
不同种植年限及覆膜栽培防风根际土壤微生物多样性研究
2023-08-14 07:24:35寇佩雯刘长乐李铂王楠宋忠兴张永生黄文静唐志书
江苏农业科学 2023年13期
寇佩雯 刘长乐 李铂 王楠 宋忠兴 张永生 黄文静 唐志书
摘要:为探究防风生长发育及栽培方式对其根际微生物群落结构的影响,采用Illumina高通量测序技术对非根际土壤(CK)、1年生(F1)、2年生(F2)和覆膜栽培1年(FM)的防风根际土壤中细菌16SrRNA和真菌18S序列进行测序分析。研究发现各处理的微生物总数、菌群丰富度和复杂程度表现为1年生组>非根际土壤组,1年生组>2年生组、覆膜组。覆膜栽培后微生物α多样性以及小囊菌科和丛赤壳科真菌丰度整体下降,假球壳科真菌丰度急剧增加。各处理土壤样品中微生物的群落组成虽存在差异,但其优势菌门相对稳定,细菌丰度最大的2个门始终是放线菌门和变形菌门,真菌丰度最大的门为子囊菌门和担子菌门。Venn分析和UPGMA聚类分析显示,F2处理中独有细菌的OTU数少于F1处理,独有真菌的OTU数则多于F1处理,F2和FM处理在菌落结构组成上较为相近。Spearman相关性分析发现,土壤全钾、硝态氮和铵态氮含量是驱动根际微生物群落的主要因子,但其与各优势菌群的相关性不显著。利用PICRUSt2进行菌群功能预测发现,2年生组较其余组菌群功能差异最大。综上可知,生长年限和覆膜均会影响防风土壤细菌群落结构和生态功能,而生长年限对其影响更加明显,该结果可为防风栽培技术体系的构建提供科学参考。
关键词:防风;根际土壤;微生物群落;多样性;功能预测
中图分类号:S182 文献标志码:A
文章编号:1002-1302(2023)13-0221-11
根际是植物-微生物-土壤三者相互作用并紧密影响的局域生态系统,其在生物和化学特性上与周围的大块土壤不同[1]。在此系统中,根际微生物参与了众多生化反应,在促进植物矿质元素吸收、刺激根系生长、加速土壤腐殖质形成和有机物分解等方面起到至关重要的作用[2-3]。因此,根际微生物常被认为是衡量土壤肥力和土壤健康的生物学指标,被誉为“植物的第二个基因组”[4]。植物在生长发育过程中所产生的萜类、黄酮和生物碱等次生代谢物常通过根系的分泌作用释放至土壤中,在改变根际土壤理化性质的同时也影响着根际微生物群落的结构特征[5]。研究发现,根际土壤微生物群落结构主要受2方面因素影响:在生物因素中,以植物种类、生长阶段和次生代谢差异[6-7]为主;在非生物因素中,土壤类型[8]、矿质元素[9]、温度及栽培方式[10]等对其影响较大。近年来,细菌 16S rRNA 和真菌 18S基因测序技术不断发展,利用第三代Illumina 高通量测序系统可同时对样品中的微生物进行检测并获得其生物群落结构特征。该方法获得的数据量大,能够产生覆盖深度较高的测序数据用来估计微生物群落的组成,由于其时间短和精准度高等特点,在微生物群落多样性研究等方面的应用越来越多。
药材防风为伞形科植物防风[Saposhnikovia divaricata (Turcz.) Schischk.]的干燥根,具有解表祛风、胜湿、止痉等作用,在我国历代本草文献中均有详细记载,属常用大宗药材[11]。现已从防风中得到色原酮、香豆素、多糖和挥发油类成分,临床常用其治疗风湿、风疹、炎症引起的急症,外感风寒引起的感冒头疼和脾胃等方面的相关疾病,因市场需求量大,现流通的防风多为人工栽培品[12]。内蒙古东部地区是目前我国防风的主产区之一,通过遥感影像解译防风的分布范围及种植面积发现,仅2019年内蒙古通辽市奈曼旗的防风种植面积就超过 35.33 hm2[13]。在之后的2年中通辽市科尔沁区及开鲁县的防风种植面积也在不断扩大,目前全市的防风在地面积预计已超666.67 hm2。种植防风通常2年采收,但在其主产区也有栽培1年便收获的现象,而不同生长年份的防风在根生物产量及有效成分积累上有一定差异[14-15]。此外,蒙东地区冬季冻土期长,常年昼夜温差较大,春季干旱少雨且时常伴有大风,土壤肥力低、沙化现象较为严重。因此,確定合理的栽培制度及采取相关保墒、提温的栽培措施对于确保防风的产量与质量显得至关重要。本研究以1年生、2年生和覆膜栽培的防风根际土壤为样本,通过高通量测序技术对根际细菌和真菌多样性的变化进行系统分析,探讨防风生长发育、栽培措施以及土壤理化性质对其根际微生物群落组成和多样性的影响,旨在为防风栽培技术体系的建立提供科学建议。
1 材料与方法
1.1 根际土壤样品的采集与处理
土壤样品于2021年9月11日采自内蒙古通辽市科尔沁区庆和镇林场3号地的防风试验田(121°55′17″E、43°43′56″N,海拔140 m),前茬作物为玉米。采用多点混合取样法,分别采集从未种植过防风的非根际土壤(CK,距防风试验田采样点 50 m 左右)和1年生(F1)、2年生(F2)、覆黑膜栽培1年(FM)的防风根际土壤。每种处理在其小区随机选3个取样点,每个取样点选取生长健康、大小一致的防风5株,除去落叶和表层土(3 cm)后,收集附着于根系表面0~0.5 cm的土壤作为根际土。将混匀后的土壤分为2份,一份装入无菌自封袋中经液氮速冻后放入干冰盒,由上海派森诺生物科技股份有限公司进行根际土壤分离、DNA提取和高通量测序,剩余土壤称量后带回实验室风干,研磨过筛后进行土壤含水量(WC)、pH值、总有机碳(TOC)、全氮(TN)、全磷(TP)、全钾(TK)、硝态氮(N)和铵态氮(A)等土壤理化因子的测定。
