
毕赤酵母发酵耦合多糖酶解一步法制备结冷胶寡糖
2023-09-13 01:49李宏雨施旭詹晓北朱莉蒋芸高敏杰
食品与发酵工业 2023年17期
李宏雨,施旭,詹晓北,朱莉,蒋芸,高敏杰
(江南大学 生物工程学院,江苏 无锡,214122)
结冷胶是鞘氨单胞菌产生的一种胞外多糖,它是由→3)-β-D-葡萄糖-(1→4)-β-D-葡萄糖醛酸-(1→4)-β-D-葡萄糖-(1→4)-α-L-鼠李糖-(1→为重复结构组成的线性阴离子杂多糖[1]。结冷胶具有良好的安全性、凝胶性、互溶性、稳定性等优越的性能,不仅在食品领域,还在生物医药、化工、环境等行业得到广泛的应用[2-4]。
目前,低聚糖的功能活性已引起广泛关注。例如,已有研究报道了α-1,3-葡寡糖和卡拉胶低聚糖具有抗肿瘤活性[5-6],壳寡糖表现出了植物的抗病性[7]。具有不同结构性质的寡糖可能具有不同的生物活性,其中聚合度被认为是对分子功能特性影响最大的参数之一。特定聚合度寡糖可能具有特定的功能活性,如麦芽七糖可以作为蛋白质和细胞的特异性识别标记[8]。此外,一些寡糖的功能活性与聚合度大小呈现了相关性,如壳寡糖对超氧自由基的清除能力也与聚合度大小成正相关[9]。近年研究者发现利用酸水解结冷胶得到的低聚结冷胶具有植物的抗菌活性[10],并且能够促进植物生长和改善植物叶片品质[11],结冷胶寡糖也被发现是一种潜在的益生元[12]。大多数关于结冷胶寡糖的应用和生物活性的研究主要是基于混合形式的,因此降解结冷胶制备特定聚合度结冷胶寡糖具有重要意义。传统的低聚糖生产方法多是对多糖进行化学降解和酶法降解。化学降解被认为是多糖降解的经典方法,化学降解具有操作简便、成本低等优点[13],但化学降解对环境污染大且得到的产物组分往往比较复杂,阻碍了后续的分离纯化。酶法降解因为其特异性高而能得到均一的目标产物,且反应温和得到了越来越多的关注。酶法降解可以有效克服化学降解产物成分复杂的缺点,简化特定聚合度结冷胶寡糖生产的过程,提高结冷胶寡糖定向生产的实用性。
本研究利用前期实验室构建的含有结冷胶裂解酶基因的重组毕赤酵母进行外源表达,得到结冷胶裂解酶,并开发了毕赤酵母发酵耦合多糖酶解一步法制备单一聚合度结冷胶寡糖的方法——在结冷胶裂解酶发酵初始添加结冷胶,以达到生产结冷胶裂解酶并实时降解多糖,实现一步制备结冷胶寡糖的目的,并将酶解制备的结冷胶寡糖与酸解结冷胶寡糖加以对比,分别利用傅里叶变换红外光谱(Fourier transform infrared spectrometer, FTIR)与基质辅助激光解析电离飞行时间质谱(matrix assisted laser desorption ionization time of flight mass spectrometry,MALDI-TOF-MS)来解析2种产物在结构与分子质量方面的差异。以期为制备单一聚合度寡糖,进一步研究特定聚合度结冷胶寡糖功能活性提供理论依据。
