好, 可同时获得高产和优质。
[1] 李明启. 作物的“碳代谢”和“氮代谢”. 广东农业科学, 1981, (2): 12–13. Li M Q. “Carbon metabolism” and “nitrogen metabolism” of crops., 1981, (2): 12–13 (in Chinese).
[2] 陈雅玲, 包劲松. 水稻胚乳淀粉合成相关酶的结构、功能及其互作研究进展. 中国水稻科学, 2017, 31: 1–12. Chen Y L, Bao J S. Progress in structures, functions and interactions of starch synthesis related enzymes in rice endosperm., 2017, 31: 1–12 (in Chinese with English abstract).
[3] 张青, 孟杉杉, 陈梓春, 王远江, 韦存虚, 王娟. 谷物胚乳淀粉合成相关酶的调控机制研究进展. 植物生理学报, 2021, 57: 1–11. Zhang Q, Meng S S, Chen Z C, Wang Y J, Wei C X, Wang J. Progress of regulation mechanism of starch biosynthesis related enzymes in cereal endosperm., 2021, 57: 1–11 (in Chinese with English abstract).
[4] 韩展誉, 管弦悦, 赵倩, 吴春艳, 黄福灯, 潘刚, 程方民. 灌浆温度和氮肥及其互作效应对稻米贮藏蛋白组分的影响. 作物学报, 2020, 46: 1087–1098. Han Z Y, Guan X Y, Zhao Q, Wu C Y, Huang F D, Pan G, Cheng F M. Individual and combined effects of air temperature at filling stage and nitrogen application on storage protein accumulation and its different components in rice grains., 2020, 46: 1087–1098 (in Chinese with English abstract).
[5] 韩展誉, 吴春艳, 许艳秋, 黄福灯, 熊义勤, 管弦悦, 周庐建, 潘刚, 程方民. 不同施氮水平下灌浆期高温对水稻贮藏蛋白积累及其合成代谢影响. 中国农业科学, 2021, 54: 1439–1454. Han Z Y, Wu C Y, Xu Y Q, Huang F D, Xiong Y Q, Guan X Y, Zhou L J, Pan G, Cheng F M. Effects of high-temperature at filling stage on grain storage protein accumulation and its biosynthesis metabolism for rice plants under different nitrogen application levels., 2021, 54: 1439–1454 (in Chinese with English abstract).
[6] 王军可, 王亚梁, 陈惠哲, 向镜, 张义凯, 朱德峰, 张玉屏. 灌浆初期高温影响水稻籽粒碳氮代谢的机理. 中国农业气象, 2020, 41: 32–42. Wang J K, Wang Y L, Chen H Z, Xiang J, Zhang Y K, Zhu D F, Zhang Y P. Mechanism of high temperature affecting carbon and nitrogen metabolism of rice grain at the early stage of grain filling., 2020, 41: 32–428 (in Chinese with English abstract).
[7] 徐国伟, 吕强, 陆大克, 王贺正, 陈明灿. 干湿交替灌溉耦合施氮对水稻根系性状及籽粒库活性的影响. 作物学报, 2016, 42: 1495–1505. Xu G W, Lyu Q, Lu D K, Wang H Z, Chen M C. Effect of wetting and drying alternative irrigation coupling with nitrogen application on root characteristic and grain-sink activity., 2016, 42: 1495–1505 (in Chinese with English abstract).
[8] 吕腾飞, 谌洁, 代邹, 马鹏, 杨志远, 郑传刚, 马均. 缓释氮肥与尿素配施对机插杂交籼稻碳氮积累的影响. 作物学报, 2021, 47: 1966–1977. Lyu T F, Chen J, Dai Z, Ma P, Yang Z Y, Zheng C G, Ma J. Effects of combined application of slow release nitrogen fertilizer and urea on carbon and nitrogen accumulation in mechanical transplanted hybrid rice., 2021, 47: 1966–1977 (in Chinese with English abstract).
[9] Ding C, You J, Chen L, Wang S, Ding Y. Nitrogen fertilizer increases spikelet number per panicle by enhancing cytokinin synthesis in rice., 2014, 33: 363–371.
