
两种新型植物蛋白替代豆粕在大规格鲤饲料中的应用效果研究
2023-09-06 08:48范泽李晨辉吴迪李晋南刘庆武王良王连生
水产学杂志 2023年4期
范泽,李晨辉,2,吴迪,李晋南,刘庆武,王良,王连生
(1.中国水产科学研究院黑龙江水产研究所,黑龙江省水生动物病害与免疫重点试验室,黑龙江 哈尔滨 150070;2.农业农村部水产品质量安全控制重点实验室,北京 100141;3.黑龙江省双鸭山市农业技术推广总站,黑龙江 双鸭山 155100;4.潍坊加易加生物科技有限公司,山东 潍坊 261100)
豆粕蛋白含量高、氨基酸组成合理及品质较稳定等优势成为淡水鱼饲料配方中最常用的植物蛋白源[1,2]。当前豆粕供需趋紧,饲料原料成本压力明显增大。因此通过豆粕减量技术,开发利用新型蛋白源成为水产饲料研究的热点[3]。单细胞蛋白方便获得和高蛋白含量等优势,是替代鱼粉、豆粕的潜在优质蛋白[4]。小球藻是目前研究较为广泛的单细胞蛋白[5],已应用于鲫(Carassius auratus)[6]、斑马鱼(Danio rerio)[7]等的饲料中。利用发酵和酶解等相关技术去除多种植物蛋白源抗营养因子是提高植物蛋白替代鱼粉的有效途径。其中,棉籽浓缩蛋白是一种脱苯脱毒后蛋白质含量较高的植物蛋白源,已在金鲳(Trachinotus ovatus)[8]、虹鳟(Oncorhynchus mykiss)[9]和大口黑鲈(Micropterus salmoides)[10]等研究中有广泛报道。但在淡水鱼饲料中上述两种形式的蛋白源替代豆粕的应用研究还未见报道。
不同的蛋白源不仅影响生长、抗氧化能力及免疫机能[11],还直接影响肌肉营养成分、氨基酸、质构、持水能力等肌肉品质[12,13]。作为评价鱼肉风味的重要指标,呈味氨基酸组成影响着鱼肉的鲜味和口感[14]。质构特性决定着鱼肉的口腔行为及感官性状,与肌肉的营养成分密切相关[15]。因此,在豆粕替代研究中也应注重对肌肉品质变化的考察。
目前,鲤商品饲料配方中豆粕常用量在20%~40%之间[16],因此豆粕已成为限制鲤养殖的主要因素。松浦镜鲤(Cyprinus carpio Songpu)具有生长速度快、肉味鲜美、含肉率高、抗逆抗病能力强等优点,已成为我国主要的鲤养殖品种[17,18]。基于此,本试验通过探究棉籽浓缩蛋白及小球藻完全替代豆粕对大规格松浦镜鲤生长性能及肌肉品质的影响,以评估两种新型蛋白源在鲤饲料中替代豆粕的潜在应用价值。
1 材料与方法
1.1 材料