1.2 土壤微生物DNA提取、PCR扩增和文库制备
采用土壤DNA提取试剂盒(FastDNA SPIN Kit for Soil)提取总DNA,使用Nanodrop 2000对DNA进行定量,并通过1.2%琼脂糖凝胶电泳检测DNA提取质量。PCR扩增采用全式金公司的Pfu高保真DNA聚合酶进行,利用通用引物338F(5′-ACTCCTACGGGAGGCAGCAG-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)对细菌 V3-V4 区进行扩增;采用SSU0817F(5′-TTAGCATGGAATAATRRAATAGGA-3′)和1196R(5′-TCTGGACCTGGTGAGTTTCC-3′)对真菌V5-V7区进行扩增。扩增产物经磁珠纯化回收后采用Quant-iTTM PicoGreenTM dsDNA Assay Kit进行荧光定量检测,根据荧光定量结果,按照每个样本的测序量需求,对各样本按相应比例等摩尔混合。混合后的扩增产物以Illumina公司的TruSeq Nano DNA LT Library Prep Kit制备测序文库。
1.3 高通量测序及土壤微生物组成分析
对制备好的文库在Agilent Bioanalyzer上进行质检,合格的文库采用Illumina MiSeq测序仪进行双端测序,原始下机数据根据序列质量进行初步筛查后按照index和Barcode信息,进行文库和样本划分。按照QIIME2 dada2分析流程和Vsearch软件的分析流程进行序列去噪及操作分类单元(operational taxonomic unit,OTUs)聚类,对比Silva数据库(Release132,http://www.arb-silva.de)进行物种分类学注释,并采用RDP classifier贝叶斯算法对在97%相似水平下的OTU进行丰度分析,揭示样品的物种组成。根据OTU在不同样本中的分布,评估每个样本的α多样性水平,包括覆盖度(Coverage指数)、丰富度(Chaol指数和ACE指数)和香农指数(Shannon指数)等分析。
1.3 根际土壤理化性质测定
土壤理化性质测定按照《土壤农化分析》[16]进行:土壤含水量采用称重法测定;土壤pH值使用浸提法利用ORP-3000酸度计测定;土壤有机碳含量采用重铬酸钾水合加热法测定;全氮含量采用浓硫酸消解-凯式定氮法测定;全磷含量采用钼锑显色法测定;全钾含量采用火焰光度法测定;硝态氮含量采用酚二磺酸比色法测定;氨态氮含量采用KCl浸提-靛酚蓝比色法测定。
1.4 数据统计与分析
使用SPSS 18.0软件对数据进行单因素方差分析(ANOVA),采用新复极差法(Duncans)比较数据组间差异。使用R语言包vegan等进行群落Alpha多样性分析、PICRUSt2(phylogenetic investigation of communities by reconstruction of unobserved states)分析和主成分分析(principal co-ordinates analysis,PCoA) 等分析,并利用Excel 2013对数据进行整理和制表。采用R语言stat包的uclust函数计算β多样性距离矩阵,计算结果以平均连接聚类法 (unweighted pair group method with arithmetic mean,UPGMA)进行分析。物种差异分析使用VennDiagram包进行,所有结果采用R脚本进行可视化。采用Spearman相关系数对各个指标进行相关性分析,将获得的数值矩阵通过Heatmap图直观展示。
2 结果与分析
2.1 防风根际土壤微生物群落多样性分析
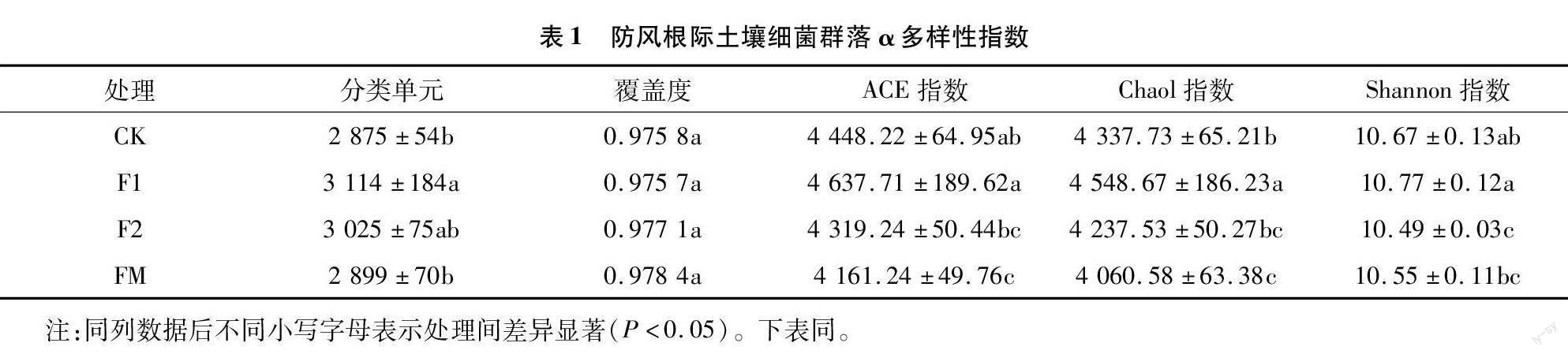
对4种处理下的12份土壤样品进行细菌16S rRNA高通量测序,共得到有效序列794 495条,对其进行聚类共获得属水平上的OTU样本35 745个。由表1可知,各样本OTU覆盖度均在0.975以上,说明测序结果能反映样品中的绝大部分信息。在4种处理中,F1、F2和FM处理下的OUT均高于CK,F1处理组显著高于CK,可见F1处理下细菌的物种总数较多。α多样性指数是反映样本内微生物群落丰富度和均匀度的综合指标,其中ACE指数和Chaol指数能够反映物种的丰富度,指数越高说明物种数量越多;Shannon指数反映群落物种的多样性,该指数越大说明物种的多样性越高[17]。F1处理下3种α多样性指数均高于其他处理,其中Chaol指数与CK差异显著。F2处理下ACE指数、Chaol指数、Shannon指数均显著低于F1处理,其中Shannon指数显著低于CK。FM处理下细菌群落ACE指数、Chaol指数、Shannon指数较F1处理均显著降低,其中ACE指数和Chaol指数分别为F1处理的89.73%和89.27%。可见防风根际土壤(F1、F2和FM)细菌的OTU数均高于非根际土壤(CK),在防风根际土壤各处里组中,2年生(F2)和覆膜(FM)较1年生(F1)细菌多样性指数降低。
