
杨梅单宁-酪蛋白复合膜的制备及性能评价
2023-08-25 07:16邓霜琪廖晶晶吴海柱杨若彤郑青状周晓剑
食品工业科技 2023年17期
邓霜琪,廖晶晶,吴海柱,杨若彤,郑青状,周晓剑
(西南林业大学,云南省木材胶黏剂及胶合制品重点实验室,云南昆明 650224)
随着“可食用包装膜”概念的兴起,蛋白质具有可食用性、良好成膜性和生物降解性,并且原料来源广泛,是近年来食品包装膜常用的天然高分子聚合物成膜材料[1-4]。酪蛋白(casein,CA)是牛奶中的主要成分,其结构中存在大量的脯氨酸肽,α-螺旋和β-折叠结构容易转变成随机卷曲结构[5],不具有双硫键,没有明显的二级结构和三级结构,因此,酪蛋白具有良好的水溶性、分散性和成膜性[6-7]。酪蛋白膜主要通过蛋白质分子链中不同氨基酸通过蛋白质分子链间形成氢键、离子键、疏水键等作用下,依靠的作用力构建膜的网状结构,形成具有一定力学强度和阻隔性能的薄膜[8]。然而,由于亲水基团的存在,纯酪蛋白膜存在水蒸气透过率高和力学性能不高等问题,极大限制了酪蛋白膜的应用[9-10]。交联改性是提高蛋白膜综合性能效果最为有效的方法之一,其中,醛类交联剂(甲醛、乙二醛、戊二醛、双醛淀粉等)使用最为广泛[11],但未反应的醛基具有很强的细胞毒性;天然产物中提取的京尼平和转谷氨酰胺酶等交联剂安全有效[12-13],但其价格昂贵,难以推广使用。
单宁是维管束植物进化过程中衍生出来的一种自身保护性的次生代谢物,是可以与生物碱、明胶和其它蛋白质发生沉淀的水溶性多酚化合物,分子量通常为500~3000 kDa[14-15]。其广泛存在于植物的树皮、根、叶、花和果等组织中,在树皮中含量尤为丰富,许多针叶树皮中单宁的含量高达20%~40%。单宁的存在可以帮助植物抵抗阳光引起的紫外线损伤和植物体内的氧化应激,防止有害自由基破坏细胞结构,保护植物免受细菌和真菌的感染[14,16]。由于单宁的酚羟基结构所带来的诸如抗氧化性、抗菌性、屏蔽紫外光等生物活性,使其在食品包装等领域备受关注[17-18]。
单宁与蛋白的相互作用机制是单宁的重要研究方向之一,早期的研究认为单宁与蛋白的结合为非共价键,单宁在疏水键的驱动作用下向蛋白质分子表面靠近,并在进入疏水袋后,与蛋白质的H-受体位点和多酚的羟基基团之间发生多点氢键结合,其他反应如离子键发生在蛋白质带正电荷的基团(如赖氨酸的ε-氨基)和多酚带负电荷的羟基之间。在改变溶液环境时,如在碱性条件下,单宁的苯环结构氧化,形成邻苯醌或者半-邻苯醌的结构,单宁与蛋白质分子链间以共价键形式结合,可以有效的增强蛋白质分子链间的作用力[19-20]。