[10] 吕川根, 李霞, 陈国祥. 超级杂交稻两优培九高产的光合特性及其生理基础. 中国农业科学, 2017, 50: 4055–4071. Lyu C G, Li X, Chen G X. Photosynthetic characteristics and its physiological basis of super high-yielding hybrid rice Liangyoupeijiu., 2017, 50: 4055–4071 (in Chinese with English abstract).
[11] 孙永健, 孙园园, 严奉君, 杨志远, 徐徽, 李玥, 王海月, 马均. 氮肥后移对不同氮效率水稻花后碳氮代谢的影响. 作物学报, 2017, 43: 407–419. Sun Y J, Sun Y Y, Yan F J, Yang Z Y, Xu H, Li Y, Wang H Y, Ma J. Effects of postponing nitrogen topdressing on post-anthesis carbon and nitrogen metabolism in rice cultivars with different nitrogen use efficiencies., 2017, 43: 407–419 (in Chinese with English abstract).
[12] 从夕汉, 施伏芝, 阮新民, 罗玉祥, 王元垒, 许有尊, 罗志祥. 施氮量对不同品种水稻氮素利用及碳氮代谢关键酶的影响. 河南农业大学学报, 2019, 53: 325–330. Cong X H, Shi F Z, Wang Y L, Ruan X M, Xu Y Z, Luo Y X, Luo Z X. Effects of nitrogen application rate on nitrogen use efficiency and key enzymes for carbon and nitrogen metabolism in different rice varieties., 2019, 53: 325–330 (in Chinese with English abstract).
[13] 胡群, 夏敏, 张洪程, 曹利强, 郭保卫, 魏海燕, 陈厚存, 韩宝富. 氮肥运筹对钵苗机插优质食味水稻产量及品质的影响. 作物学报, 2017, 43: 420–431. Hu Q, Xia M, Zhang H C, Cao L Q, Guo B W, Wei H Y, Chen H C, Han B F. Effect of nitrogen application regime on yield and quality of mechanical pot-seedlings transplanting rice with good taste quality., 2017, 43: 420–431. (in Chinese with English abstract).
[14] 陶进, 钱希旸, 剧成欣, 刘立军, 张耗, 顾骏飞, 王志琴, 杨建昌. 不同年代中籼水稻品种的米质及其对氮肥的响应. 作物学报, 2016, 42: 1352–1362. Tao J, Qian X Y, Ju C X, Liu L J, Zhang H, Gu J F, Wang Z Q, Yang J C. Grain quality and its response to nitrogen fertilizer in mid-season indica rice varieties planted in different decades from 1950s to 2010s., 2016, 42: 1352–1362 (in Chinese with English abstract).
[15] Deng F, Li B, Yuan Y J, He C Y, Zhou X, Li Q P, Zhu Y Y, Huang X F, He Y X, Ai X F, Tao Y F, Zhou W, Wang L, Cheng H, Chen Y, Wang M T, Ren W J. Increasing the number of seedlings per hill with reduced number of hills improves rice grain quality by optimizing canopy structure and light utilization under shading stress., 2022, 287: 108668.
[16] Schmierer M, Knopf O, Asch F. Growth and photosynthesis responses of a super dwarf rice genotype to shade and nitrogen supply., 2021, 28: 178–190.
[17] 杜彦修, 季新, 张静, 李俊周, 孙红正, 赵全志. 弱光对水稻生长发育影响研究进展. 中国生态农业学报, 2013, 21: 1307–1317. Du Y X, Ji X, Zhang J, Li J Z, Sun H Z, Zhao Z Q. Research progress on the impacts of low light intensity on rice growth and development., 2013, 21: 1307–1317 (in Chinese with English abstract).
[18] 王亚江, 颜希亭, 孟天瑶, 杨大柳, 魏海燕, 张洪程, 戴其根, 霍中洋, 许轲. 抽穗前后遮光对超级粳稻产量形成的影响. 中国稻米, 2014, 20: 18–21. Wang Y J, Yan X T, Meng T Y, Yang D L, Wei H Y, Zhang H C, Dai Q G, Huo Z Y, Hu K. Effects of shading before and after heading on yield formation of superrice.2014, 20: 18–21 (in Chinese with English abstract).
