
甘草多糖对断奶仔猪生长性能和生长基因表达量的影响
2023-08-14 06:51:37杨又兵李新月娄然刘永建任旭鸽曾凡林游祥宾李淦徐志谦雷莹白俊艳
江苏农业科学 2023年13期
杨又兵 李新月 娄然 刘永建 任旭鸽 曾凡林 游祥宾 李淦 徐志谦 雷莹 白俊艳


摘要:选取甘草多糖(GPS)为原料,主要研究甘草多糖对断奶仔猪生长性能、血清免疫指标和生长基因表达量的影响。结果表明,在对断奶仔猪生长性能影响中,800、1 500 mg/kg GPS处理组的ADG与对照组相比,差异显著;在对断奶仔猪腹泻率影响中,400、800、1 000 mg/kg GPS处理组和对照组相比,腹泻率显著下降;在对断奶仔猪血清免疫球蛋白中,1 000、15 000 g/kg GPS处理组与对照组相比,血清中IgG含量显著升高,800、1 500 mg/kg GPS处理组与对照组相比,血清中IgM含量显著升高;在断奶仔猪生长基因表达量中,肝脏中800 mg/kg GPS处理组的IGF-1基因mRNA表达量与对照组相比显著升高,背长肌中1 000 mg/kg GPS处理组的IGF-1基因mRNA表达量与对照组相比显著升高,800 mg/kg GPS处理组IGF-2基因的mRNA表达量与对照组相比显著升高(P<0.05)。
关键词:甘草多糖;生长性能;cDNA;基因表达量;断奶仔猪
中图分类号:S816.7 文献标志码:A
文章编号:1002-1302(2023)13-0182-07
甘草,或称光果甘草,是中药中“必不可少的草药”。甘草在我国的使用早于希腊和罗马帝国,有着悠久的传统药物和民间疗法历史[1]。甘草原产于南欧和亚洲部分地区,它被广泛用作草药和天然甜味剂。甘草被认为是最重要的6种草药之一,并且在联合应用时可以降低某些草药的毒性并提高其疗效,有一个经典的中医理论称“五方十有八九含甘草”[2]。在临床方面甘草也是一种重要中药,它含有20多种三萜类化合物和300多种黄酮类化合物[3]。甘草可以用于治疗多种疾病,如呼吸系统疾病、癫痫、发热、性衰弱、瘫痪、胃溃疡、风湿、皮肤病、出血性疾病、黄疸等,这与甘草自身的活性和药理作用有很大的联系[4]。和甘草相关的衍生物包括甘草酸、甘草次酸、甘草皂苷和甘草黄酮类中的甘草总黄酮、异甘草素、甘草苷、光甘草定、甘草多糖等,这些甘草衍生物都是近些年发现的从甘草中提取的活性成分[5-6]。
甘草多糖(glycyrrhiza polysaccharide,GPS)是一种酸性吡喃多糖,内部有小气泡状孔洞。相关研究表明,GPS因能增强免疫调节和抗肿瘤、抗氧化、抗病毒、抗菌、抗过敏、抗炎等重要生物活性而备受重视。GPS具有毒性小、易提取等优势,在动物产品上也可用做饲料添加剂,能够显著加强畜禽免疫调节功能,提高生长性能,有着广泛的应用前景。本试验主要研究了GPS作为饲料添加剂对断奶仔猪生长性能和免疫等方面指标的影响,旨在为健康养殖和畜牧业发展提供理论依据。
1 材料与方法
1.1 试验材料
甘草多糖原料购自河南省洛阳蓝斯利科技有限公司,饲料购自河南省洛阳市六和饲料有限责任公司。试验选用60头 健康、初始体质量为(8.33±0.39)kg/头的28日龄的杜洛克×长白×大白 三元杂交猪,公母各半。试验于2021年5—7月在河南省新郑市银发牧业猪场进行。
1.2 试验方法
1.2.1 样品采集 采取单因素设计,随机分为5个处理组,每个处理组包括3个重复,每个重复(栏)4头仔猪。预试期3 d,正试期28 d。试验组分别饲喂添加400、800、1 200、1 500 mg/kg GPS的试验日粮,对照组饲喂基础日粮。在每个重复中选取1头健康状况良好的断奶仔猪,空腹时进行前腔静脉采血 10 mL,并加入ACD抗凝血剂(ACD与血液的体积比为1 ∶5),将采集完成的血样放置在冰盒中,用记号笔编号,带回实验室进行离心并收集血清,用于后续的指标测定。断奶仔猪放血后进行屠宰,以组为单位,采集每个重复1头断奶仔猪的肝脏和背最长肌组织,并用无菌手术剪剪成小块(约1 cm3),放入冻存管中,用记号笔标注编号,随即装入液氮中,带回实验室保存待测。