1 材料与方法
1.1 试剂与仪器
低酰基结冷胶(食品级)购自江苏仟泊生物工程有限公司;蛋白胨、酵母粉、甘油和无水甲醇等购自国药集团化学试剂有限公司。
RE-5205型旋转蒸发器,上海亚荣生化仪器厂;3K15型高速冷冻离心机,德国Sigma公司; ultrafleXtreme型质谱仪,美国Bruker Daltonics公司;MULTISKAN FC型酶标仪,美国Thermo Fisher Scientific公司;NEXUS型傅里叶变换红外光谱仪,美国Nicolet公司。
1.2 培养基
酵母浸出粉胨葡萄糖培养基(yeast extract peptone dextrose medium, YPD)(g/L):酵母粉 10,蛋白胨 20,葡萄糖 20(固体培养基加入琼脂粉20)。
甘油缓冲复合培养基(buffered glycerol-complex medium, BMGY)(g/L):酵母粉 10,蛋白胨 20, YNB 13.4,生物素 4×10-5,甘油 10,K3PO4缓冲液浓度100 mmol/L,pH 6.0。
诱导表达培养基(buffered methanol-complex medium, BMMY)(g/L):酵母粉 10,蛋白胨 20,YNB 13.4,生物素 4×10-5,甲醇 10 mL/L,K3PO4缓冲液浓度100 mmol/L,pH 6.0。
1.3 摇瓶发酵方法
将前期实验室构建的含有结冷胶裂解酶基因的重组毕赤酵母划线接种于YPD平板培养基上,在30 ℃恒温培养箱中倒置活化60~72 h。活化结束后,挑取单菌落接种至BMGY液体培养基中(装液量10%,体积分数),30 ℃、200 r/min培养16~18 h至OD600值达到4~6,得到种子液。4 ℃下无菌收集种子液菌体接种至BMMY液体培养基(装液量20%,体积分数)中,30 ℃、200 r/min培养3~4 d发酵结束,每24 h补加体积分数1.0%的甲醇。
1.4 摇瓶发酵条件优化
按1.3节中所述方法获得种子液。设置初始发酵条件:酵母粉添加量20 g/L,甲醇添加量1%,pH 6.0。保持其他条件不变,调整发酵初始酵母粉添加量为0、5、10、15、20、25、30、35、40 g/L研究发酵初始酵母粉添加量对毕赤酵母生长、结冷胶裂解酶酶活力的影响。以同样的方法研究不同pH(3.5、4、4.5、5、5.5、6、6.5、7、7.5)、不同甲醇添加量(0、0.5%、1.0%、1.5%、2.0%、2.5%、3.0%、3.5%、4.0%,体积分数)对毕赤酵母生长、结冷胶裂解酶酶活力的影响,以寻求重组毕赤酵母的最佳发酵条件。
1.5 毕赤酵母发酵耦合多糖酶解一步法制备结冷胶寡糖及条件优化