[19] Gurevitch J, Howard T G, Ashton I W, Leger E A, Howe K M, Woo E, Lerdau M. Effects of experimental manipulation of light and nutrients on establishment of seedlings of native and invasive woody species in Long Island, NY forests., 2008, 10: 821–831.
[20] 姜楠. 遮光对北方粳稻产量和品质的形成及其生理机制的研究. 沈阳农业大学博士学位论文, 辽宁沈阳, 2013. Jiang N. Study on Development of Yield and Quality and Its Physiological Mechanism ofRice under Shading in Northern China. PhD Dissertation of Shenyang Agricultural University, Shenyang, Liaoning, China, 2013 (in Chinese with English abstract).
[21] 朱萍, 贺阳冬, 杨志远, 孙永健, 马均. 孕穗期和抽穗期弱光胁迫对2个杂交稻组合生理特性及产量的影响. 西南农业学报, 2014, 27: 559–566. Zhu P, He Y D, Yang Z Y, Sun Y J, Ma J. Effects of low-light stress at shooting and heading stage on some Photosynthetic characteristics and grain yield of two hybrid rice combinations., 2014, 27: 559–566 (in Chinese with English abstract).
[22] Elizabete C S A, Michael E S. The regulatory properties of Rubisco activase differ among species and affect photosynthetic induction during light transitions., 2013, 161: 4.
[23] 王亚江, 魏海燕, 颜希亭, 葛梦婕, 孟天瑶, 张洪程, 戴其根, 霍中洋, 许轲, 费新茹. 光、氮及其互作对超级粳稻产量和氮、磷、钾吸收的影响. 作物学报, 2014, 40: 1235–1244. Wang Y J, Wei H Y, Yan X T, Ge M J, Meng T Y, Zhang H C, Dai Q G, Huo Z Y, Xu Ke, Fei X R. Effects of light, nitrogen and their interaction on grain yield and nitrogen, phosphorus and potassium absorption insuper rice., 2014, 40: 1235–1244 (in Chinese with English abstract).
[24] 董明辉, 惠锋, 顾俊荣, 陈培峰, 杨代凤, 乔中英. 灌浆期不同光强对水稻不同粒位籽粒品质的影响. 中国生态农业学报, 2013, 21: 164–170. Dong M H, Hui F, Gu J R, Chen P F, Yang D F, Qiao Z Y. Effect of light intensity on grain quality of rice at different spike positions during grain-filling stage.,2013, 21: 164–170 (in Chinese with English abstract).
[25] 李林, 张更生. 阴害影响水稻产量的机制及其调控技术: II. 灌浆期模拟阴害影响水稻产量的机制. 中国农业气象, 1994, 15(3): 5–9 Li L, Zhang G S. Mechanism of insufficient illumination impact on rice yield and its controlling technology: II. Mechanism of impact of simulated insufficient illumination during the grain-filling period on rice yield.1994, 15(3): 5–9 (in Chinese with English abstract).
[26] 秦建权, 唐启源, 李迪秦, 张运波, 李小勇, 杨胜海, 陈立军. 抽穗后光照强度对超级杂交稻干物质生产及氮素吸收与分配的影响. 四川农业大学学报, 2010, 28: 28–34.Qin J Q, Tang Q, Li D Q, Zhang Y B, Li X Y, Yang S H, Chen L J. Effects of different light intensity after heading stage on dry matter accumulation and nitrogen uptake and distribution of super hybrid rice.2010, 28: 28–34 (in Chinese with English abstract).
[27] 刘奇华, 李天, 张建军. 生育前期遮光对水稻后期功能叶生长及稻米品质的影响. 生态学杂志, 2006, 10: 1167–1172. Liu Q H, Li T, Zhang J J. Effects of early stage shading on function leaf growth at grain-filling stage and on grain quality of rice., 2006, 25: 1167–1172 (in Chinese with English abstract).
[28] Mandal D, Tudu S, Mitra S R, De G C. Effect of common packing materials on keeping quality of sugarcane jaggery during monsoon season., 2006, 8: 137–142.