分别以豆粕(粗蛋白43.9%,粗脂肪1.24%)、棉籽浓缩蛋白(粗蛋白63.1%,粗脂肪1.38%,购自新疆金兰植物蛋白有限公司)及小球藻(粗蛋白57.5%,粗脂肪10.2%,中国科学院水生生物研究所提供)为饲料蛋白质源,豆油为脂肪源,配置蛋白水平为30%,脂肪水平为5.8%的3 组饲料,其组成及营养水平见表1,饲料氨基酸组成见表2。首先将所有干性原料粉碎后,过60 目筛,按逐级放大原则用混匀机搅拌混匀,后依据饲料配方比例加入豆油,在混匀机中充分混合后,制成直径为4.00 mm 的膨化颗粒饲料,自然风干后于-20℃冰箱中保存备用。

表1 饲料组成和营养水平(干物质基础)Tab.1 Diet ingredient and nutrient level(Dry matter basis)of the diets used in the experiment
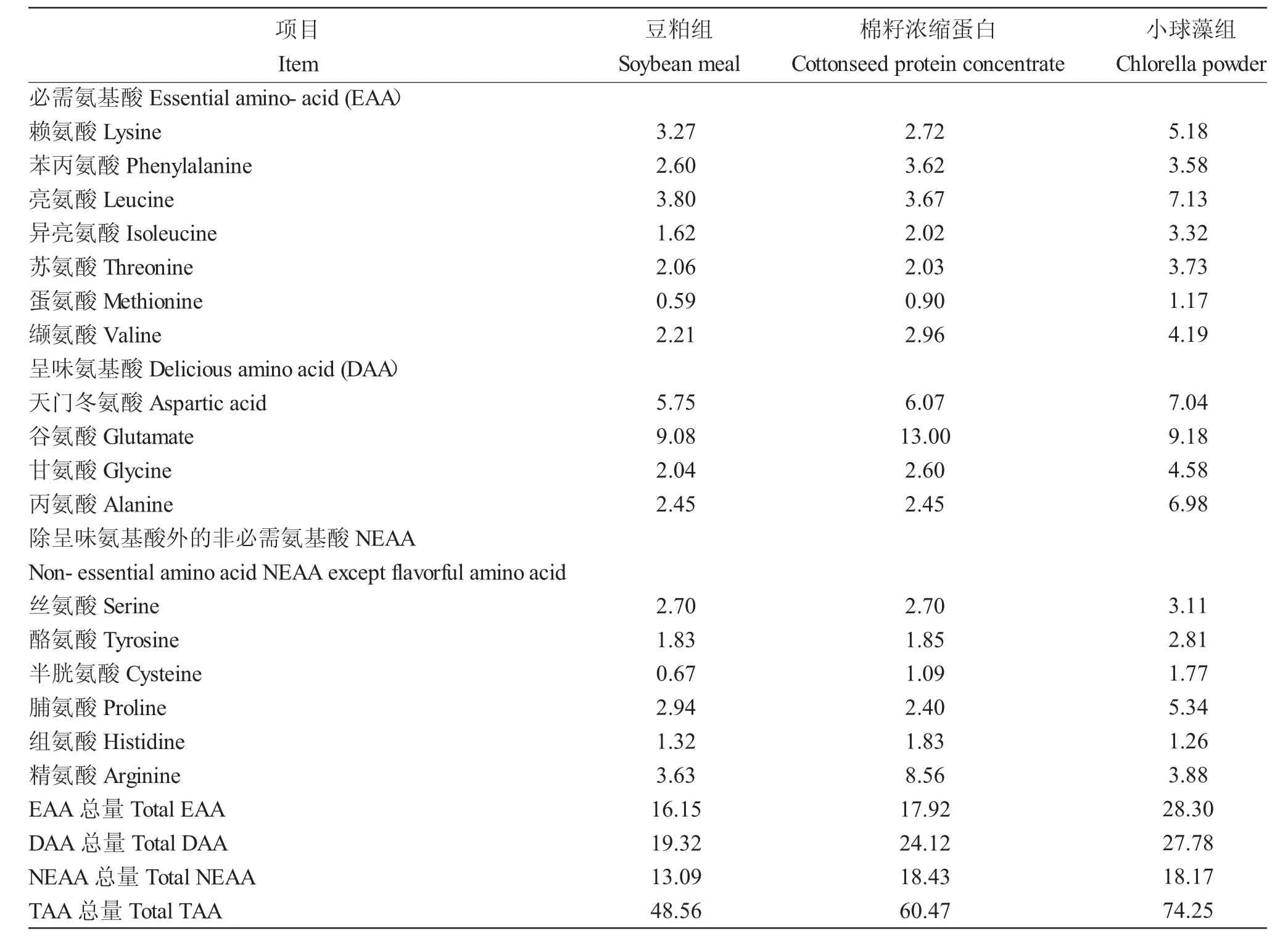
表2 饲料氨基酸组成(风干基础,%)Tab.2 Amino acid composition in experimental diets(air-dry basis,%)
1.2 方法
暂养及养殖试验均在黑龙江水产研究所呼兰试验站完成。试验鱼经消毒处理后,置于1.0 m×2.0 m×2.0 m 暂养网箱中驯化15 d,驯化期间32%蛋白的商品饲料。正式试验开始前,将松浦镜鲤饥饿24 h 后,挑选270 尾体格健壮、规格一致的初始体质量为(247.22±9.90)g 个体,养殖于9 个池塘沉性网箱(1.0 m×2.0 m×2.0 m)中,每个网箱30尾,随机分为6 组,每组3 个重复(网箱),分别投喂表1 中的3 种饲料,持续饲养8 周。整个试验周期,固定日投喂率为4% BW/d,8:00 am、13:00 pm 和17:30 pm 投喂;每14 d 测定1 次各网箱鱼的质量并按照设定的投喂率调整投喂量。试验期间水温在28 ℃~32 ℃,溶解氧为6.0 mg/L 左右,pH 为7.6~8.0。
8 周养殖试验结束时,禁食24 h 后以养殖网箱为单位进行称重,计数松浦镜鲤成活的尾数;随后每个养殖网箱随机选取3 尾鱼,称重,测定体长,用100 mg/L 的MS-222 麻醉后,取侧线上方背部肌肉用于测定肌肉品质参数。鱼体左侧背部肌肉用于肌肉品质的测定。用于质构参数和剪切力检测的肌肉样品需现采现测,质构参数样品(图1 中1 号部位)切割为2 cm×2 cm×0.5 cm 的肉块,剪切力样品(图1 中2 号部位)切割为3 cm×1 cm×0.5 cm 的肉条。滴水损失试验样品(图1 中3 号部位)约为2.00 g,pH 测定试验样品(图1 中4 号部位)约0.5 g。鱼体右侧背部肌肉用于用于基因表达的测定。