称取适量结冷胶添加至BMMY液体培养基中,收集按1.3节中所述方法获得的种子液中的菌体接种至添加了结冷胶的BMMY培养基中,30 ℃、200 r/min培养至发酵结束。保持其他条件不变,调整发酵初始结冷胶添加量为0、0.5、1.0、1.5、2.0、2.5、3.0 g/L,研究发酵初始结冷胶添加量对结冷胶寡糖产量的影响。以初始结冷胶添加量2%,其他条件不变,研究发酵时间对结冷胶寡糖产量的影响。
1.6 酸水解结冷胶条件优化
1.6.1 底物浓度优化
分别称取1、3、5、7 g/L的食品级低酰基结冷胶溶于适量蒸馏水中,90 ℃水浴加热至结冷胶完全溶解,补加蒸馏水与浓盐酸至终溶液中HCl浓度为0.5 mol/L。将配制好的水解液置于70 ℃水浴锅中加热水解3 h。水解结束后分别取样待后续薄层色谱分析。
1.6.2 HCl浓度优化
称取4份5 g/L的食品级低酰基结冷胶溶于适量蒸馏水中,90 ℃水浴加热至结冷胶完全溶解,补加蒸馏水与浓盐酸至终溶液中的HCl浓度分别为0.25、0.5、0.75、1 mol/L。将配制好的水解液置于70 ℃水浴锅中加热水解3 h。水解结束后分别取样待后续薄层色谱分析。
1.6.3 水解时间优化
称取5 g/L的食品级低酰基结冷胶溶于适量蒸馏水中,90 ℃水浴加热至结冷胶完全溶解,补加蒸馏水与浓盐酸至终溶液中的HCl浓度为0.5 mol/L。将配制好的水解液置于70 ℃水浴锅加热水解6 h,每小时取样1次待后续薄层色谱分析。
1.7 结冷胶寡糖纯化方法
1.7.1 酸解结冷胶寡糖纯化方法
酸水解结束后加入NaOH将水解液pH调至7,然后离心除去未水解的悬浮物,上清液旋转蒸发浓缩至水解液体积的1/10,得到浓缩液。在浓缩液中添加6倍体积的无水乙醇进行醇沉,4 ℃冰箱过夜后收集上清液继续添加4倍浓缩液体积的无水乙醇,4 ℃冰箱过夜后收集沉淀。用去离子水复溶沉淀,将复溶液体加入100~500 Da的透析袋进行透析除盐。
1.7.2 酶解结冷胶寡糖纯化方法
发酵降解结束后的发酵液离心除去菌体,上清液旋转蒸发浓缩至发酵液体积的1/10,得到浓缩液后添加7倍体积的无水乙醇进行醇沉,4 ℃冰箱过夜后收集上清液继续添加3倍浓缩液体积的无水乙醇,4 ℃冰箱过夜后收集沉淀。去离子水复溶沉淀后,通过Sevage[V(氯仿)∶V(正丁醇)=4∶1]法除蛋白[V(发酵液)∶V(Sevage试剂)=4∶1]。除蛋白后通过C18固相萃取小柱进一步除蛋白及色素等其他杂质。最后,将除杂后的糖液加入100~500 Da的透析袋进行透析除盐。
C18固相萃取小柱纯化方法[14]:用1~2倍柱体积的乙腈对小柱进行活化;用1~2倍体积的去离子水淋洗小柱;上样,并收集流出液体;用1~2倍体积去离子水淋洗小柱,并收集水洗液。
1.8 分析检测方法
1.8.1 还原糖含量测定
采用3,5-二硝基水杨酸(3,5-dinitrosalicylic acid, DNS)比色法。取1 mL稀释到适当浓度的发酵液,加入1.5 mL DNS试剂,沸水浴加热5 min,冷却至室温加蒸馏水至10 mL,550 nm下测定吸光度。用蒸馏水代替发酵液做空白对照。根据标准曲线计算出吸光度。
