
miR-320a-5p靶向Twist抑制肝细胞癌上皮间质转化
2023-08-07 07:18张慧明楚振升吕丹刘爽
中国老年学杂志 2023年15期
张慧明 楚振升 吕丹 刘爽
(佳木斯大学 1基础医学院,黑龙江 佳木斯 154002;2体育学院)
肝癌是全球第五大致命癌症〔1〕,死亡率高。肝癌可分为原发性和继发性两大类,原发性肝癌起源于肝脏的上皮或间叶组织;而继发性肝癌较为少见〔2〕。其中,肝细胞癌(HCC)是最常见的原发性肝癌,占原发性肝癌患者的70%~90%〔3〕。在中国,每年新增或死亡的HCC患者约占病例总数和死亡总数的50%〔4〕。目前肝癌的治疗方法有外科手术及药物治疗,但由于癌细胞不受控制的增殖和转移导致癌症迅速发展;且药物通常存在一系列副作用,极易产生肿瘤耐药性,在临床应用上受到极大的限制,目前肝癌患者的长期生存率仍然很低。其中,对于HCC,肺转移是HCC患者生存率低的主要原因之一〔5〕。因此,迫切需要阐明HCC发生发展的相关机制,确定新的治疗靶点以改善临床治疗疗效。
microRNA(miRNA)是重要的非编码RNA,通过结合mRNA的3′-非翻译区(UTR)调节蛋白质编码基因,发挥信号传递的生物学功能〔6〕。miRNA在肿瘤生物学中的重要性已得到广泛认可,且由于其特异的分子病理学特征,miRNA表达谱已成功用于分类不同肿瘤或肿瘤发展的不同阶段〔7〕。有研究指出,miR-320a-3p过表达可以通过下调信号转导和转录激活因子(STAT)3表达来抑制肺纤维化中的上皮间质转化(EMT)过程〔8〕。miR-320a可通过沉默糖皮质激素调节元件结合蛋白(GMEB)1的表达促进EMT表型改变,抑制结直肠癌细胞侵袭〔9〕。可见,miR-320a能够通过影响肿瘤细胞的生物学活动而影响肿瘤的发展,但是目前的研究暂未揭示miR-320a-5p对HCC的转移及转移的影响。因此,本研究通过开展体外试验探究miR-320a-5p对HCC的影响及其机制,以期为HCC的临床治疗提供新的治疗靶点和治疗策略。
1 材料与方法
1.1临床样本采集 HCC患者肿瘤组织(HCC组)和相邻组织学正常的组织(对照组)购于上海芯超公司。所有研究程序经佳木斯大学研究伦理委员会批准。
1.2细胞培养与处理 人正常肝细胞lo2、肝癌细胞系HepG2、Hcclm3细胞均购自中科院干细胞库。将细胞培养在含有10%胎牛血清(以色列,Bi),1%青-链霉素(博士德有限公司)的完全RPMI1640(美国,Gibco)中,37 ℃和5%CO2培养箱(美国,西蒙公司)培养。按照lipo3000(美国,Thermo)转染步骤,对HepG2和Hcclm3细胞转染阴性mimics、miR-320a-5p mimics、阴性pcDNA3.1、pcDNA3.1-Twist,将细胞分为NC mimics组、miR-320a-5p组、NC mimics+NC组、miR-320a-5p+NC组、NC mimics+Twist组、miR-320a-5p+Twist组。
1.3qRT-聚合酶链反应(PCR)检测 采用TRizol法提取癌组织、各处理组细胞总RNA,NanoDrop检测RNA的浓度及纯度。按照随机引物逆转录试剂盒制备cDNA(美国,Thermo)。按照SYBR GREEN试剂盒(日本,TaKaRa)说明对miR-320a-5p和Twist mRNA的表达水平进行检测。以U6和磷酸甘油醛脱氢酶(GAPDH)作为内参,实验设置6个重复。采用2-ΔΔCt法对数据进行定量分析,计算目的基因的相对表达量。引物序列:miR-320a-5p正向引物:5′-AAAAGCUGGGUUGAGAGGGCGA-3′,反向:5′-GCCCUCUCAACCCAGCUUUUUU-3′;Twist正向引物:5′-GTCTCCGTCGCTTTCAGACGAT-3′,反向:5′-ACA-CGGAGAAGGCGTAGCTGAG;U6正向引物:5′-CT-CGCTTCGGCAGCACA,反向:5′-AACGCTTCACGA-ATTTGCGT;GAPDH正向引物:5′-TCCCATCACCAT CTTCCA,反向:5′-CATCACGCCACAGTTTTCC。