图1 鱼体左侧用于肌肉品质测定的肌肉对应部位Fig.1 The corresponding muscle site on the left side of the fish body for muscle quality measurement
依GB/T5009.124-2003 测定试验鱼肌肉及饲料中氨基酸含量。称量冷冻干燥好的肌肉样品40~50 mg(精确至0.1 mg),装入50 mL 的安瓿瓶并加入6 mol/L 盐酸10 mL,迅速封管后放于恒温干燥箱(110±1)℃中水解24 h;冷却后打开安瓿瓶,用双蒸水定容到150 mL 旋转蒸发瓶中,在旋转蒸发器(60 ℃)中抽真空,蒸发至干;用0.02 mol/L 盐酸冲洗蒸发瓶数次,洗涤液转至10 mL 容量瓶中,并用0.02 mol/L 盐酸定溶,充分混匀后即为试样水解液吸取试样水解液及混合氨基酸标准液2~3 mL,经过滤至自动进样瓶中并加盖,用日立835-50 型全自动氨基酸分析仪测定。
使用质构检测分析仪Food Technology Corporation TPA(美国)检测肌肉样品的质构参数。配备平底圆柱型探针(型号为P/36R),用于检测各常规的质构指标。再用Auto-5g 探针型探头检测样品的剪切力,每个样品进行5 次TPA 切割压缩处理。具体TPA 检测参数设置如下:数据曲线记录的预测试速度为3 mm/s,试验过程中的测试速度为2 mm/s,测试后的记录速度为2 mm/s,停留时间为5 s,数据收集率为200 pps。鱼肉样品的高度和最大压力之间的关系(单位:g),需要设置样品的压缩高度为样品原高度的20%,以减少对肌肉结构的损伤。肌肉质构和剪切力的测试均在室温条件下进行。在剪切力测试中,记录TPA 纹理曲线,确定曲线中的峰值作为最大剪切力,一个样品测量三次,取平均值作为最终的检测结果。
滴水损失:取2.00 g 左右肌肉,称重后将其悬挂在体积为250 mL 倒扣的纸杯中,用保鲜膜封口,放置于4 ℃的冰箱中保存,24 h 后去除,用滤纸吸除表面析出的水珠后再次称重并记录,计算滴水损失率。
pH 测定:取0.5 g 背肌白肉,充分剪碎,加入10 m L 蒸馏水,匀浆,4 ℃4 500 r/min 离心10 min,取上清,用酸度计测pH,重复3 次,取平均值作为样品pH。
肠道解冻后用预冷的生理盐水冲洗干净,用滤纸吸干后称重。在预冷的生理盐水中漂洗,除去血液,滤纸拭干,用移液管加入9 倍于组织块质量的预冷匀浆介质(0.85%的氯化钠注射液),匀浆,4 ℃、4 500 r/min 离心15 min,取上清液备用。肠道中总蛋白质含量、胰蛋白酶、淀粉酶及脂肪酶活性用南京建成生物工程研究所的试剂盒测定。