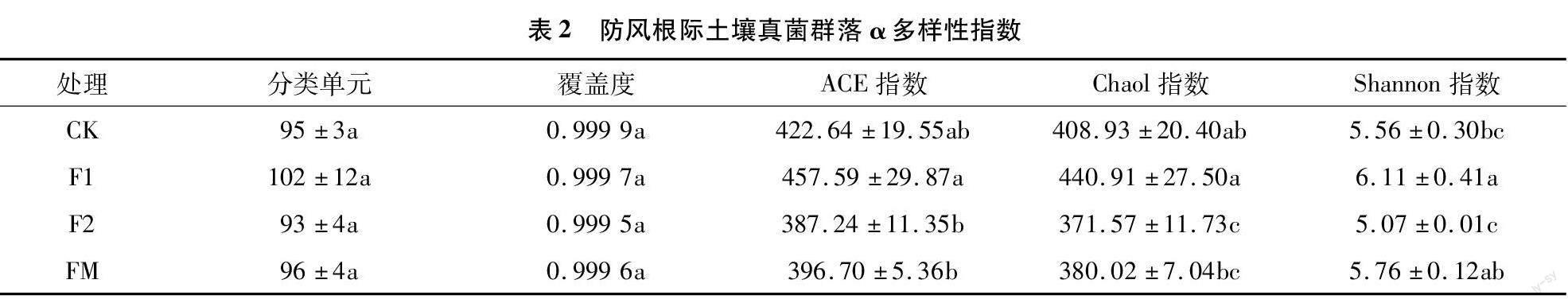
对测序后防风根际土壤真菌进行聚类OTU分析,结果(表2)表明,各样本的覆盖度均在0.999以上,各样本组间OTU数差异不显著。F1处理组真菌群落的ACE指数、Chaol指数、Shannon指數均高于F2处理、FM处理和CK组,其与F2处理组和CK组在Shannon指数中差异显著,其与F2和FM处理组在Chaol指数和ACE指数中差异显著。可见,防风根际土壤各处理组(F1、F2和FM)中真菌较非根际土壤(CK)真菌在物种种类上丰富度更高,防风根际土壤各处理组中1年生(F1)较2年生(F2)组和覆膜(FM)组真菌多样性高。
2.2 防风根际土壤微生物群落组成分析
测序后在非根际土壤中共获得23门120纲220目705科2 883属的细菌,在防风根际土壤中共获得25门122纲248目898科3 063属的细菌。从细菌门水平上分析,WPS-2门和FBP门仅在防风根际土壤中检测到。由图1-A可知,总体来看在所有样本中放线菌门(Actinobacteria)丰度最高(36.91%~50.36%),其次是变形菌门(Proteobacteria)(22.95%~39.11%)、酸杆菌门(Acidobacteria)(4.53%~12.25%)、绿弯菌门(Chloroflexi)(5.69%~10.01%)、厚壁菌门(Firmicutes)(4.07%~5.72%)和拟杆菌门(Bacteroidetes)(2.79%~5.10%)。这6个优势菌门所占相对比例均在5%以上,约占细菌总丰度的90%左右。在不同样本中各个菌门的占比不同,变形菌门在CK组的相对丰度为27.8%,在F1和FM处理中占比分别上升至29.94%和34.18%,在F2处理中的占比(38.28%)超过放线菌门的占比(28.13%),变形菌门成为F2处理组中丰度最高的优势菌群。F2和FM处理较F1处理酸杆菌门和拟杆菌门的占比增加,放线菌门和绿弯菌门的占比降低。从细菌科水平上分析(图1-B),防风根际土壤细菌主要由Subgroup-6、伯克氏菌科(Burkholderiaceae)、微球菌科(Micrococcaceae)、微单孢菌科(Micromonosporaceae)、念珠藻科(Nocardioidaceae)和鞘脂单胞菌科(Sphingomonadaceae)等组成。其中Subgroup-6在F2处理中丰度最高,为6.63%,分别是F1和FM处理的2.29倍和1.66倍。F2和FM处理较F1处理伯克氏菌科、鞘脂单胞菌科、芽单胞菌科(Gemmatimonadaceae)和亚硝化单胞菌科 (Nitrosomonadaceae)的占比增加,微球菌科、微单孢菌科、念珠藻科、地嗜皮菌科(Geodermatophilaceae)和红色杆菌科(Rubrobacteraceae)的细菌丰度则有所下降。FM处理较F1处理黄杆菌科(Flavobacteriaceae)和链霉菌科(Streptomycetaceae)细菌丰度分别增加了5.98倍和5.4倍。可见覆膜能够使原本丰度极低的一些细菌占比迅速增加。
4种处理的土壤样本中共检测到真菌12门17纲19目36科43属116种,其中芽枝霉门(Blastocladiomycota)仅在防风根际土壤中被检测到。由图2-A可知,从真菌门分类水平上看,平均相对丰度较高的菌门为子囊菌门(Ascomycota)(67.66%)、担子菌门(Basidiomycota)(10.09%)和被孢霉门(Mortierellomycota)(2.53%),其他门的丰度均在1%以下。子囊菌门在FM处理中的丰度最高,为78.25%,其次是F1和CK处理,在F2处理中的丰度最低,为49.12%,较FM处理的占比下降了37.22%。担子菌门在CK中的丰度最高,分别是F1、F2和FM处理的3.13倍、3.76倍和2.76倍。被孢霉门真菌在F2处理中较F1处理占比更高,在FM处理中较F1处理增加了2.53倍。由图2-B可知,从真菌科分类水平上看,小囊菌科(Microascaceae)、丛赤壳科(Nectriaceae)、假球壳科(Pleosporaceae)、被孢霉科(Mortierellaceae)、小双腔菌科(Didymellaceae)和暗壳腔菌科(Phaeosphaeriaceae)为优势真菌科,各科在不同处理中的丰度不同。小囊菌科在各处理中的占比由大到小依次为CK、F1、F2和FM,各根际土壤处理组分别为CK的73.11%、69.4%和40.15%。赤壳菌科在F2和FM处理中较F1处理的丰度降低。鬼伞菌科(Psathyrellaceae)在CK中丰度为13.59%,在FM和F1处理中丰度下降,FM处理中的丰度(3.77%)是F1处理中丰度(0.04%)的94倍。假球壳菌科在FM处理中丰度较高,分别是F1、F2处理的6.05、9.85倍。