1.8.2 酶活力测定方法
取250 μL酶液与250 μL磷酸盐缓冲液(10 mmol/L,pH 6.0)混合,加入500 μL结冷胶水溶液(0.5 g/L)45 ℃水浴20 min,煮沸10 min终止反应。用DNS比色法测定反应体系中的还原糖含量,方法如1.8.1节所述。酶活力定义:1 min转化底物生成1 μmol还原糖所需的结冷胶裂解酶量为1个酶活力单位(U)。
1.8.3 薄层色谱分析
将制备好的样品点于薄层板上,点样量2 μL,晾干后放入装有展开剂的层析缸中将产物进行分离。层析完晾干薄层板,均匀喷上显色剂于105 ℃烘箱显色5 min。
酸解产物展开剂:V(乙酸乙酯)∶V(乙酸)∶V(水)=5∶5∶2;酶解产物展开剂:V(正丙醇)∶V(乙酸)∶V(水)=5∶2∶3;显色剂:900 mg地衣酚,25 mL水,375 mL乙醇,50 mL浓H2SO4。
1.8.4 FTIR分析
称取制备好的冻干后的结冷胶寡糖5 mg,使用傅里叶变换红外光谱仪在波数4 000~400 cm-1区间内进行扫描。
1.8.5 MALDI-TOF-MS分析
配制1 g/L的寡糖冻干样品溶液,取1 μL基质点于质谱预制靶上,再取1 μL样品与基质混合,真空泵中抽干液体水分待测量,在m/z400~2 000的质量范围内进行分析。
2 结果与分析
2.1 酸水解制备结冷胶寡糖及其条件优化
酸水解多糖是无规则降解,所得产物往往组分复杂[15]。酸浓度、水解时间、底物浓度均能影响酸水解效果,控制几个重要的影响水解效果的因素对结冷胶进行酸水解以提高产物得率,并将制得的酸解产物与酶解产物进行对比分析。图1-a是不同底物质量浓度对酸水解效果影响的薄层色谱分析,根据斑点颜色深浅可大致判断样品浓度。各聚合度的寡糖含量随底物浓度增加而增加,但当底物质量浓度超过5 g/L时变化不再明显,因此5 g/L是一个较为合适的底物质量浓度。图1-b是不同HCl浓度对酸水解效果影响的薄层色谱分析,当HCl浓度为0.25 mol/L时,主要产物为聚合度为5和8的寡糖。随HCl浓度增加,聚合度<5的寡糖逐渐增多,但聚合度>5的寡糖逐渐减少;当HCl浓度为0.75 mol/L时,聚合度为8的寡糖在薄层色谱板上几乎消失。尽管聚合度<5的寡糖随HCl浓度增加而逐渐增加,但当HCl浓度超过0.5 mol/L时,寡糖聚合度范围却在变小,逐渐出现一些单糖和二糖,这与得到更多低聚糖的目标不符。考虑目标产物特性,最终选取0.5 mol/L HCl进行下一步实验。

图1 底物质量浓度(a)、HCl浓度(b)、水解时间(c)对酸的水解效果Fig.1 The effects of substrate concentrations (a), HCl concentrations (b), and hydrolysis time (c) on acid hydrolysis