1.2.2 生长性能指标测定 仔猪日采食量(average daily feed intake,ADFI):每天早晨在投料前对仔猪逐个称质量,并对前1 d仔猪所吃剩的料进行称质量,统计各组仔猪采食量,计算每头猪平均日采食量。仔猪平均日增质量(average daily gain,ADG):以每个组重复为单位,在试验开始进行至结束每天07:00对仔猪进行空腹称质量,并计算平均日增质量。料肉比F/G(Feed/Gain)=ADFI /ADG。
1.2.3 腹泻率测定 每天观察猪的健康状况,记录各栏猪的腹泻次数,计算各处理组的腹泻率。
腹泻率=(腹泻猪头数×猪腹泻天数)/(试验猪头数×试验天数)×100%
1.2.4 血清免疫指标测定 使用南京建成试剂盒,具体型号为免疫球蛋白IgA测定试剂盒E027-1-1、免疫球蛋白IgM测定试剂盒E025-1-1、免疫球蛋白IgG测定试剂盒H106-1-1,分别测定血清的IgA、IgG、IgM含量,并按照试剂盒中步骤进行操作。
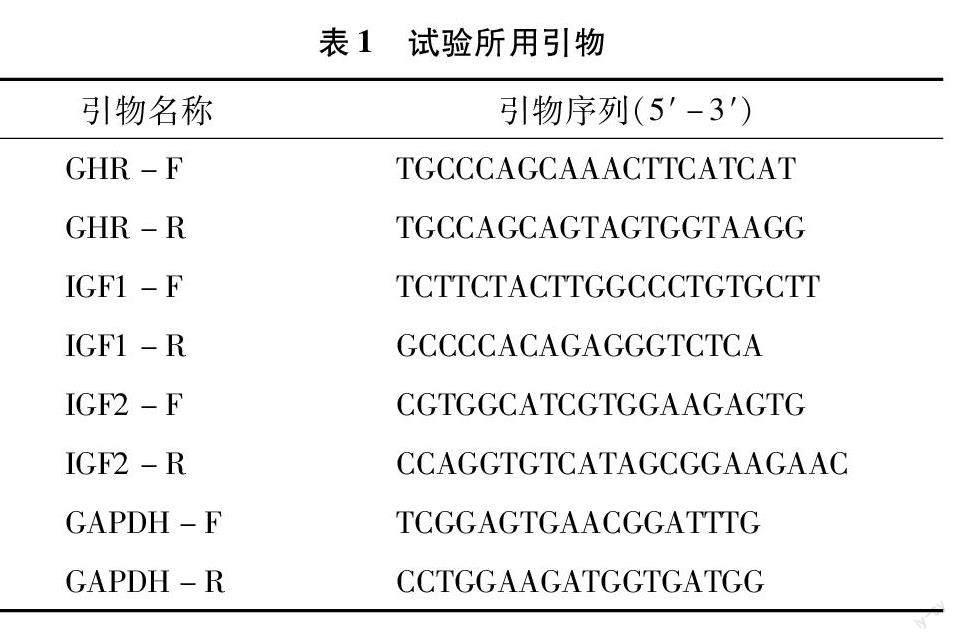
1.2.5 引物设计 引物由生工生物工程(上海)股份有限公司合成,具体引物信息见表1。
1.2.6 组织样RNA提取、检测 (1)从冻存管里取大约100 mg的组织样品,采用液氮充分碾磨后,用提前預冷的小勺把组织转入1.5 mL的RNase-free离心管中加入1 000 μL的RNAiso Plus(trizol)进行匀浆,室温静置5 min,10 000 g 4 ℃离心6 min。(2)取上清到新的1.5 mL的RNase-free离心管中,加入200 μL的三氯甲烷并振荡15 s,放置 10 min,期间不断摇晃,然后离心10 min。(3)取上清到新的1.5 mL的RNase-free离心管中,加入500 μL异丙醇,室温放置10 min,10 000 g离心 10 min,离心后底部管壁会产生白色RNA沉淀。(4)弃上清,保留白色沉淀物,加1 mL 75%乙醇,然后把离心管放入4 ℃、7 500 g的离心机内离心 5 min,弃上清,再次离心1 min,打开盖子晾干 5 min,乙醇易挥发,可以将离心管中的残留液体挥发带走。(5)往晾干的离心管内加入20 μL DEPC水,用枪头吹打混匀。(6)用超微量分光光度计检测,检测D260 nm/D280 nm值在1.8~2.0之间,表明RNA纯度较高,可以用于后续反转录和荧光定量试验。
1.2.7 cDNA合成 试验使用TaKaRa反转录试剂盒合成cDNA。反转录的体系(1)共10 μL,具体配制见表2。将体系(1)各分部试剂混合均匀后,放入PCR仪中,并按说明书程序进行设置,来进行反转录反应,具体设置为42 ℃ 2 min,4 ℃冷却。反转录体系(2)共20 μL,具体配制见表3。混匀体系(2)各部分试剂,按说明书进行反转录反应,设置为 37 ℃ 15 min,85 ℃ 5 s,4 ℃冷却。上述试操作都在冰盒上进行,合成的cDNA储存于-20 ℃。
1.2.8 实时荧光定量PCR 试验使用TaKaRa试剂盒,反转录的体系共20 μL,具体配制见表4。
1.2.9 数据处理 生产数据采用SPSS 25.0进行单因素方差分析,并用Duncan s法进行多重比较检验,结果以平均数±标准差表示。相对定量数据采用Excel 2010对数据进行整理,用2-ΔΔCT法进行计算。试验结果进行t检验分析其显著性,使用GraphPad Prism 8.0.1进行作图。*代表差异显著(P<0.05);**代表差异极显著(P<0.01)。
2 结果与分析
2.1 甘草多糖对断奶仔猪生长性能的影响