由此可见,在单宁-蛋白膜的成膜过程中,蛋白在pH 的作用下,三维空间结构展开,亚基解离,原来埋藏在分子内部的部分疏水基团、巯基和二硫键等暴露出来,在热力学作用下,单宁通过与蛋白质分子链形成非共价键和共价键,强化了蛋白质分子链间的作用力,进而形成更致密均匀的网络结构[19-20]。单宁近年来常作为蛋白膜的天然多功能交联剂,用于提高蛋白膜的力学强度和水蒸气阻隔作用[21-25],如橡树单宁改性明胶膜[10]、黑荆树单宁改性的纤维素膜[26]和大豆蛋白膜[24]等研究表明,改性后的薄膜不仅力学性能得到改善,还具有较好的抗氧化性和紫外线阻隔能力,可以延长食品的保质期[6]。由此可见,木材单宁抽提物在食品包装领域极具应用前景[27-28],但其改性酪蛋白膜的研究还非常有限。
杨梅单宁(BT)是我国最主要的商业单宁类型之一,来源广泛,价格低廉。课题组前期的研究表明,杨梅单宁交联改性大豆分离蛋白膜后,其力学性能和水蒸气阻隔能力有明显提升[29]。由于单宁和蛋白结构性能上的多样性,单宁与不同蛋白相互作用力不同,将直接影响最终蛋白膜的性能[10]。为探究杨梅单宁对酪蛋白膜交联改性作用的影响,本研究使用不同比例的杨梅单宁作为交联剂,采用延流成膜的方式制备复合膜,并对杨梅单宁改性酪蛋白膜(BT-CA)的水蒸气透过率、拉伸、抗氧化等性能进行探讨,以期获得安全环保、功能多样化的酪蛋白复合膜,同时为杨梅单宁在酪蛋白膜中的应用提供实践和理论依据。
1 材料与方法
1.1 材料与仪器
酪蛋白(CA) 纯度90%,国药集团化学试剂有限公司;杨梅单宁 纯度72%,工业级,武鸣栲胶厂;丙三醇 分析纯,天津市风船化学试剂科技有限公司;DPPH(2,2-联苯基-1-苦基肼基) 纯度96%,上海麦克林生化科技有限公司;氢氧化钠 分析纯,成都市科龙化工试剂厂;无水氯化钙 分析纯,广东光华科技股份有限公司。
JJ200 电子天平 常州奥华仪器有限公司;101电热鼓风干燥箱 北京市永光明医疗仪器有限公司;BC801 埃维特数显厚度仪 河南省邦特工量具有限公司;SUST5569 万能力学试验机 珠海市三思泰捷电气设备有限公司;WFZUV-4802S 紫外分光光度计珠海市三思泰捷电气设备有限公司;UH5300 紫外分光光度计 日本三立有限公司;Zeiss Sigma 300扫描电子显微镜 德国卡尔蔡司公司;TG209F1 热重分析仪 德国耐驰公司;FE28 pH 计 梅特勒-托利多仪器(上海)有限公司;TENSOR27 傅里叶红外红外光谱仪 德国布鲁克光谱仪器公司。