肌肉中蛋白合成相关基因表达的测定:利用Trizol 法提取各组织总RNA,参照TaKaRa Prime-ScriptTM RT reagent Kit with gDNA Eraser(Perfect Real Time)(Code No:RR047A)说明书,将总RNA 反转录为cDNA。根据Genbank 中现有的鲤鱼雷帕霉素靶蛋白(TOR)、蛋白激酶B(AKT)、S6 蛋白激酶1(S6K1)、4E 结合蛋白1(4EBP1)及β-actin 保守序列设计引物(表3)。所有引物均由生工生物工程(上海)有限股份公司合成。

表3 基因表达试验所用引物Tab.3 Primers for gene expression used in this study
1.3 数据统计与分析
所有数据均以平均值±标准差表示,用Excel 2007 及SPSS23.0 进行分析处理和单因素方差分析(One-way ANOVA),若差异达到显著水平(P<0.05),则进行Duncan′s 法多重比较。
式中:t 为试验天数(d);Nt为终末存活尾数;W0为平均初始体质量(g);W 为平均终末体质量(g);F为饲料摄取量干重(g);P 为饲料蛋白质含量(%)。
2 结果与分析
2.1 棉籽浓缩蛋白及小球藻替代豆粕对大规格鲤生长性能及饲料利用的影响
各组鲤生长性能及饲料利用结果如表4 所示。小球藻组的终末体质量、增重率、特定生长率及蛋白质效率均最高,其中增重率、特定生长率及蛋白质效率显著高于豆粕组和棉籽浓缩蛋白组(P<0.05),终末体质量显著高于棉籽浓缩蛋白组(P<0.05)。小球藻组的饲料系数为最低,显著低于豆粕组(P<0.05)。
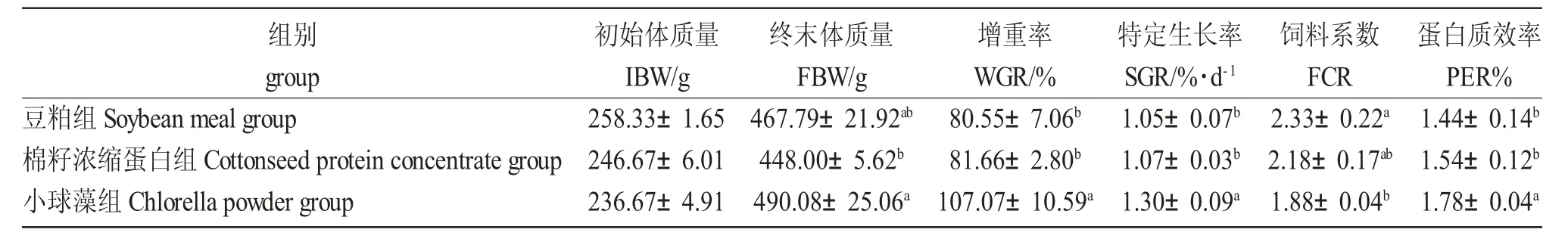
表4 棉籽浓缩蛋白及小球藻替代豆粕对大规格鲤生长性能及饲料利用的影响Tab.4 Effect of cottonseed protein concentrate and chlorella powder as soybean meal alternative on growth performance and feed utilization of large-sized Songpu mirror carp(C.carpio Songpu)
2.2 两种新型植物蛋白源对大规格鲤肠道消化酶活性的影响
小球藻组鲤肠道胰蛋白酶、酸性磷酸酶及碱性磷酸酶活性最高,其中肠道胰蛋白酶及酸性磷酸酶活性显著高于豆粕组和棉籽蛋白组,碱性磷酸酶活性显著高于豆粕组(P<0.05)。不同蛋白源组间肠道脂肪酶及淀粉酶活性无显著差异(P>0.05)(表5)