[29] 王志琴, 杨建昌, 朱庆森, 郎有忠. 水稻抽穗期茎鞘中储存的可用性糖与籽粒充实的关系. 江苏农学院学报, 1997, (4): 14–18. Wang Z Q, Yang J C, Zhu Q S, Lang Y Z. Relation of the usable carbohydrate reserved in stems and sheaths at heading stage with grain-filling in rice plantas., 1997, (4): 14–18 (in Chinese with English abstract).
[30] 黄丽芬, 全晓艳, 张蓉, 袁毅, 赵伟, 姜玲玲, 施金琦, 庄恒扬.光氮及其互作对水稻干物质积累与分配的影响. 中国水稻科学, 2014, 28: 167–176. Huang L F, Quan X Y, Zhang R, Yuan Y, Zhao W, Jiang L L, Shi J Q, Zhuang H Y. Interactive effects of light intensity and nitrogen supply on dry matter production and distribution of rice.2014, 28: 167–176 (in Chinese with English abstract).
[31] 秦建权, 唐启源, 李迪秦, 张运波, 李小勇, 杨胜海, 陈立军. 抽穗后光照强度对超级杂交稻干物质生产及氮素吸收与分配的影响. 四川农业大学学报, 2010, 28: 28–34. Qin J Q, Tang Q, Li D Q, Zhang Y B, Li X Y, Yang S H, Chen L J. Effects of different light intensity after heading stage on dry matter accumulation and nitrogen uptake and distribution of super hybrid rice.2010, 28: 28–34 (in Chinese with English abstract).
[32] 任万军, 杨文钰, 张国珍, 朱霞, 樊高琼, 徐精文. 弱光对杂交稻氮素积累、分配与子粒蛋白质含量的影响. 植物营养与肥料学报, 2003, 9: 288. Ren W J, Yang W Y, Zhang G Z, Zhu X, Fan G Q, Xu J W. Effect of low-light stress on nitrogen accumulation distribution and grains protein content of Indica hybrid., 2003, 9: 288 (in Chinese with English abstract).
[33] 方笑堃, 陈志炜, 程兆康, 姜海波, 邱丹, 罗小三. 太阳辐射减弱对水稻光合生理特性和中微量元素积累的影响. 应用生态学报, 2021, 32: 1345–1351. Fang X K, Chen Z W, Cheng Z K, Jiang H B, Qiu D, Luo X S. Effects of reduced solar radiation on photosynthetic physiological characteristics and accumulation of secondary and micro elements in paddy rice., 2021, 32: 1345–1351 (in Chinese with English abstract).
[34] Liang Y P, Wang J Y, Zeng F L, Wang Q, Zhu L, Li H Y, Guo N H, Chen H W. Photorespiration regulates carbon-nitrogen metabolism by magnesium chelatase D subunit in rice., 2020, 69: 112–125.
[35] Wei H Y, Zhu Y, Qiu S, Han C, Hu L, Xu D, Zhou N B, Xing Z P, Hu Y J, Cui P Y, Dai Q G, Zhang H C. Combined effect of shading time and nitrogen level on grain filling and grain quality insuper rice., 2018, 17: 41–53.
[36] 徐冉, 陈松, 徐春梅, 刘元辉, 章秀福, 王丹英, 褚光. 施氮量对籼粳杂交稻甬优1540产量和氮肥利用效率的影响及其机制.作物学报, 2023, 49: 1630–1642. Xu R, Chen S, Xu C M, Liu Y H, Zhang X F, Wang D Y, Chu G. Effects of nitrogen fertilizer rates on grain yield and nitrogen use efficiency of-hybrid rice cultivar Yongyou-1540 and its physiological bases., 2023, 49: 1630–1642 (in Chinese with English abstract).
[37] 侯红乾, 林洪鑫, 刘秀梅, 冀建华, 刘益仁, 蓝贤瑾, 吕真真, 周卫军. 长期施肥处理对双季晚稻叶绿素荧光特征及籽粒产量的影响. 作物学报, 2020, 46: 280–289. Hou H Q, Lin H X, Liu X M, Ji J H, Liu Y R, Lan X J, Lyu Z Z, Zhou W J. Influence of long-term fertilizer application on chlorophyll fluorescence characteristics and grain yield of double cropping late rice., 2020, 46: 280–289 (in Chinese with English abstract).