1.2 实验方法
1.2.1 薄膜的制备 首先,配制浓度为6wt%的CA水溶液,用2 mol/L 的氢氧化钠溶液调节其pH 为11,在室温下进行磁力搅拌至完全溶解。其次,将一定量的杨梅单宁加入到2 mL 蒸馏水中,摇匀。将杨梅单宁水溶液加入到CA 水溶液中,再加入丙三醇,继续搅拌10 min,使其充分混匀后置于80 ℃水浴中加热搅拌20 min 后取出,冷却消泡后的成膜液静置待用。将45 mL 成膜液浇铸到尺寸为16.6 cm×26.6 cm 的特氟龙涂层金属模具中,保持水平放置,在室温下干燥48 h。干燥好的薄膜揭膜后放置于干燥器中储存,用于薄膜性能的测试。薄膜制备配方如表1 所示,成膜样品依次标记为C1、C2、C3、C4、C5,代表单宁的添加量分别为酪蛋白质量的0%、5%、10%、15%、20%。
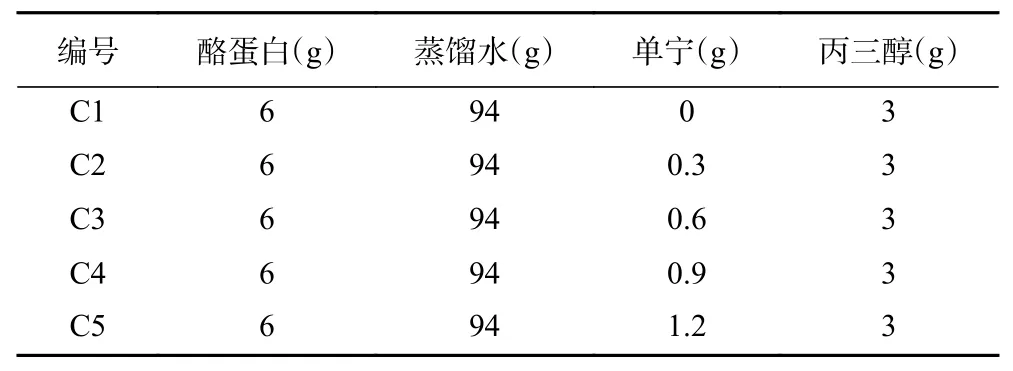
表1 杨梅单宁改性酪蛋白膜的制备配方Table 1 Formulation for the preparation of BT-CA films
1.2.2 测试与表征
1.2.2.1 化学结构分析 膜的化学结构及相互作用力通过傅里叶红外光谱仪采用衰减全反射模式(ATR)进行表征,扫描范围为4000~600 cm-1,扫描次数为32 次,分辨率为4 cm-1。
1.2.2.2 厚度测定 采用随机取样法在薄膜样品上随机选取10 个点,使用测厚仪进行薄膜的厚度测量,取其平均值。
1.2.2.3 水溶性测定 将2 cm×2 cm 的薄膜样品放置于105 ℃烘箱中干燥至恒重,然后精确称重,计为m1(g),将称重后的样品浸入盛有50 mL 蒸馏水的培养皿中静置24 h,将蒸馏水溶液过滤后,取出不溶物,在105 ℃下再次干燥至恒重,计为m2(g)。根据溶解前后的质量变化来计算蛋白膜的水溶性(WS),每组样品平行测试5 次,计算公式如下:
1.2.2.4 不透明度测定 薄膜的不透明度使用紫外分光光度计UH 5300 在600 nm 扫描吸光度进行表征。薄膜样品规格为4 cm×1 cm,放进调试好的仪器里测量吸光度,测定时样品紧贴比色皿一侧的内表面,每一样品平行测量3 次,取平均值。不透明度通过公式(2)计算:
式中:A 为薄膜在600 nm 的吸光度(Abs);x 是薄膜的厚度(mm);结果表示为Abs/mm。
1.2.2.5 微观结构研究 样品裁成1 cm×4 cm 的条状,使用扫描电子显微镜(SEM),加速电压20 kV,使用液氮脆断薄膜,并在横截面上喷金后通过SEM 扫描观察横截面形貌。