图1-c是不同水解时间对酸水解效果影响的薄层色谱分析,随水解时间的增加,聚合度<5的寡糖逐渐增加,聚合度>5的寡糖逐渐减少;但从4 h开始,聚合度>5的寡糖逐渐消失。并且水解时间越长产生的单糖和二糖越多,因此选择水解3 h进行下一步实验。最终酸水解选择底物质量浓度5 g/L,HCl浓度0.5 mol/L,水解3 h作为适宜水解条件制备结冷胶寡糖。
2.2 结冷胶裂解酶发酵条件优化
酶解多糖制备寡糖具有产物专一且温和的特点,本研究是利用前期实验室构建的含有结冷胶裂解酶基因(来源于Bacillussp.GL1)的重组毕赤酵母进行外源表达得到结冷胶裂解酶[16]。酶活力对于寡糖产量至关重要,酶活力越高则产物产量越高,因此有必要对发酵条件进行优化。重组结冷胶裂解酶是在毕赤酵母中以AOX1为启动子进行表达,因此在发酵过程中是以甲醇为唯一碳源诱导表达蛋白。发酵过程中氮源、碳源与pH都是微生物生长过程中的重要因素[17],因此分别研究了发酵初始酵母粉添加量、甲醇添加量与发酵初始pH对酶活力的影响。由图2-a可知,在酵母粉添加量为20 g/L时,重组结冷胶裂解酶酶活力最大;如图2-b所示,甲醇添加量为1%时,重组结冷胶裂解酶酶活力最大;从图2-c可知,pH为6.0时,重组结冷胶裂解酶酶活力最大。综上,选择发酵初始酵母粉添加量20 g/L,甲醇添加量1%,pH 6.0是重组结冷胶裂解酶的适宜发酵条件。

a-酵母粉添加量;b-甲醇添加量;c-pH图2 结冷胶裂解酶发酵条件优化Fig.2 Optimization of fermentation conditions for gellan lyase
2.3 酶解制备结冷胶寡糖及条件优化
传统的酶降解法是先通过发酵生产目标酶,然后直接用粗酶液或是将酶分离纯化后对多糖进行降解。用粗酶液直接进行降解需要先将酶生产出来再降解多糖,而酶的分离纯化因蛋白质结构复杂、易失活等原因往往需要几种纯化技术联用才能够得到目标蛋白[18],降解结束后还需要一系列的寡糖纯化步骤,这样极大地降低了酶法降解结冷胶生产结冷胶寡糖的效率。为了缩短酶法降解的时间,提高酶法降解效率,提出了毕赤酵母发酵耦合多糖水解一步法制备结冷胶寡糖的策略——在结冷胶裂解酶发酵初始添加结冷胶,以达到生产结冷胶裂解酶并实时降解多糖,实现一步制备结冷胶寡糖的目的。为此分别对发酵初始结冷胶添加量与发酵时间进行优化,以确定结冷胶寡糖产量更高的降解条件。图3-a显示结冷胶寡糖随结冷胶添加量增加而逐渐增加,当结冷胶添加量>2.0 g/L时,结冷胶寡糖几乎不再增加。图3-b显示结果与薄层色谱分析结果大致相同,随结冷胶添加量的增加还原糖产量逐渐增加,但当结冷胶添加量>2.5 g/L时,还原糖产量增加量变少。在优化过程中,观察到由于结冷胶在常温下溶解度不高的原因,当结冷胶添加量>2 g/L时,会造成发酵培养基凝结的问题,为了既能够生成较多的结冷胶寡糖又不影响结冷胶裂解酶的生产,综合考虑最终确定发酵初始添加2.0 g/L的结冷胶较为合适。