2.3 防风根际土壤微生物群落结构相关性和差异性分析
去除各处理组中未明确分类(unclassified)的微生物群落OTU后,以科一级水平细菌和种一级真菌的OTU制作韦恩图。由图3-A可知,不同处理土壤中共有的细菌OTU数为405个,分别占CK、F1、F2和FM处理的59.47%、61.64%、69.83%和63.88%。1年生和对照组共有510个,2年生和对照组共有469个,覆膜组和对照组共有503个,1年生和覆膜组共有506个。各处理独有OTU数由高到低依次为:对照组(101个)、1年生(76个)、覆膜组(58个)、2年生(38个)。可见,共有OTU数在CK、F1、F2、FM处理中占59.47%~70.19%,每2组处理的共有OTU数在469~510个,对照组独有的OTU数最多。2年生独有OTU数最少,仅分别为F1和FM处理的50.00%和65.51%。
由圖3-B可知,不同处理土壤中共有的真菌OTU数为85个,分别占CK、F1、F2和FM的44.04%、45.70%、45.45%和47.49%。与土壤中独有的细菌OTU数在F2处理中最低相反,F2处理中独有的真菌OTU数在4个处理中最高,为28个,其次为CK、F1和FM处理。整体看来,F2与F1相比,无论在细菌还是真菌的总OTU数上都逐渐减小,说明土壤中微生物群落的整体多样性在不断减少。F2中独有的细菌OTU数远小于F1,而独有真菌的OTU数则高于F1,从侧面说明了土壤有从“细菌型”向“真菌型”转变的趋势。
为探明土壤微生物群落结构的差异性,对不同处理土壤样品的细菌和真菌群落组成相似度进行UPGMA聚类分析。由图4-A可以看出,4个处理的土壤细菌群落大致可分成3类:非根际土壤CK归为一类,与其他3个防风根际土壤的细菌群落组成相似度最低;F2和FM归为一类,表明2年生和覆膜栽培防风根际土壤中细菌群落组成相似度最高;1年生防风则介于前两类之间。由图4-B可以看出,土壤真菌群落的组成也可大致分为3类:CK和F1归为一类,两者的组成相似度最高;F2和FM各归为一类,其中FM的真菌群落结构相似度距离与其他3个处理的最远,表明覆膜栽培后其土壤中真菌群落结构经历了比较剧烈的变化。
2.4 防风根际土壤理化因子分析
通过对各处理土壤的理化因子分析发现(表3),除总有机碳(TOC)、含水量(WC)和pH值外,作为对照的非根际土壤各项理化因子均与1年生和2年生防风有显著差异。除全氮(TN)外,CK土壤中的全磷(TP)、全钾(TK)、硝态氮(N)和铵态氮(A)的含量均高于F1和F2,随着生长年限的增加防风根际土壤中的TK和A含量显著减少,TP和N含量无显著变化。虽然覆膜组较1年生组对土壤的TOC和TN无显著影响,但其能够在一定程度上增加土壤的TP、TK、N和WC,其中对TP和WC的影响达显著水平。此外覆膜后土壤的pH值降低,与非根际土壤组、1年生组和2年生组处理差异显著。土壤TOC含量在各处理间的差异均不显著,TN含量虽然在防风根际土壤中无显著差异,但其在F1、F2、FM处理中的含量却远高于CK,分别为CK的1.71、1.76、2.03倍。
2.5 土壤微生物群落种类与理化因子的相关性分析
为进一步探明不同处理土壤微生物群落种类与其理化因子之间的关系,对土壤中相对丰度前15的细菌和全部真菌(门水平)与土壤理化因子进行Spearman 相关性分析。如图5-A所示,在土壤中为优势菌群的几个细菌门中除变形菌门(Proteobacteria)的丰度与TK、A和N达到极显著负相关或显著负相关外,其他几个菌门如放线菌门(Actinobacteria)、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)和拟杆菌门(Bacteroidetes)细菌丰度受土壤理化因子影响不显著。与变形菌门相反,厚壁菌门(Firmicutes)细菌的丰度与N和A呈极显著正相关关系,与TP和TK呈显著正相关关系。此外,硝化螺菌门(Nitrospirae)、蓝细菌(Cyanobacteria)和疣微菌门(Verrucomicrobia)细菌的丰度分别与A、TOC和TK呈显著正相关关系;髌骨门(Patescibacteria)细菌的丰度与TK呈显著负相关关系。整体来看,对细菌丰度影响最大的理化因子是TK,其次是A和N,而TN、WC和pH等对各门细菌的丰度影响不明显。