1.2.2.6 水蒸气透过率测定 薄膜的水蒸气透过率用透湿杯法,在23 ℃、相对湿度90%±2%条件下进行,每隔12 h 将测试杯取出,精确称量测试杯质量的变化,通过公式(3)计算水蒸气透过率(WVP):
式中:W 为水增加量(g);x 为膜厚(mm);A 为膜面积(mm2);Δp 为水蒸气压差(Pa);t 为时间(s);水蒸汽压根据测试时的温度取值。
1.2.2.7 拉伸力学性能测试 薄膜需裁成条状试件规格为2 cm×16 cm,使用万能力学试验机根据国标《塑料 拉伸性能的测定 第三部分:薄膜和薄片的试验条件》 GB/T 1040.3-2006 进行测试。调试测试距离为50 mm,拉伸速率为50 mm/min,测试的环境温度为23 ℃,湿度45%~55%。每个样品重复5 次。
1.2.2.8 热稳定性 称取5~10 mg 薄膜碎片于坩埚内,使用热重分析仪进行测试,检测时采用氮气(N2)保护,测试温度范围30~600 ℃,升温速率为10 ℃/min。
1.2.2.9 抗氧化活性 薄膜的抗氧化活性通过DPPH自由基清除能力进行测定,将0.1 g 薄膜样品置于10 mL 的蒸馏水中搅拌30 min,所得水溶液备用。取4 mL 0.15 μmol/L 的DPPH 乙醇溶液与1 mL 制备好的水溶液,避光静置反应30 min。测量反应后的溶液在紫外分光光度计517 nm 处的吸光度值,重复三次试验取平均值[30]。自由基清除率(RSA)通过以下公式(4)计算:
式中:A1为样品水溶液在517 nm 处吸光度 (Abs);A0为乙醇水溶液在517 nm 处吸光度(Abs)。
1.3 数据处理
每项测定至少重复3 次,数据用均值±标准差表示,差异的显著性采用SPSS 26 检验,当P<0.05 时,均值被认为具有统计学意义,数据绘制软件采用Origin 2018。
2 结果与分析
2.1 杨梅单宁-酪蛋白复合膜可能存在的非共价键作用及红外光谱分析
酪蛋白在单宁的交联作用下如图1,交联的非共价键作用有氢键(主要)、疏水作用、离子键(次要),通常这三种作用是协同进行:单宁羟基与蛋白质中的羰基和肽键结合氢键;疏水键则是由单宁上的非极性芳香环与疏水性氨基酸蛋白质残基作用形成;离子键则是带正电荷的氨基基团与带负电荷的羟基基团作用形成[31]。
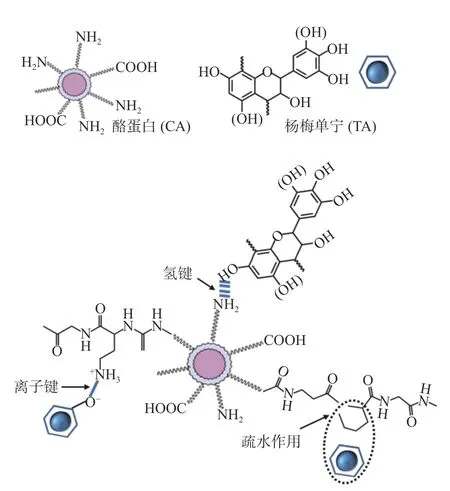