由表5可知,400、800、1 000、1 500 mg/kg GPS处理组与对照组相比,始质量、末质量、ADFI、F/G没有显著差异,但ADFI、ADG相比对照组有所升高;800、1 500 mg/kg GPS处理组的ADG与对照组相比,差异显著(P<0.05)。
2.2 甘草多糖对断奶仔猪腹泻率的影响
由表6可知,400、800、1 000 mg/kg GPS处理组的腹泻率和对照组相比,显著下降(P<0.05)。1 500 mg/kg GPS处理组也可以使腹泻率降低,但差异不显著。
2.3 甘草多糖对断奶仔猪血清免疫球蛋白的影响
由表7可知,试验各组血清中IgA含量没有显著差异;1 000、1 500 g/kg GPS处理组与对照组相比,血清中IgG含量显著升高(P<0.05);800、1 500 mg/kg GPS处理组与对照组相比,血清中IgM含量显著升高(P<0.05)。
2.4 甘草多糖对断奶仔猪生长基因表达量的影响
由图1可知,在肝脏中,GPS处理组和对照组相比,GHR基因和IGF-2基因的mRNA表达量差异不显著;800 mg/kg GPS处理组的IGF-1基因的mRNA表达量与对照组相比,显著升高(P<0.05);其余各处理组IGF-1基因的mRNA表达量与对照组相比,差异不显著。
由图2可知,在背长肌中,与对照组相比,GPS处理组GHR基因的mRNA表达量差异不显著;1 000 mg/kg GPS处理组的IGF-1基因的mRNA表达量显著升高(P<0.05);其余各处理组的IGF-1基因mRNA表达量差异不显著。800 mg/kg GPS处理组IGF-2基因的mRNA表达量与对照组相比显著升高(P<0.05),其余各处理组IGF-2基因的mRNA表达量与对照组相比差异不显著。
3 讨论
3.1 甘草多糖对断奶仔猪生长性能的影响
自1943年首次将多糖用作药物以来,随着生物化学的发展以及各种分离技术的研究和应用,多糖的复杂结构和多样性逐渐被人们所认识[7]。植物多糖对动物机体有诸多有益功效,能够改善畜禽的生长性能,提升动物机体的抗病能力,并且增进机体对营养物质的消化吸收和利用等[8-9]。在基础日粮中添加一定剂量的多糖可以提高仔猪ADFI、ADG,并降低F/G。Zhao等研究发现,添加桑叶多糖饲喂断奶仔猪,在低(0.3 g/kg)、中(0.6 g/kg)、高(0.9 g/kg)剂量组中的ADFI均高于对照组[10]。另有研究表明,在饲粮中添加酵母壁多糖和黄芪多糖、刺五加皂苷混合物,可以增高断奶仔猪的ADG、ADFI,并且有减弱F/G的趋势[11-12]。本试验结果表明,日粮中添加800、1 500 mg/kg的GPS可以显著提高断奶仔猪的ADG,而其余指标与各处理组之间没有明显差别,但添加GPS各剂量组的ADFI均高于对照组。据前人研究,在断奶仔猪饲粮中加入太子参茎叶多糖和白术多糖,可以使断奶仔猪的ADG增加;添加白术多糖还可以降低F/G[13-14],本试验结果与之相似。本试验各处理组GPS增加了断奶仔猪的ADFI,显著增加了ADG,并且在高剂量组中数值最大,生长性能基本呈剂量依赖性关系。总体而言,GPS有助于改善饲料的适口性和增加饲料的消化吸收率,提高断奶仔猪的生长性能。
3.2 甘草多糖对断奶仔猪腹泻率的影响
断奶是仔猪生活中营养方式与生存环境改变的转折点。仔猪腹泻目前仍是养猪业需要关注的重点问题,防控不到位会造成巨大损失;伴随着胃肠结构、肠道功能和免疫系统的明显变化,断奶后仔猪肠道疾病和腹泻的发病率较高,生长迟缓[15-16]。在實际生产过程中,还可能由于一些致病菌的侵入,导致猪的机体出现腹泻等症状。多糖对降低断奶仔猪的腹泻率有良好的作用效果,在断奶仔猪饲粮中单独添加黄芪多糖、白术多糖和牛膝多糖,可以明显改善断奶仔猪发生腹泻的情况,3种多糖类添加剂联合使用也有较好的功效[17]。刘佳等在饲粮中添加酵母多糖,可以抑制断奶仔猪发生腹泻的情况,添加0.2%酵母多糖作用效果最好[18]。另有研究发现,白术多糖与上述多糖作用效果类似,并且仔猪在14~28日龄时,添加 9 g/kg 白术多糖腹泻率最低[19]。本试验结果显示,添加400、800、1 000 mg/kg GPS,可以明显控制断奶仔猪腹泻情况的发生,与其他多糖类饲养试验结果相同;添加 400 mg/kg GPS时,仔猪腹泻率最低,随着GPS剂量的增加,仔猪腹泻率有所升高,这可能是由断奶仔猪胃肠消化及吸收能力的下降而引起;但相比于对照组,GPS处理组的腹泻率显著性减少。本试验说明,在饲粮中添加GPS可以改善仔猪的腹泻率,在添加剂量为400 mg/kg时,效果最佳。