1.4CCK8法检测 将处理后对数生长期细胞接种于96孔板中103个/孔,分别培养24、48、72 h后每孔加入CCK8溶液(1∶9),于培养箱内继续培养1 h。用酶标仪测定波长570 nm处吸光度(OD)值。
1.5细胞集落形成实验 使用0.25%胰酶消化细胞,用含0.35%琼脂糖的RPMI1640完全培养基重悬细胞,以1×105个/孔的密度接种于含0.6%琼脂糖的6孔板中,置于37 ℃ 5%CO2培养箱中培养。当肉眼可见集落形成,终止培养,0.1%结晶紫溶液染色,用倒置显微镜拍照并统计集落数量。
1.6Transwell实验检测 基质胶包被Transwell小室上室,并置于 37 ℃、5% CO2培养箱中静置约30 min。在Transwell小室上室加入100 μl待测细胞悬液,在小室下室中加入700 μl含20%胎牛血清的培养基。37 ℃,5% CO2培养12~24 h,用磷酸盐缓冲液(PBS)清洗小室3次,用1%戊二醛固定小室30 min,擦去基质胶及未侵袭的细胞,0.1%结晶紫染色。用PBS洗净,干燥后正置显微镜观察,随机观察6~10个视野,记录每个视野内阳性细胞数并进行统计分析。
1.7划痕实验检测 将细胞接种到六孔板中,分别进行转染处理。用10 μl无菌枪头垂直于横线划痕。用PBS洗涤去除悬浮细胞和细胞碎片。添加新鲜的无血清培养基,培养24 h后,用倒置显微镜拍照,并采用ImageJ软件计算划痕面积。
1.8双荧光素酶报告基因实验 通过生信网站预测miR-320a-5p与Twist的结合位点,将miR-320a-5p与Twist结合部位的序列及其突变体序列插入萤火虫荧光素酶基因下游构建表达载体(美国,Promega)。将miR-320a-5p mimics与pmirGLO-Twist-WT/MUT重组质粒,对照组mimics NC与Twist的野生型或突变型重组质粒,利用Lipo3000转染至293T细胞。转染48 h后,根据双荧光素酶报告基因检测试剂盒说明书检测荧光素酶活性。
1.9Western印迹检测 使用细胞裂解液提取细胞总蛋白,用二喹啉甲酸(BCA)试剂盒测定提取蛋白的浓度。取20 μg蛋白加1×上样缓冲液煮沸变性,通过十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)将蛋白分离,且将蛋白转移到聚偏氟乙烯(PVDF)膜上。5%脱脂奶粉封闭1 h后,加入一抗,4 ℃过夜孵育。次日洗膜3次,加入二抗,在温室中孵育1 h。再洗膜3次后,加入化学发光试剂显影蛋白,置于凝胶成像系统中采集图像,采用ImageJ软件分析蛋白条带的灰度水平。以GAPDH为内参,计算蛋白相对表达量。
1.10统计学分析 采用GRAPHPAD9.0软件进行方差分析、独立样本T检验和Pearson相关性分析。
2 结 果
2.1两组miR-320a-5p mRNA在HCC组织及细胞中表达比较 在细胞中,与对照组miR-320a-5p表达水平(1.00±0.00)比较,HCC组细胞系HepG2、Hcclm3中miR-320a-5p表达水平(分别为0.28±0.12、0.62±0.06)显著降低(均P<0.05),且在HepG2细胞中表达低,在Hcclm3中表达高。在组织中,与对照组(5.88±1.31)相比,HCC组miR-320a-5p mRNA表达水平显著降低(4.61±1.00,P<0.001)。由此可知,miR-320a-5p可能参与了HCC的进展。
2.2miR-320a-5p抑制HCC细胞增殖 与NC mimics组比较,miR-320a-5p组HepG2及Hcclm3细胞miR-320a-5p mRNA表达水平显著升高(均P<0.05)。CCK8和细胞集落实验结果显示,miR-320a-5p过表达后HCC细胞HepG2增殖能力在48 h显著降低(P<0.05),HepG2及Hcclm3细胞活力显著降低(P<0.01),见表1、表2、图1。