图1 杨梅单宁-酪蛋白复合膜可能存在的非共价键作用Fig.1 Cross-linked (non-covalent) interaction between Chinese bayberry tannin and casein
图2 为杨梅单宁改性酪蛋白膜的红外光谱图。在纯酪蛋白膜C1 的红外光谱中,位于3227 cm-1的宽峰为N-H 和O-H 的伸缩振动特征峰。2987 cm-1和2885 cm-1为亚甲基-CH2的伸缩振动峰。典型的蛋白质特征吸收带为:酰胺I 带(C=O 的伸缩振动峰)位于1638 cm-1;酰胺II 带(N-H 弯曲振动峰)位于1552~1558 cm-1;酰胺III 带(C-N 的伸缩振动峰和N-H 的弯曲振动峰)位于1236~1238 cm-1。经杨梅单宁改性后,BT-CA 复合膜的酰胺I、酰胺II 和酰胺III 带的峰谷位置都发生不同程度的偏移,这一现象与前人单宁改性酪蛋白膜、大豆分离蛋白膜是研究结果一致[9,32],研究结果推测单宁可能通过氢键、疏水键、离子键等化学键与酪蛋白反应,增强了酪蛋白分子链间的作用力。
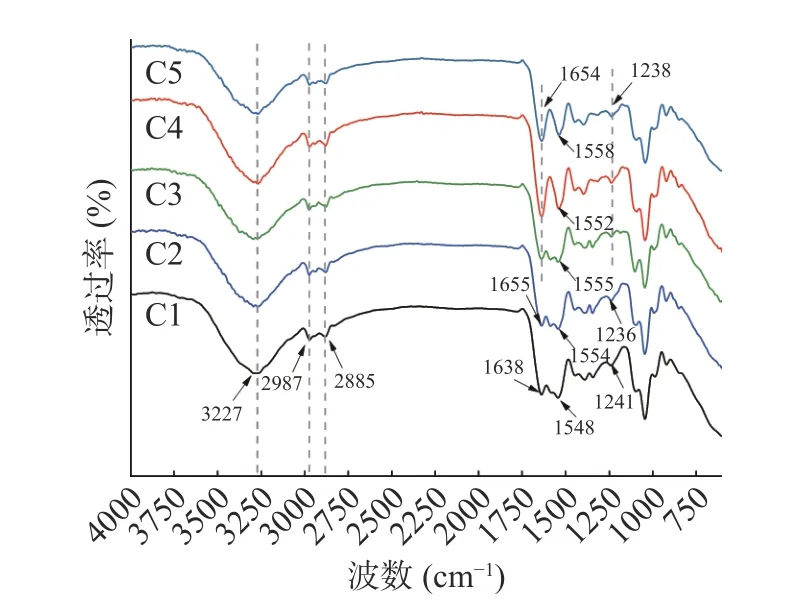
图2 杨梅单宁改性酪蛋白膜的FT-IR 光谱图Fig.2 The FT-IR spectroscopy of BT-CA films
2.2 BT-CA 复合膜的厚度、水溶性和不透明度
表2列出了不同比例杨梅单宁改性酪蛋白膜(BTCA 复合膜)的厚度和水溶性值。随着单宁的加入,成膜液的浓度略有提高,因此,BT-CA 复合膜的厚度有不同程度增加。BT-CA 复合膜的水溶性随着单宁含量的增加呈先减后增的趋势。与其它单宁交联改性蛋白膜的现象类似[9,21]:形成难溶于水的复合物,水溶性下降。然而随着单宁含量的增加,与酪蛋白以弱键连接或游离的单宁析出,导致水溶性增加。
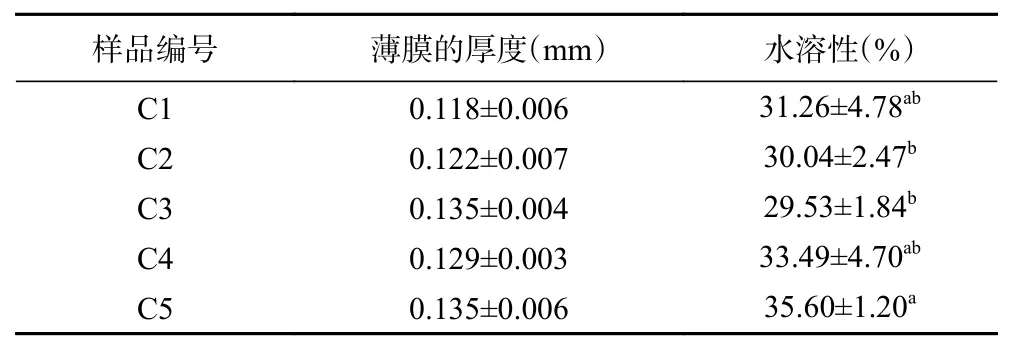
表2 杨梅单宁改性酪蛋白膜的厚度和水溶性Table 2 Thickness and water solubility of BT-CA films
如图3 所示,杨梅单宁改性酪蛋白膜不透明度的变化受到单宁添加量的影响。随着单宁量的增加,BT-CA 复合膜的不透明度显著升高。和纯蛋白膜C1 相比,BT-CA 复合膜会表现出更暗淡的颜色,薄膜颜色逐渐加深,这是因为:一方面,添加的酚类氧化成醌类物质,使薄膜颜色变暗、透明度下降;另一方面,添加酚类物质后,光会被交联结构散射或折射产生较暗沉的视觉感受,这与前人的研究结果相吻合[9]。不透明度的升高导致光线透过率降低,在具有避光性功能需求的包装薄膜领域有较好的应用前景。