3.3 甘草多糖对断奶仔猪血清中免疫球蛋白的影响
近年来,随着畜牧业绿色健康养殖的发展,人们对营养调控和免疫的联系更加关注。许多研究表明,良好的营养条件可以增强动物机体的免疫防御能力,促进机体的免疫活动[20]。免疫球蛋白是一种由抗原刺激动物体内产生的球蛋白,它具有结合抗原、刺激补体产生等生物学功能,普遍存在于血液、组织液和外分泌液中[21]。其中,IgA、IgG、IgM普遍存在于各种哺乳动物的血清中,形成反映机体体液免疫的主要指标。因此,在断奶仔猪阶段,IgA、IgG、IgM含量的提高可以在一定程度上减少疾病的发生[22]。科学研究已证实,众多植物多糖可产生免疫调控作用,是天然的免疫调节剂,并能够活化T、B淋巴细胞,提高动物机体血清中的抗体水平,增强免疫调节[23]。有研究称,在日粮中添加高剂量浓度(0.08%)的决明子多糖,能够使血清中IgA、IgM、IgG的含量明显提高[24]。Chen等在断奶仔猪日粮中添加1 000、1 500 mg/kg牛膝多糖,仔猪在第14天和第28天的血清中IgG、IgA、IgM含量明显增多[25]。本试验结果表明,添加800、1 500 mg/kg GPS,可以使仔豬血清中IgM含量显著升高;添加 1 000、1 500 mg/kg GPS,可以使仔猪血清中IgG含量显著升高。胡菁等研究发现,GPS可以增加小鼠血清中IgG、IgM浓度,在600 mg/kg浓度时有显著性的增加,从而增强机体的体液免疫[26],本试验研究结论与之一致。表明GPS能够增加血清中免疫球蛋白浓度,提高动物机体的体液免疫。
3.4 甘草多糖对断奶仔猪生长基因表达量的影响
生长激素受体(GHR)基因与GH基因有很高的关联度,GHR可以调控GH基因的表达,也是生长轴的重要组成部分,被认为是产后体细胞生长的主要调节因子,在刺激细胞分裂、骨骼生长、蛋白质合成等过程中有所影响,在动物的生长发育中起着重要作用[27-28]。GH发挥作用的前提需要与GHR结合;如果GHR不足,GH发挥作用也会受到抑制。以往认为GHR仅在鱼类和哺乳动物的肝脏中表达,介导IGF1的释放以响应GH结合;另有数据表明,GHR也在外周组织中表达,局部调节GH的促生长作用[29]。不同的营养水平下,不同物种GHR基因的mRNA表达量也有所不同。有研究表明,在饲粮中添加复方中药制剂,能够提高仔猪胸腺、脾脏GHR基因的mRNA表达量[30]。Kareem等研究发现,在饲料中添加益生元和菊糖,对肉鸡肝脏中GHR基因的mRNA表达量有所升高[31]。本试验结果发现,添加不同剂量的GPS,肌肉中GHR基因的mRNA表达量与对照组相比没有显著差异,但添加GPS后各组的表达水平有所提高;在添加 800 mg/kg 剂量GPS时,GHR基因的mRNA表达量最高。Gasparino等发现,在高饲料效率(FE)肉鹌鹑肝脏和肌肉中IGF-1的mRNA表达量较高,肌肉中GHR基因mRNA表达量较高[32]。饲喂玉米须粉和非淀粉多糖酶提高了仔猪的生长激素受体(GHR)和胰岛素生长因子(IGF)的mRNA相对表达量[33]。但本试验中,肝脏和背最长肌中,各剂量组和对照组相比,GHR基因的mRNA表达量没有显著性的提高,但800、1 000 mg/kg GPS处理组与对照组相比,GHR基因的mRNA表达量有升高趋势,这表明添加适当浓度的GPS对动物机体GHR基因的表达有上调作用。综上所述,在添加 800 mg/kg 剂量的GPS时,GHR基因表达量最高,效果最佳。