图1 细胞集落形成实验检测细胞活力(结晶紫染色)

表1 两组不同时间HCC细胞增殖能力
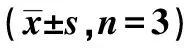
表2 两组HCC细胞活力、迁移率、侵袭个数、miR-320a-5p mRNA及Twist表达比较
2.3miR-320a-5p抑制HCC细胞的转移和EMT 在HepG2、Hcclm3细胞中,与NC mimics组比较,miR-320a-5p组细胞的侵袭、迁移能力显著降低(P<0.001,P<0.01,P<0.05)。见表2、图2。Western印迹结果显示,过表达miR-320a-5p后,HepG2和Hcclm3细胞中E-钙黏蛋白(cadherin)表达显著升高,波形蛋白(Vimentin)表达水平显著降低(P<0.001,P<0.01,P<0.05),见图3、表3。以上数据表明miR-320a-5p参与抑制HCC细胞的转移和EMT形成。

图2 miR-320a-5p抑制HCC细胞侵袭(结晶紫染色,×100)、迁移(×100)

图3 Western印迹检测各组细胞EMT相关蛋白表达
表3 Western印迹检测各组目的蛋白表达
2.4miR-320a-5p靶向Twist 通过生信网站预测可知,miR-320a-5p与Twist间存在互作序列(图4)。双荧光素酶实验发现HepG2、Hcclm3细胞中miR-320a-5p mimics可以显著降低Twist的荧光素酶活性(P<0.001),证实miR-320a-5p可以靶向结合Twist,见表4。与对照组(3.66±1.15)比较,HCC组中Twist mRNA表达显著升高(4.81±0.94,P<0.001)。在HepG2、Hcclm3细胞中,与NC mimics组比较,过表达miR-320a-5p后细胞中Twist表达显著升高(均P<0.05)。见表2。Pearson相关性分析证实miR-320a-5p和Twist在HCC中的表达呈负相关(r=-0.621 3,P=0.005 9)。见图5。上述结果表明,Twist是miR-320a-5p的靶向基因。

图4 miR-320a-5p与Twist的结合位点

图5 miR-320a-5p和Twist在肝癌细胞中表达呈负相关
表4 HCC细胞相对荧光素酶活性
2.5miR-320a-5p通过靶向Twist抑制HCC的进展 与NC mimics+NC组比较,NC mimics+Twist组细胞的增殖、侵袭、迁移能力显著升高(P<0.05,P<0.001)。与miR-320a-5p+NC组比较,miR-320a-5p+Twist组细胞增殖、侵袭、迁移能力显著升高(P<0.01,P<0.05)。与NC mimics+Twist组比较,miR-320a-5p+Twist组细胞侵袭、迁移及24 h增殖能力显著降低(P<0.001)。见表5、表6、图6。以上结果说明,miR-320a-5p过表达可逆转Twist对HCC细胞恶性生物学活动的促进作用。