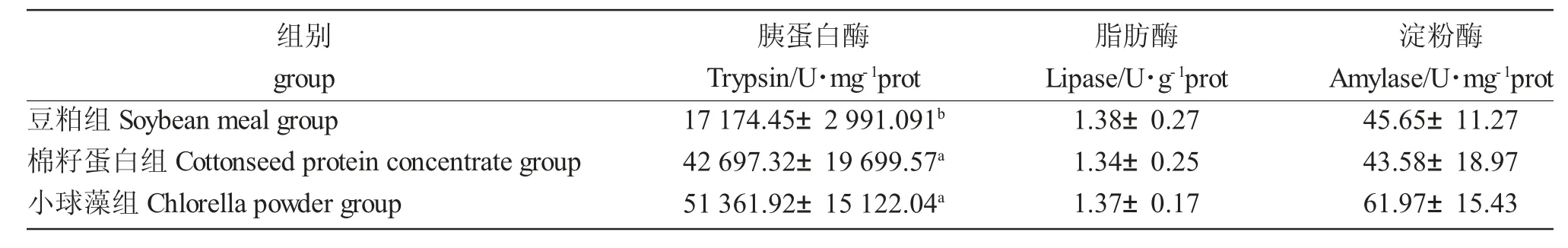
表5 两种新型植物蛋白源对大规格鲤肠道消化酶活性的影响Tab.5 Effects of two new pla nt protein sources on the intestinal digestive enzyme activities of large-sized Songpu mirror carp
2.3 棉籽浓缩蛋白及小球藻替代豆粕对大规格鲤肌肉氨基酸组成的影响
在必需氨基酸中,小球藻组肌肉亮氨酸含量显著高于棉籽浓缩蛋白组(P<0.05),其余必需氨基酸含量在各组间无显著差异(P>0.05)。在呈味氨基酸中,小球藻组肌肉谷氨酸含量显著高于豆粕组(P<0.05),各组间其余呈味氨基酸含量在无显著差异(P>0.05)。在除呈味氨基酸外的非必需氨基酸中,小球藻组肌肉脯氨酸含量显著高于豆粕组和棉籽浓缩蛋白组(P<0.05),其余必需氨基酸含量在各组间无显著差异(P>0.05)。从氨基酸组成看,小球藻组的必需氨基酸总量、呈味氨基酸总量及其余非必需氨基酸总量均为最大值,其中小球藻组的呈味氨基酸总量显著高于豆粕组(P<0.05)(表6)。
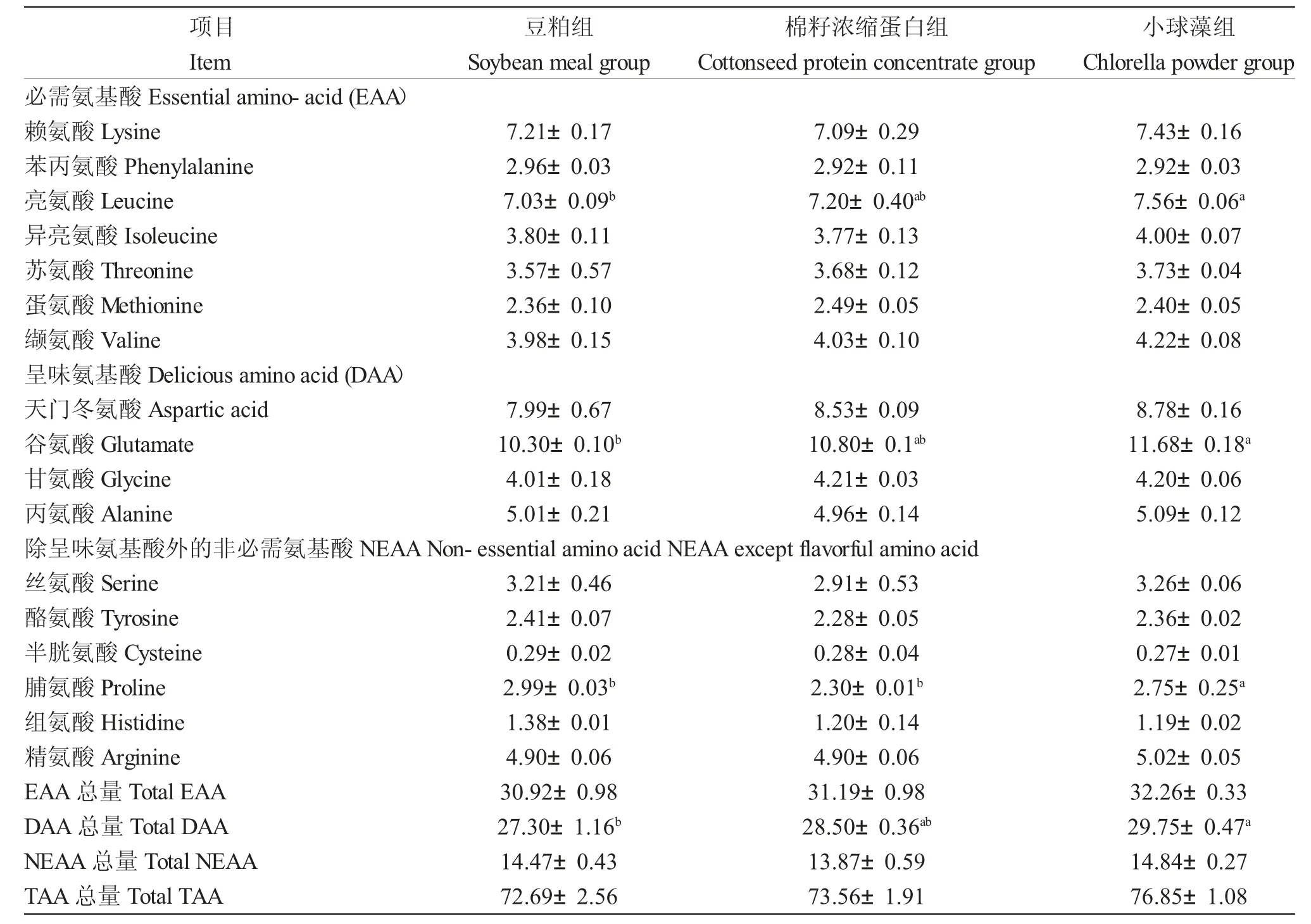
表6 棉籽浓缩蛋白及小球藻替代豆粕对大规格鲤肌肉氨基酸组成的影响(占肌肉干重比例,%)Tab.6 Effect of cottonseed protein concentrate and chlorella powder as soybean meal alternative on muscular amino acid composition of large-sized Songpu mirror carp(C.carpio Songpu)(Proportion of dry muscle weight,%)
2.4 棉籽浓缩蛋白及小球藻替代豆粕对大规格鲤肌肉质构参数的影响
各组鲤肌肉质构的参数如表7 所示,小球藻组鲤肌肉的硬度、内聚性、弹性及咀嚼性均最高,其中硬度及胶粘性显著高于豆粕组和棉籽浓缩蛋白组(P<0.05),内聚性、弹性及咀嚼性显著高于豆粕组(P<0.05)。

表7 棉籽浓缩蛋白及小球藻替代豆粕对大规格鲤肌肉质构参数的影响Tab.7 Effect of cottonseed protein concentrate and chlorella powder as soybean meal alternative on muscle texture parameters of large-sized Songpu mirror carp(C.carpio Songpu)