IGF-1可以通过调节生长激素的多种作用,参与发育、生长、生殖、代谢相关的关键生物学功能[34-36]。安静等分离培养绵羊原代成肌细胞,用重组慢病毒感染细胞等过程绘制细胞生长曲线,发现IGF-1能够促进绵羊成肌细胞的增殖[37]。同时,IGF-1也能够促进兔脂肪干细胞的生长和成骨分化,并且呈量效依赖性关系[38]。以上研究说明,IGF-1 最典型的功能之一是促生长作用,可以促进细胞的增殖和分化,进而来调节细胞的有丝分裂。本试验研究表明,添加800 mg/kg GPS后,肝脏中IGF-1基因mRNA表达量与对照组相比显著升高;添加1 000 mg/kg的GPS后,背最长肌中IGF-1基因的mRNA表达量与对照组相比显著升高。有研究称,在断奶仔猪饲粮中添加桑叶多糖,肝脏和背最长肌组织中IGF-1基因的mRNA表达水平都有了明显提高,低剂量组的背最长肌中IGF-1基因的mRNA表达量显著高于抗生素组,本试验结果与之相似;通过试验发现IGF-1基因mRNA表达量总体呈量效依赖性关系,在表达量达到最高后相对下降[39]。文贵辉等利用白术多糖饲喂樱桃谷鸭发现,白术多糖可以提高樱桃谷鸭肌肉中IGF-1基因的表达量[40]。本试验结果表明,GPS有一定的促生长作用,可能是由于GPS可以促进组织中 IGF-1 基因的mRNA表达量,进而提高细胞的增殖和分裂,促进动物的生长发育,但其具体机制还需要进一步验证。
IGF-2是一种小分子单链结构的肽类物质(分子量7 471 u,67个氨基酸残基),在胎儿发育和产后生长发育中起着主要作用,因此也称为胚胎样生长因子[41],在猪体内位于2号染色体上。研究表明IGF-2对牛骨骼肌卫星细胞的生长具有促进作用[42]。郭玉姣等利用荧光定量PCR技术,在长白猪和太湖猪身上发现IGF-2与早期脂肪细胞的增殖分化有关[43]。以上结果说明IGF-2在肌肉生长和分化过程中和脂肪性状方面也扮演着重要的作用。IGF-2基因在不同猪种不同组织中的表达量也有些许差异,朱晓峰等表明IGF-2基因在从江香猪肝脏和肺脏中的表达量最高,而在大白猪背最长肌和心脏的表达量均超过其他组织[44]。本研究结果表明,在肝脏中,各剂量组与对照组相比,IGF-2基因的mRNA表达量差异不显著。在背长肌中,添加800 mg/kg GPS,IGF-2基因mRNA表达量显著升高;在添加剂量为800 mg/kg时,IGF-2基因的表达量最高,在此浓度基础上增加剂量而表达量逐渐降低。而张冰等研究发现,IGF-2基因对猪的平均日增质量影响显著,与前面结果发现添加800、1 500 mg/kg GPS对断奶仔猪平均日增质量影响有显著差异结果相似,说明不同剂量的GPS可以调节IGF-2基因的表达,可能是由于IGF-2促进机体的骨骼肌细胞增殖分裂,提高了断奶仔猪的生长发育[45]。
4 结论
日粮中添加不同剂量的GPS能够增加断奶仔猪的日采食量,添加800、1 500 mg/kg浓度的GPS能够明显增加断奶仔猪的平均日增质量,400、800 mg/kg 低剂量组可以降低料肉比,但差异不明显。说明GPS在一定程度上可以提高断奶仔猪的生长性能,添加剂量在800 mg/kg时,效果最佳。日粮中添加400、800、1 500 mg/kg GPS,能显著降低断奶仔猪的腹泻率,添加400 mg/kg GPS效果最佳。日粮中添加1 000、1 500 mg/kg GPS对IgG含量有显著性提高;添加800、1 500 mg/kg GPS对IgM含量有显著性提高,但各试验组之间IgA含量没有显著差异。日粮中添加GPS在一定程度上可以上调GHR、IGF-1、IGF-2基因mRNA的表达量并有一定的显著性关系,对提高动物生长性能具有正向调控作用。
参考文献:
[1]Pastorino G,Cornara L,Soares S,et al. Liquorice (Glycyrrhiza glabra):a phytochemical and pharmacological review[J]. Phytotherapy Research,2018,32(12):2323-2339.