由图5-B可知,土壤中优势菌群子囊菌门(Ascomycota)真菌的丰度与pH值呈显著负相关关系,担子菌门(Basidiomycota)真菌的丰度和TN呈极显著负相关关系,而被孢霉门(Mortierellomycota)真菌的丰度受土壤理化因子的影响不显著。各土壤理化因子中,TP、TK、N和A对一些门类的真菌影响较大,一些未分类的真菌门类(unclassified Fungi)和球囊菌门(Glomeromycota)真菌的丰度均与其呈显著负相关或极显著负相关关系;而N和A则与捕虫霉门(Zoopagomycota)真菌的丰度呈显著正相关关系。在14类真菌中,TOC和TN各对其中一类真菌产生了显著或极显著的负面影响;TP对其中两类产生了显著的负面影响;TK对其中3类产生了显著或极显著的负面影响;N、A和pH对其中至少3类产生了显著的正面或负面影响;WC对土壤真菌的丰度影响不明显。
2.5 微生物群落功能分析
本研究利用PICRUSt2对所有样本基于KEGG数据库进行细菌功能预测,结果显示其代谢通路可归为6大类,包括代谢(metabolism)、遗传信息处理(genetic information processing)、环境信息处理(environmental information processing)、细胞进程(cellular processes)、生物体系统(organismal systems)和人类疾病(human diseases),其中富集到代谢过程的数目最多,由高到低依次为异生素生物降解和代谢(xenobiotics biodegradation and metabolism)、碳水化合物代谢(carbohydrate metabolism)、萜类化合物和聚酮化合物的代谢(metabolism of terpenoids and polyketides)、氨基酸代谢(amino acid metabolism)、脂质代谢(lipid metabolism)、辅因子和维生素的代谢(metabolism of cofactors and vitamins)、其他次生代谢物的生物合成(biosynthesis of other secondary metabolites)。
基于MetaCyc数据库进行真菌功能预测,结果显示其功能可以划分为生物合成(biosynthesis)、退化/利用/同化(degradation/utilization/assimilation)、前体代谢物和能量的产生(generation of precursor metabolite and energy)、聚糖途径(glycan pathways)和代谢簇(metabolic clusters)五大类,其中聚集在生物合成过程的基因数目最多,特别在辅因子、辅基、电子载体和维生素生物合成(cofactor,prosthetic group,electron carrier,and vitamin biosynthesis),脂肪酸和脂质生物合成(fatty acid and lipid biosynthesis),核苷和核苷酸生物合成(nucleoside and nucleotide biosynthesis),碳水化合物降解(carbohydrate degradation),氨基酸生物合成(amino acid biosynthesis),碳水化合物生物合成(carbohydrate biosynthesis),氨基酸降解(amino acid degradation),次生代谢物生物合成(secondary metabolite biosynthesis)中。
如图6所示,对各样本的细菌和真菌功能分别进行PCoA分析,当2点在坐标轴上的投影距离越近,表明这2个样本在相应维度中的功能组成越相似。结果显示77.6%的PCoA轴1和2组合解释了功能类别变化,除F2处理组外其他组的投影距离较为接近,由此可见,防风根际土壤微生物的功能差异受种植年限的影响较为明显。
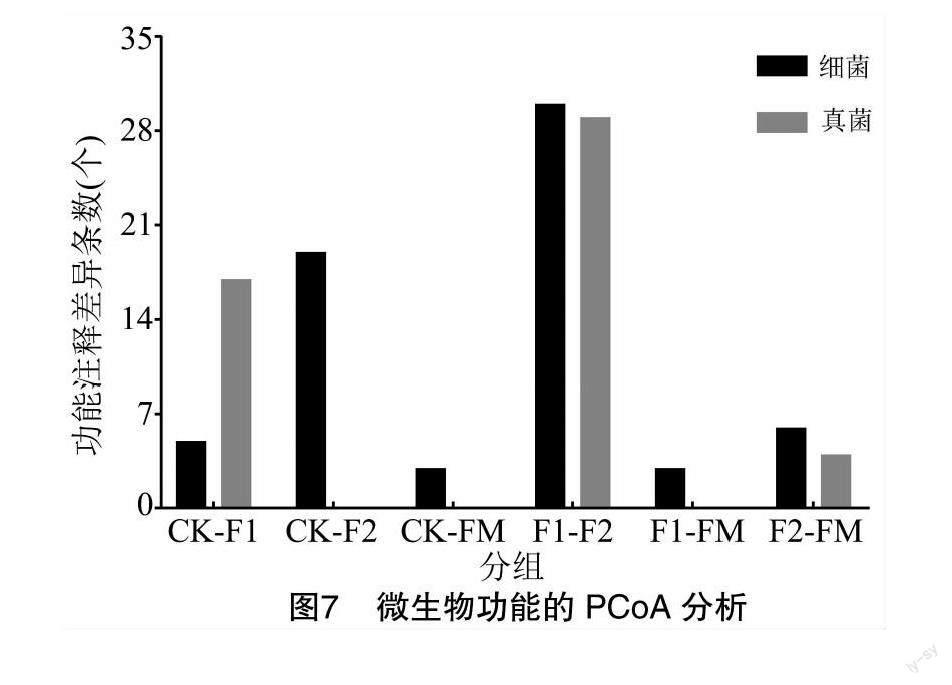
在不同生长年限和不同栽培模式下的防风,尽管其各样品所含根际微生物代谢通路有相似性,但不同样本所囊括的代谢通路的丰度值不同。如图7所示,对不同样本有显著差异(P<0.05)的通路进行统计分析发现,在细菌功能注释中各样本差异通路由大到小依次为F1与F2(30条)、CK与F2(19条)、F2与FM(6条)、CK与F1(5条)、F1与FM(3条)、CK与FM(3条);与其类似,各真菌功能注释有显著差异(P<0.05)的主要表现在F1与F2、CK与F1和F2与FM间,分别有29条、17条和4条。可见,F1与F2间所囊括的差异性功能注释条目数最多,其次是CK与不同年份(F1、F2)间和F2与FM间。可见相较于覆膜处理,生长年限对微生物功能的影响更加明显,特别是对于生长2年的防风其根际土壤微生物功能变化最为显著。
3 讨论与结论