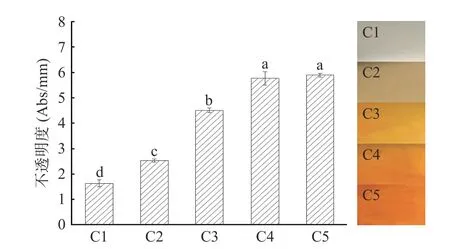
图3 杨梅单宁改性酪蛋白膜的不透明度和宏观图片Fig.3 The opacity and optical image of BT-CA films
2.3 拉伸性能
图4为杨梅单宁改性酪蛋白膜的拉伸强度及断裂伸长率。纯酪蛋白膜(C1)的拉伸强度为3.88 MPa,断裂伸长率为164.1%,图4 中可观察到BT-CA 复合膜(C2、C3、C4)的拉伸强度随单宁添加量增加而提高,其中C4 拉伸强度最佳为6.63 MPa,断裂伸长率稍微下降为140.4%,综合性能较好。这一现象产生的原因可能是单宁通过氢键、疏水键及可能存在的共价键,提高了酪蛋白膜分子链间的相互作用力,并且酪蛋白特有的开放结构有利于肽链上的羰基和多酚羟基间形成氢键,酪蛋白中的脯氨酸含量高,这种环状的亚氨基酸可以与单宁芳环间形成更多的疏水键,进而薄膜形成更为稳固的三维网状结构,因此薄膜强度得到提升、力学性能得到可观的改善[10]。与单宁酸改性酪蛋白的研究结果不同[9],在单宁添加量达20%时,拉伸强度下降,这可能由于缩合单宁过量,导致未与酪蛋白形成复合物的单宁填充在薄膜的内部结构中,影响酪蛋白成膜时的内聚力[33]。此外,杨梅单宁分子量较大、聚合度高,羟基间的氢键作用、芳环的π-π 堆积作用,使得过量的单宁更容易在酪蛋白基材中聚集,导致薄膜存在不连续结构,影响拉伸强度[30,34],这在后面的褶皱脆断面形貌特征中可以得到验证。单宁加入后,随着酪蛋白分子链间交联程度的提高,BT-CA 复合膜的断裂伸长率下降,而C5 断裂伸长率略有提高,可能是由于薄膜内部结构中存在过量的单宁类似于增强体,使得复合膜的断裂伸长率有所提高[35]。
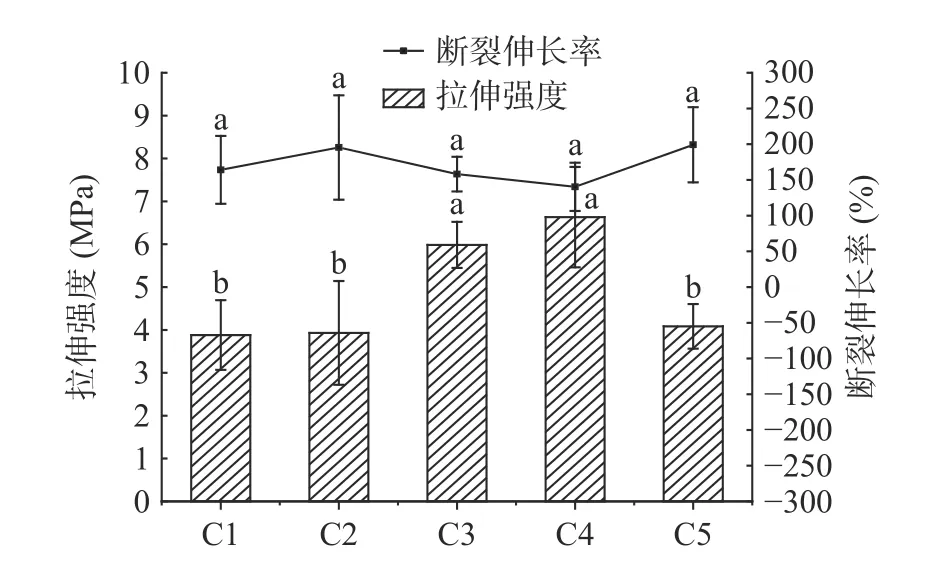
图4 杨梅单宁改性酪蛋白膜的拉伸强度及断裂伸长率Fig.4 Tensile strength and elongation at break of BT-CA films
2.4 微观形貌
图5为杨梅单宁改性酪蛋白膜在1000×(左图)和2000×(右图)放大倍数下的脆断横截面扫描电镜图。由图5 可见,杨梅单宁加入后,BT-CA 复合膜的脆断横截面保留了酪蛋白膜致密的内部结构,未出现分层现象。说明在成膜过程中,单宁与酪蛋白通过氢键、疏水键、共价键等化学键的交联作用与酪蛋白形成复合物,并在成膜过程中,依靠分子间作用力形成致密的微观结构,两者具有良好的相容性[36]。除C5外,BT-CA 复合膜的断面较为光滑,特别是C3 和C4的脆断横截面致密,说明10%~15%的杨梅单宁与酪蛋白间的交联作用及BT-CA 复合物间的分子间作用力最强,这与C3 和C4良好的拉伸性能相互印证。而C5 脆断截面呈褶皱状,这是由于单宁过量聚集在复合膜内部结构中,导致复合膜存在不连续的弱界面结构造成的[37]。