[2]Jiang M,Zhao S,Yang S,et al. An “essential herbal medicine”-licorice:a review of phytochemicals and its effects in combination preparations[J]. J Ethnopharmacol,2020,249:112439.
[3]Wang D N,Liang J,Zhang J,et al. Natural chalcones in Chinese materia Medica:licorice[J]. Evidence-Based Complementary and Alternative Medicine,2020,2020:3821248.
[4]El-Saber B G,Magdy B A,El-Mleeh A,et al. Traditional uses,bioactive chemical constituents,and pharmacological and toxicological activities of Glycyrrhiza glabra L.(Fabaceae)[J]. Biomolecules,2020,10(3):352.
[5]Karahan F,Avsar C,Ozyigit I I,et al. Antimicrobial and antioxidant activities of medicinal plant Glycyrrhiza glabra var. glandulifera from different habitats[J]. Biotechnology & Biotechnological Equipment,2016,30(4):797-804.
[6]李 想,李 冀. 甘草提取物活性成分藥理作用研究进展[J]. 江苏中医药,2019,51(5):81-86.
[7]Huang H L,Huang G L.Extraction,separation,modification,structural characterization,and antioxidant activity of plant polysaccharides[J]. Chemical Biology & Drug Design,2020,96(5):1209-1222.
[8]Liu W,Liu Y M,Zhu R,et al. Structure characterization,chemical and enzymatic degradation,and chain conformation of an acidic polysaccharide from Lycium barbarum L.[J]. Carbohydrate Polymers,2016,147:114-124.
[9]杨 玲,胡睿智,夏嗣廷,等. 植物多糖的功能性研究进展及其在动物生产中的应用[J]. 动物营养学报,2019,31(6):2534-2543.
[10]Zhao X J,Li L,Luo Q L,et al. Effects of mulberry (Morus alba L.) leaf polysaccharides on growth performance,diarrhea,blood parameters,and gut microbiota of early-weanling pigs[J]. Livestock Science,2015,177:88-94.
[11]贺 琴,王自蕊,游金明,等. 酵母壁多糖对断奶仔猪生长性能和小肠黏膜形态结构的影响[J]. 动物营养学报,2016,28(11):3536-3541.