[38] 王琰, 陈志雄, 姜大刚, 张灿奎, 查满荣. 增强叶片氮素输出对水稻分蘖和碳代谢的影响. 作物学报, 2022, 48: 739–746. Wang Y, Chen Z X, Jiang D G, Zhang C K, Zha M R. Effects of enhancing leaf nitrogen output on tiller growth and carbon metabolism in rice., 2022, 48: 739–746 (in Chinese with English abstract).
[39] 李睿, 娄运生, 张震, 马莉, 李君. 节水灌溉和遮光强度对水稻生长发育的耦合影响. 中国农业气象, 2018, 39: 702–714. Li R, Lou Y S, Zhang Z, Ma L, Li J. Coupling effects of water-saving irrigation and shading intensity on growth and development for rice., 2018, 39: 702–714 (in Chinese with English abstract).
[40] Mu X H, Chen Y L. The physiological response of photosynthesis to nitrogen deficiency., 2021, 158: 76–82.
[41] François B, Thomas P, Marion L, Maria-Dolores P G, Quentin B, Jakub R, Stéphanie B M, Sylvie C, Remi L, Benoît P, Hanaé R, Nathalie L, José L G, Jessica B, Soulaiman S. Sucrose is an early modulator of the key hormonal mechanisms controlling bud outgrowth in Rosa hybrida., 2015, 66: 9.
[42] Wang F, Han T W, Song Q X, Ye W X, Song X G, Chu J F, Li J Y, Chen Z J. The rice circadian clock regulates tiller growth and panicle development through strigolactone signaling and sugar sensing.,2020, 32: 3124–3128.
[43] 董桂春, 于小凤, 赵江宁, 居静, 田昊, 李进前, 张燕, 王余龙.不同穗型常规籼稻品种氮素吸收利用的基本特点. 作物学报, 2009, 35: 2091–2100. Dong G C, Yu X F, Zhao J N, Ju J, Tian H, Li J Q, Zhang Y, Wang Y L. General characteristics of nitrogen uptake and utilization in conventional indica rice cultivars with different panicle weight types., 2009, 35: 2091–2100 (in Chinese with English abstract).
[44] 魏海燕, 张洪程, 杭杰, 戴其根, 霍中洋, 许轲, 张胜飞, 马群, 张庆, 张军. 不同氮素利用效率基因型水稻氮素积累与转移的特性. 作物学报, 2008, 34: 119–125. Wei H Y, Zhang H C, Hang J, Dai Q G, Huo Z Y, Xu K, Zhang S F, Ma Q, Zhang Q, Zhang J. Characteristics of N accumulation and translocation in rice genotypes with different N use efficiencies., 2008, 34: 119–125 (in Chinese with English abstract).
[45] Zhang J, Zhang Y Y, Song N Y, Chen Q L, Sun H Z, Peng T, Huang S, Zhao Q Z. Response of grain-filling rate and grain quality of mid-seasonrice to nitrogen application., 2021, 20: 1465–1473.
[46] 黄发松, 孙宗修, 胡培松, 唐绍清. 食用稻米品质形成研究的现状与展望. 中国水稻科学, 1998, 12: 172–176. Huang F S, Sun Z X, Hu P S, Tang S Q. Present situations and prospects for the research on rice grain quality forming., 1998, 12: 172–176 (in Chinese with English abstract).
[47] 任万军, 杨文钰, 徐精文, 樊高琼, 马周华. 弱光对水稻籽粒生长及品质的影响. 作物学报, 2003, 29: 785–790. Ren W J, Yang W Y, Xu J W, Fan G Q, Ma Z H. Effect of low light on grains growth and quality in rice., 2003, 29: 785–790 (in Chinese with English abstract).
[48] 陈莹莹, 胡星星, 陈京都, 杨雄, 马群, 陈乔, 葛梦婕, 戴其根. 氮肥水平对江苏早熟晚粳稻食味品质的影响及其品种间差异. 作物学报, 2012, 38: 2086–2092. Chen Y Y, Hu X X, Chen J D, Yang X, Ma Q, Chen Q, Ge M J, Dai Q G. Effect of nitrogen fertilizer application on eating quality of early-maturing laterice in Jiangsu and its difference among varieties., 2012, 38: 2086–2092 (in Chinese with English abstract).