2.5 棉籽浓缩蛋白及小球藻替代豆粕对大规格鲤肌肉剪切力、滴水损失及pH 的影响
各组鲤肌肉剪切力、滴水损失及pH 的结果如表8 所示。小球藻组的剪切力最高,但与其余两组无显著差异(P>0.05)。小球藻组鲤的肌肉滴水损失最低,显著低于豆粕组(P<0.05)。不同蛋白源组间肌肉pH 无显著差异(P>0.05)。
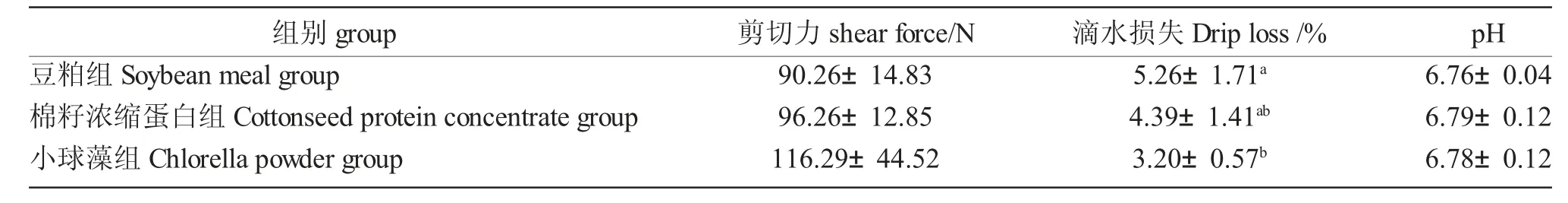
表8 棉籽浓缩蛋白及小球藻替代豆粕对大规格鲤肌肉剪切力、滴水损失及pH 的影响Tab.8 Effect of cottonseed protein concentrate and chlorella powder as soybean meal alternative on muscular shear force,drip loss and pH of large-sized Songpu mirror carp(C.carpio Songpu)
2.6 棉籽浓缩蛋白及小球藻替代豆粕对大规格鲤肌肉蛋白合成相关基因表达的影响
各组鲤肌肉蛋白合成相关基因表达的结果如表9 所示。小球藻组的tor、akt、s6k 及4ebp1 基因表达量均为最高,其中tor、akt、s6k 及4ebp1 基因表达量显著高于豆粕组和棉籽浓缩蛋白组,s6k 基因表达量显著高于棉籽浓缩蛋白组(P<0.05)。

表9 棉籽浓缩蛋白及小球藻替代豆粕对大规格鲤肌肉蛋白合成相关基因表达的影响Tab.9 Effect of cottonseed protein concentrate and chlorella powder as soybean meal alternative on protein synthesis gene expression in muscle of large-sized Soybean meal group(C.carpio Songpu)
3 讨论
蛋白源是水产动物饲料中价格最为昂贵、营养价值最高的部分。为了评估在大规格松浦镜鲤饲料中棉籽浓缩蛋白及小球藻替代豆粕的潜在应用价值,本研究采用了完全替代豆粕的方式,发现以69.1%豆粕作为唯一饲料蛋白源时,会显著降低大规格松浦镜鲤的生长性能及饲料利用率,这与孙碧桂[19]在鲤上的研究结果相符,即添加54%豆粕完全替代鱼粉时显著抑制了鲤生长及饲料利用率。而本研究中棉籽浓缩蛋白及小球藻完全替代豆粕并未降低松浦镜鲤生长性能及饲料利用率,小球藻组的增重率、特定生长率及蛋白质效率甚至显著优于豆粕组。这说明相较于豆粕、棉籽浓缩蛋白及小球藻具有更好的促生长效应。