[12]Kang P,Xiao H,Hou Y Q,et al. Effects of Astragalus polysaccharides,Achyranthes bidentata polysaccharides,and Acantbepanax senticosus saponin on the performance and immunity in weaned pigs[J]. Asian-Australasian Journal of Animal Sciences,2010,23:750-756.
[13]蔡旭滨,陈凌锋,檀新珠,等. 太子参茎叶多糖对断奶仔猪生长性能和血清抗氧化指标、免疫指标及生化指标的影响[J]. 动物营养学报,2016,28(12):147-154.
[14]Li L L,Yin F G,Zhang B,et al. Dietary supplementation with Atractylodes Macrophala Koidz polysaccharides ameliorate metabolic status and improve immune function in early-weaned pigs[J]. Livestock Science,2011,142(1/2/3):33-41.
[15]Heo J M,Opapeju F O,Pluske J R,et al. Gastrointestinal health and function in weaned pigs:a review of feeding strategies to control post-weaning diarrhoea without using in-feed antimicrobial compounds[J]. Journal of Animal Physiology and Animal Nutrition,2013,97(2):207-237.
[16]Jayaraman B,Nyachoti C M.Husbandry practices and gut health outcomes in weaned piglets:a review[J]. Animal Nutrition,2017,3(3):205-211.
[17]谢红兵,邹 云,刘丽莉,等. 植物多糖对断奶仔猪生长性能及肠道内环境的影响[J]. 动物营养学报,2018,30(7):2662-2671.
[18]刘 佳,何书海,朱忠珂,等. 酵母多糖对仔猪生长性能、免疫力、抗氧化及肠道健康的影响[J]. 饲料研究,2021,44(14):38-41.
[19]Li L L,Wu X,Peng H Z,et al. The effect of dietary addition of a polysaccharide from Atractylodes macrophala Koidz on growth performance,immunoglobulin concentration and IL-1β expression in weaned piglets[J]. The Journal of Agricultural Science,2009,147(5):625-631.
[20]皇甫冰. 浅谈幼龄动物的营养与免疫[J]. 中国畜牧业,2022(15):126-127.
[21]Wu H Z,Yang J,Wang S B,et al. Effects of soybean isoflavone and Astragalus polysaccharide mixture on colostrum components,serum antioxidant,immune and hormone levels of lactating sows[J]. Animals,2021,11(1):132.
[22]Wu Y,Pan L,Shang Q H,et al. Effects of isomalto-oligosaccharides as potential prebiotics on performance,immune function and gut microbiota in weaned pigs[J]. Animal Feed Science and Technology,2017,230:126-135.
[23]尚慶辉,解玉怀,张桂国,等. 植物多糖的免疫调节作用及其机制研究进展[J]. 动物营养学报,2015,27(1):49-58.
[24]Deng Z Y,Zhang J W,Wu G Y,et al. Dietary supplementation with polysaccharides from semen cassiae enhances immunoglobulin production and interleukin gene expression in early-weaned piglets[J]. Journal of the Science of Food and Agriculture,2007,87(10):1868-1873.
[25]Chen Q H,Liu Z Y,He J H.Achyranthes bidentata polysaccharide enhances immune response in weaned piglets[J]. Immunopharmacology and Immunotoxicology,2009,31(2):253-260.
[26]胡 菁,敖明章,崔永明,等. 甘草多糖的抗肿瘤活性及对免疫功能的影响[J]. 天然产物研究与开发,2008,20(5):911-913,38.
[27]Sharma A,Dutt G,Sivalingam J,et al. Novel SNPs in IGF1,GHR and IGFBP-3 genes reveal significant association with growth traits in Indian goat breeds[J]. Small Ruminant Research,2013,115(1/2/3):7-14.
[28]Chen C,Zheng Y,Wang M L,et al. SINE insertion in the intron of pig GHR may decrease its expression by acting as a repressor[J]. Animals,2021,11(7):1871.
[29]Bertucci J I,Blanco A M,Canosa L F,et al. Direct actions of macronutrient components on goldfish hepatopancreas in vitro to modulate the expression of ghr-Ⅰ,ghr-Ⅱ,igf-Ⅰ and igf-Ⅱ mRNAs[J]. General and Comparative Endocrinology,2017,250:1-8.