图6 miR-320a-5p逆转Twist在HCC细胞的促肿瘤作用(结晶紫染色,×100)
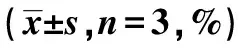
表5 4组细胞侵袭、迁移能力比较
表6 CCK8法检测不同处理组HepG2细胞增殖
3 讨 论
肝癌是最常见的侵袭性癌症之一,其中HCC是最常见的原发性肝癌。虽然目前手术切除、化学疗法和放射能够被用于治疗HCC患者,但由于HCC具有较强的侵袭和转移性质,患者在术后往往会出现复发、肿瘤转移或生存差的情况〔10〕。因此,迫切需要揭示HCC发生发展的分子机制,以开发新颖、有效的HCC治疗策略。
miRNA作为癌症中基因表达必不可少的表观遗传调控因子,通过与mRNA 3′-UTR结合导致mRNA降解或翻译受阻〔11〕,在癌症发生发展过程中发挥复杂多样的功能。以HCC为例,Lu等〔12〕发现,miR-124能够通过靶向转录激活因子STAT3抑制癌细胞增殖并诱导其凋亡,进而抑制HCC的生长;Hua等〔13〕发现,miR-142-3p能够靶向乳酸脱氢酶(LDHA)抑制糖酵解,从而抑制HCC细胞的增殖;Lin等〔14〕研究表明miR-122可以通过直接靶向内源性凋亡调节因子B细胞淋巴瘤(Bcl)-w的3′-UTR结合位点,降低Bcl-w的mRNA和蛋白水平,抑制癌细胞活力,活化含半胱氨酸的天冬氨酸水解酶(caspase)-3而促进凋亡。而正如前言所述,miR-320a-5p同样被确定为在许多癌症中能够抑制肿瘤〔15〕,发挥不同功能角色,但目前在HCC中的作用及相关机制尚不清楚。因此,本研究提示,miR-320a-5p与HCC的发展和转移有关。体外试验发现,过表达miR-320a-5p能够显著抑制HCC细胞的增殖能力、细胞活力、迁移能力、侵袭能力和EMT形成。其中,EMT对于肿瘤转移是必不可少的重要机制,具体表现为上皮细胞形状显著改变,获得运动及浸润能力及间充质表型。目前认为,EMT发生的主要标志是E-cadherin表达降低、N-cadherin和Vimentin的表达升高,引起肿瘤细胞的黏附分子结构功能异常,肿瘤细胞间的黏附能力下降,运动能力增加,肿瘤细胞更易发生浸润或转移。这与本实验研究结果一致。这些数据共同证实miR-320a-5p能够抑制HCC细胞增殖、侵袭、迁移及EMT形成,进而抑制HCC体内生长和转移。
Twist是twist家族的bHLH转录因子1,是EMT关键的诱导转录因子,通过上皮表型相关基因和上调间质细胞表型相关基因来调控EMT相关基因的转录〔16〕,参与促进细胞分化、增殖、转移,具有抗凋亡作用,并与癌症干细胞表型相关〔17〕。Grzegrzolka等〔18〕指出,Twist在乳腺癌高表达,激活EMT趋势,是原位乳腺癌发展为侵袭性形式的关键。而在HCC方面,Meng等〔19〕指出,Twist通过Circular RNA-Cul2上调Vimentin,促进HCC的EMT趋势,说明Twist是介导EMT进而促进肿瘤发生转移的关键转录因子。本研究发现,Twist在HCC中上调,提高HCC的EMT趋势。通过生物信息学预测分析和双荧光素酶实验结果得到,miR-320a-5p可靶向Twist蛋白,且二者表达水平呈负相关趋势。进一步探究发现,Twist会促进HCC的发展和转移,而miR-320a-5p能够逆转Twist对肿瘤的促进作用。由此可知,miR-320a-5p通过靶向Twist抑制HCC的转移和EMT。
致谢:黑龙江省北药与功能食品优势特色学科建设项目、佳木斯大学基础医学院微生态-免疫调节网络与相关疾病重点实验室提供资助!
猜你喜欢
保健医苑(2022年5期)2022-06-10
天津医科大学学报(2021年4期)2021-08-21
中国临床医学影像杂志(2021年6期)2021-08-14
中日友好医院学报(2021年1期)2021-04-14
肝博士(2020年5期)2021-01-18
山东医药(2020年9期)2020-05-20
天津医科大学学报(2019年3期)2019-08-13
肿瘤预防与治疗(2015年1期)2015-09-26
医学研究杂志(2015年7期)2015-06-22
中国当代医药(2015年16期)2015-03-01