图5 杨梅单宁改性酪蛋白膜的脆断截面扫描电镜图Fig.5 SEM micrographs of fractured cross-sections for BT-CA films
2.5 水蒸气透过率
水蒸气透过率(WVP)指水蒸气可以通过薄膜扩散的能力,图6 中随着单宁添加量的增加,BT-CA 复合膜的水蒸气透过率下降。由图6 可见,仅5%的杨梅单宁即可减少酪蛋白膜的水蒸气透过率50%,说明单宁的交联作用可以显著提高酪蛋白膜的水蒸汽阻隔性能,这与许多单宁改性蛋白膜的研究结果类似[10,34]。从复合膜的扫描电镜图看,杨梅单宁与酪蛋白形成致密的微观结构可以抑制水蒸气透过薄膜,尤其是C4、C5 的表现更为优异,添加单宁量达15%以上时,过剩的单宁填充在酪蛋白膜的孔隙中,使得水分子透过薄膜的通道更为曲折,因此水分子透过薄膜的难度增加,进一步降低水蒸气透过率[34,38]。
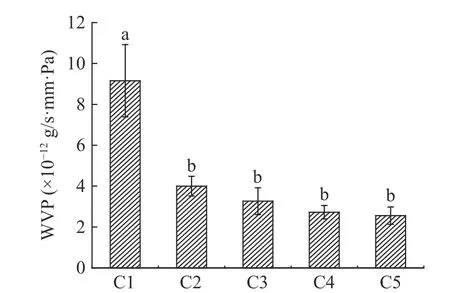
图6 杨梅单宁改性酪蛋白膜的水蒸汽透过率Fig.6 The water vapor permeability of BT-CA films
2.6 热稳定性
图7为杨梅单宁改性酪蛋白膜的热失重曲线图。由TG 曲线图可见,薄膜的热失重曲线可以分为三个阶段:30~100 ℃范围内的失重主要源于薄膜中吸附的游离水挥发;150~250 ℃范围内的失重主要为增塑剂丙三醇化合物与结合水的受热挥发;300~400 ℃范围内的失重是由于酪蛋白分子链断裂,薄膜稳定结构逐渐分解[9]。BT-CA 复合膜在第一阶段的失重率比纯酪蛋白膜低,说明单宁通过与酪蛋白分子链的交联作用降低了酪蛋白的亲水基团含量,并且交联结构提高了薄膜分子链间的作用力,降低了复合膜对环境中水分的吸附。从DTG 图可知,酪蛋白膜经过杨梅单宁改性后,复合膜第三阶段的最大失重速率温度后移,失重速率减缓,说明单宁的交联改性可以显著减缓酪蛋白分子链断裂,提高酪蛋白膜的热稳定性。
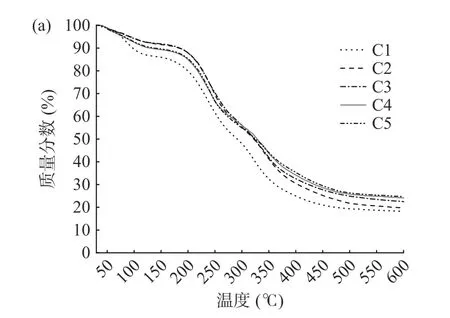
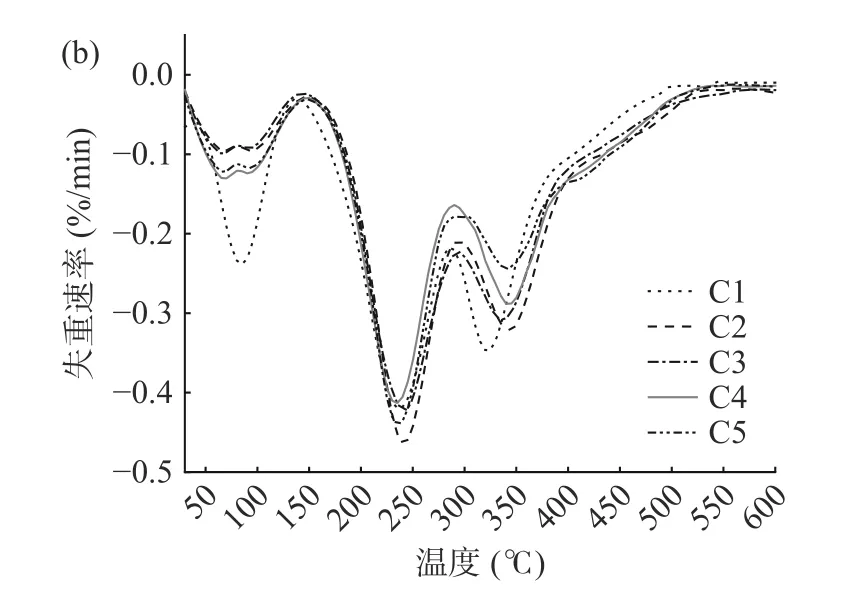