大量研究表明,植物-根际微生物-环境因子之间存在着彼此依赖、往复调控和相互制约的复杂关系。植物根际有着丰富的微生物,这些微生物可以直接或间接的影响药用植物的生长发育和品质形成[1]。根际微生物群落是植物性能的关键调节因子,根际微生物群落是由农业管理和宿主选择过程相互作用决定的[17]。因此,研究植物根际土壤微生物多样性、群落结构特征以及与周围环境因子之间的相互关系,有利于解析植物生长发育调控机制,尤其对药用植物栽培生产具有十分重要的意义。土壤微生物的多样性水平一直被认为是评价土壤生态功能的重要指標。武睿等研究发现,随着生长年限的增加,甘肃贝母根际土壤中的细菌群落多样性会显著降低,5年生贝母细菌群落的OTU分类单元、Chaol指数和Shannon指数均低于对照和1年生贝母[18]。不同种植年限牡丹根际土壤中真菌群落多样性之间有显著差异,随着生长年份的增加,真菌多样性总体上呈先上升后降低的趋势,特别是在种植10年的土壤中真菌的ACE指数和Sobs指数降至最低[19]。在对高温覆膜栽培的韭菜根际土壤研究时发现,覆膜前后细菌和真菌的ACE指数、Chaol指数和Shannon指数等并无显著差异[20]。程茂高等通过稀释培养计数法对防风根际土壤研究发现,随着防风生长期的延长,土壤中微生物总量逐渐下降,其中1年生和2年生防风根际土壤的细菌和真菌总量分别是对照的91.78%、55.31%和79.45%、67.12%[21]。与其研究结果不同,本研究发现,1年生防风根际土壤中无论是细菌还是真菌的α多样性指数均高于作为对照的非根际土壤,说明栽培防风后土壤中微生物的多样性在一定时期内是增加的;F2和FM处理的ACE等指数低于CK和F1处理,说明随着种植时间由1年变为2年和覆膜栽培的实施,根际土壤中微生物的多样性降低。总体来看,微生物的总数、菌群丰富度和复杂程度在测定样本中的大小为:1年生组>非根际土壤组;1年生组>2年生组、覆膜组。这与前人在华重楼[22]、太白贝母[23]、三七[24]和枸杞[25]等药用植物中的研究一致。
植物的种类和生长发育阶段直接影响着土壤微生物的群落组成结构。大多数植物,如大豆、玉米、花生、豇豆的根际土壤中,细菌以变形菌门和放线菌门为优势菌群[26]。而在种植辣椒的土壤中以变形菌门和酸杆菌门为优势细菌[27],变形菌门、厚壁菌门和酸杆菌门则是香蕉园土中的主要细菌类群[28]。本研究发现,防风土壤样品中细菌丰度最大的2个门始终是放线菌门和变形菌门,真菌丰度最大的也一直是子囊菌门和担子菌门,由此不难看出,这些门类的细菌和真菌不但在土壤中广布存在,还能极好地适应各种植物的根际环境。不同处理下防风的土壤微生物群落结构具有一定差异,2年生组和覆膜组较1年生组中细菌的变形菌门和酸杆菌门占比增加,放线菌门细菌丰度占比减小。程茂高等在其研究中发现,2年生防风根际放线菌门细菌的丰度是不断增高的,推测可能是因防风品种及种植地条件不同所致[21]。胡志娥等对长期覆膜条件下的农田土壤微生物群落研究發现,长期覆膜使土壤真菌共生网络变得简单而脆弱,其关键物种仅有粪壳菌目(Sordariales)中的未知菌属一种,而这种菌种的单一性会对农田土壤生态环境带来潜在风险[29]。从门水平上看,覆膜栽培增加了防风根际土壤中子囊菌门和担子菌门真菌的占比,但对二者的影响并不显著。从科一级水平看,覆膜不但显著的降低了在物质和能量循环中起重要作用的小囊菌科真菌丰度,而且极大地增加了假球壳科等植物病原真菌群落占比。很多药用植物在长期种植后,根际土壤细菌的多样性会逐渐降低,土壤微生物区系也会由高肥力的“细菌型”向低肥力的“真菌型”过渡[23,30-32]。通过聚类分析发现,2年生和覆膜栽培防风土壤样品,无论是细菌还是真菌在群落的组成上都比对照和1年生防风更为接近。结合土壤微生物群落多样性在F2中的整体降低以及F2中独有真菌种类的增多不难看出,防风生长年限由1年增长到2年时其根际土壤微生物区系有从“细菌型”向“真菌型”转变的趋势。
除植物种类等生物因素外,根际土壤微生物群落结构还受气候和土壤理化因子如pH值、含水量、有机碳、全氮等多方面影响。在甘肃贝母根际土壤中,除TN、TK和TP外,pH值、有机质、水解氨、速效磷和速效钾是驱动其根际土壤细菌群落的主要因子[18]。野生北苍术土壤的pH值、有机质、有效钾是AM真菌多样性和丰度的主要影响因子[33]。柴胡、党参和远志根际土壤的TN、总碳、pH值、脲酶、过氧化氢酶等分别影响了硝化螺旋菌门、绿弯菌门、浮霉菌门、芽单胞菌门细菌类群的丰度[34]。在以上研究中,pH值是影响土壤微生物群落结构的重要因子,通常情况下pH值与土壤中酸杆菌的相对丰度呈显著负相关关系[35],但在本研究中,pH值对酸杆菌门细菌的丰度无显著影响,甚至所有检测到的细菌门类都不受pH值影响,而酸杆菌门细菌与所检测的8种土壤理化因子之间也无明显相关性。pH值对防风根际土壤细菌门类的影响不显著,但子囊菌门、一些未分类门和捕虫霉门(Zoopagomycota)真菌的丰度却与其显著相关。李毳等研究发现,土壤环境因子对根际微生物群落结构和多样性具有显著影响,但植物种类本身对微生物群落结构的影响更为重要[34,36]。