[30]刘 莉,郝福星,冒海银. 四君子汤对仔猪胸腺及脾脏生长激素受体mRNA表达的影响[J]. 动物医学进展,2009,30(6):66-69.
[31]Kareem K Y,Loh T C,Foo H L,et al. Effects of dietary postbiotic and inulin on growth performance,IGF1 and GHR mRNA expression,faecal microbiota and volatile fatty acids in broilers[J]. BMC Veterinary Research,2016,12(1):163.
[32]Gasparino E,del Vesco A P,Voltolini D M,et al. The effect of heat stress on GHR,IGF-I,ANT,UCP and COXⅢ mRNA expression in the liver and muscle of high and low feed efficiency female quail[J]. British Poultry Science,2014,55(4):466-473.
[33]Kirrella A A,Abdo S E,El Naggar K,et al. Use of corn silk meal in broiler diet:effect on growth performance,blood biochemistry,immunological responses,and growth-related gene expression[J]. Animals,2021,11(4):1170.
[34]Liu J L,LeRoith D.Insulin-like growth factor I is essential for postnatal growth in response to growth hormone[J]. Endocrinology,1999,140(11):5178-5184.
[35]Fontanesi L,Scotti E,Buttazzoni L,et al. Analysis of association between a microsatellite at intron 1 of the insulin-like growth factor 1 (IGF1) gene and fat deposition,meat production and quality traits in Italian large white and Italian duroc pigs[J]. Italian Journal of Animal Science,2013,12(3):e72.
[36]Buonomo F C,Lauterio T J,Baile C A,et al. Determination of insulin-like growth factor 1 (IGF1) and IGF binding protein levels in swine[J]. Domestic Animal Endocrinology,1987,4(1):23-31.
[37]安 靜,张雪梅,刘晨曦,等. 绵羊IGF-1基因慢病毒表达载体的构建及其促肌细胞生长作用研究[J]. 西北农林科技大学学报(自然科学版),2014,42(4):15-21.
[38]朱思斌,张 雪,江 飞,等. IGF-1对兔脂肪干细胞增殖和成骨分化的影响[J]. 安徽医科大学学报,2019,54(12):1913-1917.
[39]Zhao X J,Yang R L,Bi Y H,et al. Effects of dietary supplementation with mulberry (Morus alba L.) leaf polysaccharides on immune parameters of weanling pigs[J]. Animals,2019,10(1):35.
[40]文贵辉,李丽立,张 彬,等. 白术粗多糖对樱桃谷鸭IGF-1基因表达的影响[J]. 中国畜牧杂志,2011,47(13):63-65.
[41]Liu X F,Liu H B,Wang M,et al. Disruption of the ZBED6 binding site in intron 3 of IGF2 by CRISPR/Cas9 leads to enhanced muscle
development in Liang Guang Small Spotted pigs[J]. Transgenic Research,2019,28(1):141-150.
[42]孙晓丽,李树峰,佟慧丽,等. 不同肌肉特异性启动子IGF2表达载体构建及对牛骨骼肌卫星细胞增殖的影响[J]. 畜牧兽医学报,2015,46(4):555-560.
[43]郭玉姣,唐国庆,李学伟,等. 猪脂肪组织中IGF2和IGFBP3基因表达的发育性变化及其品种差异[J]. 遗传,2008,30(5):602-606.
[44]朱晓锋,许厚强,陈 伟,等. 从江香猪IGF-1和IGF-2基因的克隆、表达及生物信息学分析[J]. 农业生物技术学报,2019,27(8):1382-1391.
[45]张 冰,马青艳,覃 倩,等. 陆川猪和大白猪IGF2基因对生长性状的效应分析[C]. 广西畜牧兽医学会养猪分会2014年年会暨学术报告会论文集. 南宁,2014.
猜你喜欢
湖北农业科学(2017年3期)2017-03-21 18:48:04
湖北农业科学(2016年24期)2017-03-18 00:40:45
湖北农业科学(2016年18期)2016-12-08 18:27:31
河北渔业(2016年10期)2016-11-22 10:13:19
湖北畜牧兽医(2016年6期)2016-09-28 05:18:29
湖北畜牧兽医(2016年1期)2016-03-26 04:55:36
湖北农业科学(2015年19期)2015-10-28 22:17:03
吉林农业(2015年6期)2015-10-16 17:06:12
湖北农业科学(2015年18期)2015-10-13 22:19:41
江苏农业科学(2015年8期)2015-09-10 19:35:54