图7 杨梅单宁改性酪蛋白膜的TG 曲线(a)和DTG 曲线(b)Fig.7 TG curve (a) and DTG curve (b) of BT-CA films
2.7 抗氧化性能分析
图8为单宁改性酪蛋白膜的DPPH 自由基清除率,自由基清除率可以间接反应薄膜的抗氧化性能。由图8 可知,纯酪蛋白膜具有一定自由基清除能力,这是由于酪蛋白的肽链中含有一些可以与自由基反应的氨基酸[39]。C2 的自由基清除率比纯酪蛋白膜低,可能是碱性条件下,单宁的酚羟基氧化成醌,与酪蛋白肽链上的氨基反应,两者形成稳固的复合物[19],导致缺少捕捉自由基的氢供体,自由基清除率下降。随着杨梅单宁含量的提高(>5%),以弱连接键酪蛋白的单宁含量增加,单宁容易从薄膜中释放,因此,BTCA 复合膜的自由基清除能力增强。特别是C4 和C5,填充在复合膜中的过剩单宁遇水迅速溶解,从薄膜中释放,与自由基反应。
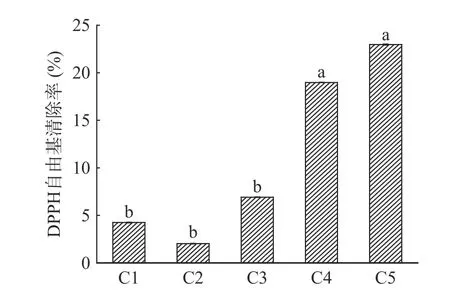
图8 杨梅单宁改性酪蛋白膜的DPPH 自由基清除率Fig.8 DPPH radical scavenging performance of BT-CA films
3 结论
将杨梅单宁作为交联剂,采用溶液流延法,制备了单宁改性酪蛋白复合膜。通过红外光谱分析发现单宁通过氢键、疏水键、离子键等化学键与酪蛋白反应,增强了酪蛋白分子链间的作用力。在拉伸力学测试中,当杨梅单宁的添加量为15%,复合膜拉伸强度达到最大值6.63 MPa,明显高于纯酪蛋白薄膜。热重分析、水蒸气透过率、不透明度和抗氧化性测试表明,单宁与酪蛋白的交联可以明显的提升酪蛋白复合膜的热稳定性和水蒸气阻隔能力、降低膜的透明度。根据观察SEM 电镜图,单宁交联改性后的复合膜具有致密的微观结构。综上所述,适量杨梅单宁的加入能改性酪蛋白膜且添加量在15%时复合膜的性能效果较佳,赋予膜材料一定的功能特性,因此在包装薄膜领域具有一定的应用潜力。
猜你喜欢
中国饲料(2022年5期)2022-04-26
——“单宁”
山东国资(2020年6期)2020-07-09
西南国防医药(2016年6期)2016-12-01
中国塑料(2016年1期)2016-05-17
中国酿造(2016年12期)2016-03-01
中国塑料(2015年3期)2015-11-27
中国塑料(2014年10期)2014-10-17
应用化工(2014年11期)2014-08-16
食品工业科技(2014年3期)2014-03-22
食品科学(2013年13期)2013-03-11