也有研究发现,变形菌门细菌更偏好于在高养分的土壤环境中生存,其菌群丰度与土壤有机质和全氮含量显著相关[37]。在本研究中,变形菌门细菌的丰度受TOC和TN影响并不显著,而是和TK、N和A显著相关。根据相关性分析结果,TK、N和A不仅是防风根际土壤细菌群落的主要驱动因子,也是影响真菌组成的主要因子,但特别的是,本研究中几个优势的细菌门类和真菌门类(除未分类门)均与TK、N和A无显著相关性。土壤微生物通过参与土壤有机质分解和矿化等过程来影响土壤养分循环,并调节土壤的功能[38]。利用PICRUSt方法基于KEGG数据库和MetaCyc数据库分别对细菌和真菌菌群代谢功能预测,发现细菌功能主要集中在异生素生物降解代谢和碳水化合物代谢,真菌功能主要集中在生物合成。在甘蔗和水稻上研究发现此类功能对植物生长、生存具有积极作用[39-40]。PCoA分析表明微生物功能差异主要表现在2年生组。对差异性功能注释条目进行统计也表明,2年生组较1年生组显著差异条目数最多。再结合各样本OTU数和Chao1指数对比来看,2年生组和覆膜组根际微生物较1年生丰富度减小、微生物功能减弱。综合来看,土壤理化因子和各处理均对防风根际土壤微生物群落结构造成影响,各个处理对微生物群落结构组成影响更大,特别是生长年份的影响作用尤为明显,覆膜影响次之。
本研究初步揭示了不同生长年限和覆膜栽培防风根际土壤中微生物群落的变化规律,发现生长年份是影响防风根际微生物群落结构组成的重要因子。研究结果提示我们在防风的田间管理中,尤其是在栽培的第2年应适量增施农家肥和生物肥,以保证其土壤微生物的多样性。在今后的研究中我们还将继续开展其余年份和其他类型的覆膜实验,为防风合理栽培措施的选择提供科学依据。
参考文献:
[1]祝 蕾,严 辉,刘 培,等. 药用植物根际微生物对其品质形成的影响及其作用机制的研究进展[J]. 中草药,2021,52(13):4064-4073.
[2]江 曙,段金廒,钱大玮,等. 根际微生物对药材道地性的影响[J]. 土壤,2009,41(3):344-349.
[3]肖艳红,李 菁,刘祝祥,等. 药用植物根际微生物研究进展[J]. 中草药,2013,44(4):497-504.
[4]Berendsen R L,Pieterse C M J,Bakker P A H M.The rhizosphere microbiome and plant health[J]. Trends in Plant Science,2012,17(8):478-486.
[5]Nihorimbere V,Ongena M,Smargiassi M,et al. Beneficial effect of the rhizosphere microbial community for plant growth and health[J]. Biotechnology,Agronomy,Society Environnement,2011,15(2):327-337.
[6]Akyol ,Ince O,Ince B. Crop-based composting of lignocellulosic digestates:focus on bacterial and fungal diversity[J]. Bioresource Technology,2019,288:121549.
[7]Lundberg D S,Lebeis S L,Paredes S H,et al. Defining the core Arabidopsis thaliana root microbiome[J]. Nature,2012,488(7409):86-90.
[8]Schreiter S,Ding G C,Heuer H,et al. Effect of the soil type on the microbiome in the rhizosphere of field-grown lettuce[J]. Frontiers in Microbiology,2014,5:144.
[9]李欣玫,左易靈,薛子可,等. 不同荒漠植物根际土壤微生物群落结构特征[J]. 生态学报,2018,38(8):2855-2863.
[10]刘 思,徐国前,张军翔. 葡萄行内覆盖对土壤细菌群落结构的影响[J]. 核农学报,2020,34(12):2865-2871.
[11]郑少奇,任北大,张保春. 关于张元素应用防风经验之探析[J]. 中医文献杂志,2018,36(2):16-18.
[12]冯学锋,付桂芳,格小光,等. 中药防风栽培品与野生品药材性状显微组织差异比较研究[J]. 中国中药杂志,2009,34(22):2862-2866.
[13]贾俊英,曹 瑞,张小波,等. 基于纹理和光谱信息的奈曼旗防风遥感解译[J]. 中国中药杂志,2019,44(19):4111-4115.