[49] 张诚信, 郭保卫, 唐健, 许方甫, 许轲, 胡雅杰, 邢志鹏, 张洪程, 戴其根, 霍中洋, 魏海燕, 黄丽芬, 陆阳, 唐闯, 戴琪星, 周苗, 孙君仪. 灌浆结实期低温弱光复合胁迫对稻米品质的影响. 作物学报, 2019, 45: 1208–1220. Zhang C X, Guo B W, Tang J, Xu F F, Xu K, Hu Y J, Xing Z P, Zhang H C, Dai Q G, Huo Z Y, Wei H Y, Huang L F, Lu Y, Tang C, Dai Q X, Zhou M, Sun J Y. Combined effects of low temperature and weak light at grain-filling stage on rice grain quality., 2019, 45: 1208–1220 (in Chinese with English abstract).
[50] Hu W X, Chen J, Xu F, Chen L, Zhao J W. Study on crystalline, gelatinization and rheological properties ofrice flour as affected by starch fine structure., 2020, 148:1232–1241.
[51] Deng F, Li Q P, Chen H, Zeng Y L, Li B, Zhong X Y, Wang L, Ren W J. Relationship between chalkiness and the structural and thermal properties of rice starch after shading during grain-filling stage., 2021, 252: 117212.
[52] Dutta S S, Tyagi W, Rai M. Physiological and molecular response to low light intensity in rice: a review., 2017, 38: 209–215.
[53] Zhang L X, Zhang C Q, Yan Y, Hu Z J, Wang K, Zhou J H, Zhou Y, Cao L M, Wu S J. Influence of starch fine structure and storage proteins on the eating quality of rice varieties with similar amylose contents., 2020, 101: 101.
[54] 石吕, 张新月, 孙惠艳, 曹先梅, 刘建, 张祖建. 不同类型水稻品种稻米蛋白质含量与蒸煮食味品质的关系及后期氮肥的效应. 中国水稻科学, 2019, 33: 541–552. Shi L, Zhang X Y, Sun H Y, Cao X M, Liu J, Zhang Z J. Relationship of grain protein content with cooking and eating quality as affected by nitrogen fertilizer at late growth stage for different types of rice varieties., 2019, 33: 541–552.
[55] Xu Y J, Zhang W Y, Jian C Q, Wang W L, Li K, Tian Y F, Wang Z Q, Zhu K K, Chang Y J. The role of polyamines in regulating amino acid biosynthesis in rice grains., 2021, 10: 1002.
[56] Noori Z, Qarluq A G, Kakar K, Abdiani S, Nawakht N A. Structural features of white-belly and white-core rice endosperm under scanning electron microscopic observation., 2022, 11: 615–620.
[57] 朱盈. 长三角地区软米粳稻品质特征及优质丰产氮高效协同的碳氮代谢机理. 扬州大学博士学位论文, 江苏扬州, 2022. Zhu Y. Quality Characteristics and Carbon and Nitrogen Metabolism Mechanism of Good Quality High Yield and High Nitrogen Efficiency of SoftRice in Yangtze River Delta. PhD Dissertation of Yangzhou University, Yangzhou, Jiangsu, China, 2022 (in Chinese with English abstract).