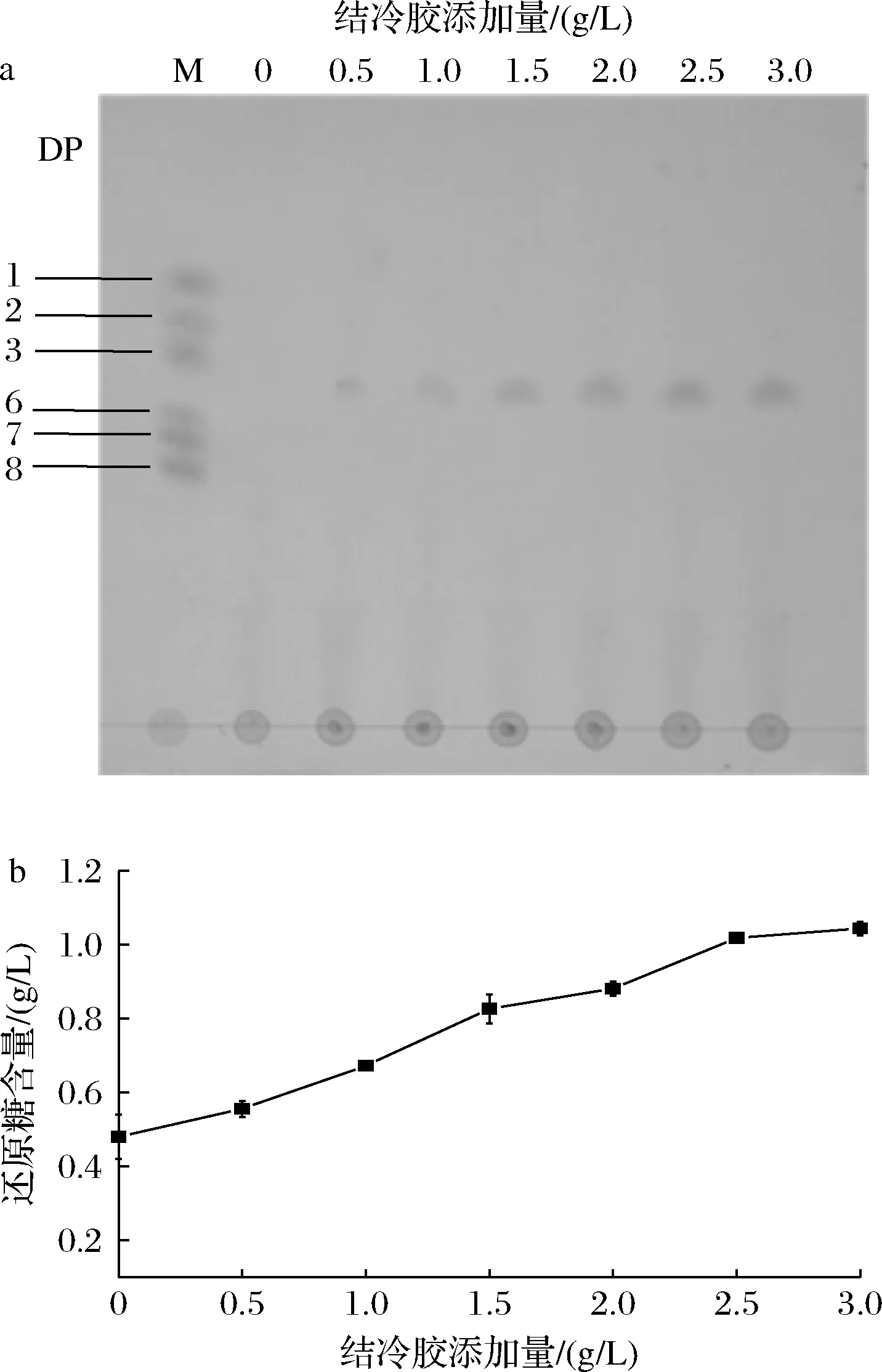
a-薄层色谱分析;b-还原糖产量图3 不同结冷胶添加量对酶降解效果的影响Fig.3 The effect of different gellan gum addition on enzymatic degradation
为进一步提高酶法降解的效率,对发酵时间进行优化,图4-a显示,结冷胶寡糖随时间增加而逐渐增多。图4-b可知,还原糖产量随发酵时间增加而增多,但当发酵96 h后、还原糖产量增速变慢,可能是发酵后期一些细胞产生自噬现象[19],限制了甲醇利用效率,导致发酵后期还原糖产量基本不再增加,最终确定发酵96 h为最佳发酵时间。

a-薄层色谱分析;b-还原糖产量图4 发酵时间对酶降解效果的影响Fig.4 The effect of fermentation time on enzymatic degradation
2.4 产物分析


a-商品结冷胶;b-酸水解结冷胶寡糖;c-酶水解结冷胶寡糖图5 降解产物的FTIR谱Fig.5 FTIR spectra of the degradation products
进一步通过MALDI-TOF-MS来确定2种水解方式得到的寡糖的分子质量。如图6-a所示,在[M-H]-m/z845.12、1 309.45、1 955.67处检测到3个结冷胶寡糖成分,表明酸水解后得到了聚合度为5、8、12的结冷胶寡糖。图6-b是酶解结冷胶寡糖的质谱分析图,在[M+Na]+m/z669.34处检测到一个结冷胶寡糖成分,表明酶解后得到了聚合度为4的结冷胶寡糖,这一结果正是酶解产物的专一性导致的。因此,酶解法能够克服酸解法产物组分复杂的特点,成功制备出单一聚合度的结冷胶寡糖。
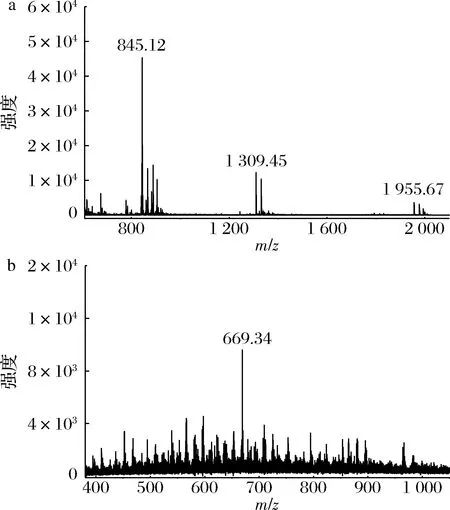
a-酸解产物MALDI-TOF-MS分析;b-酶解产物MALDI-TOF-MS分析图6 降解产物的MALDI-TOF-MS谱图Fig.6 MALDI-TOF-MS spectrum of the degradation products
3 结论
猜你喜欢
阅读(快乐英语中年级)(2022年5期)2022-06-03
农药科学与管理(2019年7期)2019-11-29
江西水产科技(2019年3期)2019-09-10
天津农业科学(2019年4期)2019-05-31
中国食物与营养(2018年10期)2018-11-12
邢台学院学报(2016年4期)2016-02-28
云南科技管理(2015年5期)2015-12-22
中国茶叶加工(2015年3期)2015-02-27
中国海洋大学学报(自然科学版)(2014年8期)2014-02-28
食品工业科技(2013年10期)2013-08-07