[14]王思淼,霍金海,孙国东,等. 基于UPLC-Q-TOF/MS技术的不同生长年限防风中香豆素成分分析[J]. 中国中医药科技,2019,26(3):362-367,483.
[15]王英范,刘双利,刘桓宇,等. 一、二年生栽培防风有效成分比较研究[J]. 吉林农业大学学报,2006,28(3):289-291.
[16]鲍士旦. 土壤农化分析[M]. 3版. 北京:中国农业出版社,2000.
[17]Schmidt J E,Kent A D,Brisson V L,et al. Agricultural management and plant selection interactively affect rhizosphere microbial community structure and nitrogen cycling[J]. Microbiome,2019,7(1):146.
[18]武 睿,陈 垣,郭凤霞,等. 生长年限对甘肃贝母根际土壤细菌群落结构的影响[J]. 干旱地区农业研究,2021,39(6):153-161.
[19]耿晓东,周 英,汪成忠,等. 不同种植年限对凤丹牡丹根际真菌群落组成和多样性的影响[J]. 江苏农业科学,2021,49(23):145-151.
[20]宋 健,张海剑,刘 莉,等. 高通量测序分析高温覆膜对韭菜根际微生物多样性的影响[J]. 中国生物防治学报,2020,36(6):938-945.
[21]程茂高,乔卿梅,王新民,等. 防风根际微生物区系和土壤酶活性的变化研究[J]. 广东农业科学,2009,36(12):81-83.
[22]冼康华,苏 江,付传明,等. 不同生长年限华重楼根际土壤微生物多样性研究[J]. 广西植物,2022,42(12):2087-2098.
[23]母茂君,张弟桂,张 华,等. 太白贝母根际微生物分布与生物碱含量的相关性[J]. 中国中药杂志,2019,44(11):2231-2235.
[24]Dong L,Xu J,Feng G,et al. Soil bacterial and fungal community dynamics in relation to Panax notoginseng death rate in a continuous cropping system[J]. Scientific Reports,2016,6:31802.
[25]肖龙敏,唐 明,张好强. 不同种植年限宁夏枸杞根际微生物的群落多样性[J]. 西北林学院学报,2018,33(6):31-39,163.
[26]唐 杰,陈知青,郭安南,等. 不同作物根际土壤微生物的群落结构特征分析[J]. 核农学报,2021,35(12):2830-2840.
[27]康林玉,刘周斌,欧立军,等. 辣椒种植对根际土壤微生物多样性的影响[J]. 湖南农业大学学报(自然科学版),2018,44(2):151-156,175.
[28]黄 珍,谭志琼,阮云泽. 香蕉园土壤16S rDNA文库分析[J]. 热带作物学报,2010,31(6):989-993.
[29]胡志娥,肖谋良,丁济娜,等. 长期覆膜条件下农田土壤微生物群落的响应特征[J]. 环境科学,2022,43(10):4745-4754.
[30]周 浓,戚文华,肖国生,等. 滇重楼根际微生物分布与甾体皂苷含量的相关性[J]. 中国中药杂志,2015,40(6):1055-1060.
[31]孙世中,官会林,张云峰,等. 设施栽培下香石竹不同植株状态根际土壤微生物类群变化分析[J]. 土壤,2011,43(1):72-75.
[32]廖海兵,李云霞,邵晶晶,等. 連作对浙贝母生长及土壤性质的影响[J]. 生态学杂志,2011,30(10):2203-2208.
[33]李铭杰,周志杰,邢礼军,等. 北苍术根区土壤中AM真菌多样性及其与土壤养分相关性分析[J]. 中国生物防治学报,2021,37(6):1288-1297.
[34]李 毳,刘 怡,刘晋仙. 药用植物根际细菌群落多样性驱动因素分析[J]. 生态环境学报,2020,29(10):1988-1993.
[35]Griffiths R I,Thomson B C,James P,et al. The bacterial biogeography of British soils[J]. Environmental Microbiology,2011,13(6):1642-1654.
[36]李 毳,刘 怡. 晋东南3种道地药材植物根际真菌群落特性[J]. 生态环境学报,2019,28(7):1388-1393.
[37]Liu J J,Sui Y Y,Yu Z H,et al. Soil carbon content drives the biogeographical distribution of fungal communities in the black soil zone of northeast China[J]. Soil Biology and Biochemistry,2015,83:29-39.
[38]Wang J H,Liu L,Gao X Y,et al. Elucidating the effect of biofertilizers on bacterial diversity in maize rhizosphere soil[J]. PLoS One,2021,16(4):e0249834.
[39]Huang W J,Lu Y L,Chen L J,et al. Impact of pesticide/fertilizer mixtures on the rhizosphere microbial community of field-grown sugarcane[J]. 3 Biotech,2021,11(5):210.
[40]许忠伟,刘春月,石佳悦,等. 稻蟹共作田不同生育期水稻根际土壤细菌群落结构和功能分析[J]. 中国稻米,2022,28(4):61-66.
猜你喜欢
发明与创新(2023年36期)2023-11-29 12:57:16
基层中医药(2021年3期)2021-11-22 08:07:58
小哥白尼(趣味科学)(2021年11期)2021-02-28 08:34:58
戏剧之家(2016年22期)2016-11-30 15:13:39
散文百家·下旬刊(2016年9期)2016-11-23 22:23:14
散文百家·下旬刊(2016年10期)2016-11-17 21:22:22
科技资讯(2016年19期)2016-11-15 10:39:12
人间(2016年27期)2016-11-11 16:27:23
农业与技术(2016年15期)2016-11-09 07:11:20
云南科技管理(2015年5期)2015-12-22 10:34:38