Effects of light intensity and nitrogen fertilizer interaction on carbon and nitrogen metabolism at grain-filling stage and its relationship with yield and quality of southern softrice
CHEN Xin-Yi, ZHU Ying, MA Zhong-Tao, ZHANG Ming-Yue, WEI Hai-Yan*, ZHANG Hong-Cheng, LIU Guo-Dong, HU Qun, LI Guang-Yan, and XU Fang-Fu
Jiangsu Key Laboratory of Crop Genetics and Physiology / Innovation Center of Rice Cultivation Technology in Yangtze Valley, Ministry of Agriculture / Co-Innovation Center for Modern Production Technology of Grain Crops, Yangzhou University, Yangzhou 225009, Jiangsu, China
Southern softrice Nanjing 9108 and Yangnongxiang 28 were selected as the experimental materials, and two light intensity treatments, and four nitrogen treatments were set. Light intensity treatments [100% natural light intensity (L1) and 50% natural light intensity (L2)] and four nitrogen treatments [no nitrogen fertilizer (N1) in the middle and late growth stages, one-time nitrogen fertilizer applied at the top sixth leaf stage (N2), one-time nitrogen fertilizer applied at the top fourth leaf stage (N3), and one-time nitrogen fertilizer applied at the top second leaf stage (N4)] were conducted at grain-filling stage. The difference of carbon and nitrogen metabolism at grain-filling stage and its effects on rice yield and quality under the conditions of different light intensity and nitrogen application period as well as light-nitrogen interaction conditions were investigated. The results showed that with the decrease of light intensity at grain-filling stage the net photosynthetic rate of flag leaf decreased by 7.35%–42.36% on average, sucrose phosphate synthase (SPS), and sucrose synthase (SS) had low activity, the C/N ratio of leaves decreased by 3.98–6.49, the transportation of photosynthetic products to grains decreased, and the content of grain starch (including amylose) decreased. Meanwhile, the activities of nitrate reductase (NR), glutamine synthetase (GS), and glutamate synthetase (GOGAT) increased, plant nitrogen concentration increased, and the accumulation of protein increased relatively, which were not conducive to the formation of yield and good quality. After the application of nitrogen fertilizer at the middle and late growth stages, the activities of key enzyme in carbon and nitrogen metabolism in leaves were significantly increased, the aging of leaves was slowed down, and the grain-filling period of rice was prolonged, which were conducive to the increase of yield. With the delay of nitrogen fertilizer application period, nitrogen metabolism became more vigorous, and the protein content in grain had a relative significant increase, resulting in the decrease of the ratio of starch to protein and the ratio of amylose to protein, and the decrease of taste value. Under the experimental condition, normal light intensity combined with nitrogen fertilizer treatment (L1–N3) at the top fourth leaf stage synergistically improved the activities of key enzymes of carbon and nitrogen metabolism in leaves, thus the photosynthetic products and nitrogen-containing compounds were transported to grains in the appropriate proportions. Ultimately, the ratio of starch to protein in grain ranged from 11.43 to 12.03, and the ratio of amylose to protein ranged from 1.34 to 1.50, the rice had low hardness, high viscosity, and balance as well as good taste, high yield, and excellent quality could be obtained simultaneously.
rice; light; nitrogen fertilizer; carbon and nitrogen metabolism; yield; rice quality
10.3724/SP.J.1006.2023.22054
本研究由国家自然科学基金项目(31971841), 财政部和农业农村部国家现代农业产业技术体系建设专项(水稻, CARS-01), 山东省重点研发计划课题项目(2021LZGC020-03), 国家重点研发计划项目(2022YFD2301401)和江苏高校优势学科建设工程项目(PAPD)资助。
This study was supported by the National Natural Science Foundation of China (31971841), the China Agriculture Research System of MOF and MARA (Rice, CARS-01), the Key Research Program of Shandong Province, China (2021LZGC020-03), the National Key Research and Development Program of China (2022YFD2301401), and the Priority Subject Program Development of Jiangsu Higher Education Institutions (PAPD).
魏海燕, E-mail: wei_haiyan@163.com, Tel: 0514-87974595
E-mail: 2215502977@qq.com
2022-09-25;
2023-05-24;
2023-05-31.
URL:https://kns.cnki.net/kcms2/detail/11.1809.S.20230530.1505.004.html
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
猜你喜欢
今日农业(2021年15期)2021-10-14 08:20:44
粮油食品科技(2020年6期)2020-12-08 03:26:36
农民致富之友(2020年32期)2020-12-03 13:49:21
红楼梦学刊(2020年2期)2020-02-06 06:14:32
天然产物研究与开发(2018年10期)2018-11-06 07:43:46
现代装饰(2018年4期)2018-05-22 02:57:11
河南农业(2017年7期)2017-07-25 09:22:05
浙江农业科学(2016年11期)2016-05-04 04:16:39
现代检验医学杂志(2014年3期)2014-02-02 02:42:24
中国神经精神疾病杂志(2013年1期)2013-03-11 20:23:37