消化是影响蛋白源用于鱼体生长的首要限制因素。胰蛋白酶是由胰腺分泌后广泛分布于鱼类的肝胰脏及肠道中,在鱼类消化蛋白质的过程中发挥着关键作用。在本实验中,豆粕组鲤的胰蛋白酶活性明显低于棉籽浓缩蛋白组及小球藻组,而脂肪酶和淀粉酶活性未受显著影响。这可能是由于豆粕作为唯一饲料蛋白源时,饲料中胰蛋白酶抑制因子浓度达到一定浓度,使鱼类肠道消化酶活性被抑制,引发肠道机能紊乱,导致营养物质吸收能力减弱,抑制鱼体生长[20]。可通过物理加工方式[21]或复合蛋白源配伍[22]的方式消除豆粕相关抗营养因子,或可开发低抗营养因子的新型蛋白源。棉籽浓缩蛋白经过脱酚处理降低了自身抗营养因子含量。申建飞等[23]研究发现,棉籽浓缩蛋白替代24%、36%鱼粉显著提高了卵形鲳鲹(Trachinotus ovatus)幼鱼前肠及后肠中的糜蛋白酶活性,提高了肠道消化蛋白能力,与本研究发现一致;小球藻自身营养较为均衡,不含抗营养因子,其独有的细胞活性物质——生长因子(Chlorella Growth Factor,CGF)能提高水产动物肠道消化酶活性和消化酶功能[24]。因此蛋白源中抗营养因子对鱼类肠道消化酶活性的影响是引起生长差异的主要因素之一。
鱼类的生长与饲料及肌肉中必需氨基酸的组成存在密切关系。豆粕饲料组必需氨基酸含量低于棉籽浓缩蛋白和小球藻组,支链氨基酸(亮氨酸、异亮氨酸及缬氨酸)含量相对缺乏。豆粕组鲤肌肉的必需氨基酸、亮氨酸、异亮氨酸及缬氨酸含量低于棉籽浓缩蛋白和小球藻组,其中亮氨酸含量显著低于小球藻组。上述结果表明,支链氨基酸含量及组成差异是造成豆粕组鲤生长性能弱于棉籽浓缩蛋白和小球藻组的重要原因。支链氨基酸能够参与蛋白质代谢,调节蛋白质合成的支链联氨基酸主要是亮氨酸[25],其可通过参与调节雷帕霉素靶蛋白(TOR)信号通路,提升鱼类合成蛋白质效率,促进机体生长[26,27]。在团头鲂(Megalobrama amblycephala)[28]饲料中添加1.74%亮氨酸显著上调肌肉tor 基因表达量,增强鱼体蛋白质沉积。本试验也发现,肌肉亮氨酸含量较高的小球藻组tor、akt 及4ebp1 基因表达量显著高于豆粕组,这表明小球藻可能通过提升机体亮氨酸水平增强mTOR 信号通路活性,促进机体蛋白沉积,这可能小球藻促生长效应优于豆粕的原因之一,但具体调控机制需深入研究。
肌肉品质由肌肉风味及肌肉质量两个因素共同决定。非必需氨基酸中的呈味氨基酸组成是决定鱼类肌肉风味的主要因素,包括呈甘味的甘氨酸和丙氨酸及呈鲜味的谷氨酸和天门冬氨酸,其中作用最大的是谷氨酸,甘氨酸次之[29]。豆粕组的天门冬氨酸,谷氨酸及甘氨酸含量均低于棉籽浓缩蛋白和小球藻组,肌肉呈味氨基酸组成的变化与之相似。与小球藻组相比,以豆粕为唯一蛋白源时会显著降低大规格松浦镜鲤谷氨酸含量,破坏鱼肉的风味,这与姜俊等[30]对建鲤的研究结果一致。该研究表明,豆粕完全替代鱼粉作为主要饲料蛋白源后,建鲤肌肉中谷氨酸和甘氨酸等呈味氨基酸含量显著降低。揭示不同蛋白源呈味氨基酸组成的不同会引起肌肉中呈味氨基酸的相应变化,影响鱼肉的良好风味。
影响肌肉质量的物理因素主要有质构参数、剪切力、系水力及pH 等。质构参数及剪切力是描述肌肉组织特性的重要指标,通常受鱼种类、基因型、饲料及养殖环境变化等多种因素的影响[31]。质构参数是通过模拟人类牙齿咀嚼来表征肉质口感,主要参数包括:硬度、内聚性、弹性、咀嚼性等,而剪切力是通过模拟人类牙齿切割肌纤维来表征肌肉嫩度[15]。与豆粕组相比,棉籽浓缩蛋白及小球藻组通过增强大规格松浦镜鲤肌肉剪切力来增加肌肉硬度,提升肌肉口感[32];其次,两种蛋白源通过提高内聚性来加强肌肉细胞间的结合力,提升鱼肉抵抗受损的能力,使鱼肉在被咀嚼时更加细腻,口感更佳[33]。系水力通常用滴水损失来表征,滴水损失越小表明系水力越强,肉质越好[34]。本试验中,相较于豆粕组,棉籽浓缩蛋白及小球藻组能够降低大规格松浦镜鲤背部肌肉的滴水损失率,增强其系水力,尤其是在小球藻组。肌肉的质构特性及系水力同其蛋白质合成能力关系密切,丰富的蛋白质和水化层形成网状结构,增强肌肉纤维密度及抵抗外力的能力[35]。本研究中,相较于豆粕组,小球藻组能够通过显著上调TOR 信号通路相关基因的表达来提升肌肉蛋白质沉积能力,这可能是小球藻能够更好地改善大规格松浦镜鲤肌肉质量的机理之一,但具体机制有待于深入挖掘。
结论
本试验通过完全替代的方式,从生长性能及肌肉品质角度入手揭示了棉籽浓缩蛋白及小球藻替代豆粕作为大规格松浦镜鲤饲料蛋白的应用前景。结果表明,棉籽浓缩蛋白和小球藻均在一定程度上替代豆粕蛋白源,但小球藻效果更佳。小球藻替代豆粕后可以提高肠道消化蛋白能力,促进鱼体的生长及对饲料的利用效率,还可以改善肌肉的氨基酸组成,提升鱼肉的风味,并通过改善质构特性和系水力来增强鱼肉的口感。
猜你喜欢
——全棉籽的加工与利用
中国畜牧杂志(2020年8期)2020-12-18
当代水产(2019年11期)2019-12-23
当代水产(2019年7期)2019-09-03
当代水产(2019年6期)2019-07-25
陶瓷学报(2019年5期)2019-01-12
河南畜牧兽医(2017年8期)2017-11-24
山西农业科学(2016年1期)2017-01-05
中国棉花加工(2016年5期)2016-12-09
电源技术(2016年2期)2016-02-27
湖北工业大学学报(2014年1期)2014-10-13
