
渤海西岸多管藻类的形态分类学研究
2023-07-29 07:21闫盼竹丁兰平黄冰心刘美媛王雪聪秦佳男
海洋科学 2023年5期
闫盼竹, 楚 越, 丁兰平, 黄冰心, 刘美媛, 王雪聪, 秦佳男
渤海西岸多管藻类的形态分类学研究
闫盼竹, 楚 越, 丁兰平, 黄冰心, 刘美媛, 王雪聪, 秦佳男
(天津师范大学 生命科学学院 天津动植物抗性重点实验室, 天津 300387)
利用显微观察及生物冷冻切片技术, 对采集自河北秦皇岛的多管藻类(Complex)海藻样品进行形态分类学研究。经鉴定, 样品中包括: 多管藻属()2种: 多管藻(Harvey)和丛托多管藻(Harvey); 新管藻属()2种: 倾伏新管藻((Segi) Kim et Lee)和球果新管藻((Børgesen) Kim et Lee), 其中丛托多管藻、倾伏新管藻及球果新管藻均为渤海西岸首次记录。本文对样品物种的外部形态及内部结构特征进行了详细描述, 并提供了清晰的特征图片。研究结果补充了渤海湾内的藻类资源数据, 为多管藻类相关研究提供了分类学参考。
多管藻属(); 新管藻属(); 秦皇岛; 分类学; 红藻
松节藻科(Rhodomelaceae)由ARESCHOUG[1]于1847年建立, 隶属于红藻门(Rhodophyta)、仙菜目(Ceramiales), 是目前世界上红藻门海藻中属种最多的科[2], 国际上报道了超过1 000个物种[3], 中国报道了其中的26属139种[2, 4]。多管藻类(Complex)是该科中的一大类群, 在中国主要包括多管藻属()和新管藻属()。两属物种丰富, 藻体富含多种活性物质, 具有抗凝血、抗病毒、调节免疫、抗肿瘤等活性[5-7], 具有良好的研究价值、经济价值和药用价值。且因海水养殖及船舶运输等人类活动干扰的外在因素以及该类群环境适应能力强等内在因素的影响, 两属中不乏存在入侵物种[8-9]。
多管藻属广泛分布于世界各海域, 多生长于潮间带或潮下带的岩礁上或潮池中, 或附生于马尾藻()类、蜈蚣藻()类、珊瑚藻()类等大型藻体上。在分类上, 1823年GREVILLE[4]提出的替代了最初与十字花科(Brassicaceae)内属名重复的, 并在同期十几个属名中沿用至今。该属的主要鉴定特征为侧枝原基不产生在连续的关节上、假根不被围轴细胞的侧壁切隔、毛丝体罕见或缺乏、4细胞的果胞枝、精子囊枝取代毛丝体、四分孢子囊群呈直线排列等[10]。目前, 多管藻属在全世界已报道确认了186种及38个种下单位[3], 其中中国记录了20种[2, 11]。由于多管藻属物种的形态特征可塑性高, 种间形态差异小导致同物异名及错误鉴定问题。近年来, 为理清多管藻属物种界定, 基于DNA分子片段的系统发育分析被逐步开展[4, 12-15], 但因属内物种过于繁多, 分子数据库有待于进一步完善。
新管藻属是KIM等[10]于1999年从多管藻属中独立出来的, 该类群多附生于潮间带大型海藻上, 如鼠尾藻()、江蓠()、石花菜()、刚毛藻()、刺松藻((Suringar) Hariot)等。该属的主要鉴定特征为在连续的关节上产生侧枝原基和毛丝体、假根被侧壁切隔、毛丝体丰富、3细胞的果胞枝、精子囊枝产生于毛丝体第二节的一分枝上、四分孢子囊群呈螺旋排列等[10]。目前AlgaeBase中记录了49种及2个种下单位[3], 中国记录了14种, 包括2个新种[2, 4]。2017年DÍAZ-TAPIA[16]等基于分子系统发育分析及形态学数据, 将新管藻属中大部分物种移入, 其余移回多管藻属及冠管藻属()。新管藻属建属时间较晚, 且与多管藻属、及其他亲缘属的划分仍多具争议, 研究尚浅。中国记录的该属物种是否应移入仍需补充形态及分子数据加以分析支持。
中国关于多管藻类的分类学研究已进行了几十年。曾呈奎院士[17]曾于1944年报道了6个多管藻类物种, 而后NODA[18]报道了3个多管藻属物种; 项斯端[19]依据KIM等[10]的分类观点, 报道了包括曾呈奎[17]及NODA[18]的报道在内的多管藻属16种及新管藻属11种; 《中国海藻志》收录并报道了包括之前研究在内的两属物种共30种, 而后夏邦美等[11]报道了一新种即海南多管藻(Xia et Wang)。近年来, 作者对红藻门仙菜目做了一系列分类学研究工作, 如仙菜科(Ceramiaceae)、松节藻科(Rhodomelaceae)爬管藻属()等[20-24], 但国内对于多管藻类的研究主要集中在东南沿海, 如地理分布的扩展[24]、新种和新记录的报道以及8个物种分子数据的补充[4, 25]等, 而有关渤海西岸的研究较少。作者在分类鉴定采集自河北秦皇岛的大型海藻样品过程中, 对其多管藻属及新管藻属样品的形态学特征进行了详细观察, 将为该地区藻类资源补充数据。
1 材料与方法
1.1 材料
实验材料为2019年5月—2021年9月采自河北秦皇岛求仙入海处及老龙头的多管藻属及新管藻属海藻样品, 将样品制作为标本, 分为液浸标本(用95%酒精预处理, 并用6%福尔马林海水浸泡)、腊叶标本(用于观察藻体宏观形态)和干燥标本(用变色硅胶及吸水纸阴干后, 放入–20℃冰箱中冻存), 并相互对应, 编号保存于天津师范大学生命科学学院藻类实验室。样品采集信息见表1, 河北省秦皇岛市求仙入海处和山海关老龙头景区的经纬度分别为119°37′24.99″E、39°54′45.60″N和119°48′6.24″E、39°58′21.83″N。
1.2 方法
1.2.1 外形特征比较
使用体视显微镜Nikon SMZ25观察测量并拍照记录藻体的基部、分枝方式、小枝顶端、毛丝体、四分孢子囊群排列方式、囊果外形等形态特征。
1.2.2 内部特征比较
挑选合适的液浸标本, 利用生物冷冻切片机进行横切, 或对选定位置进行压片, 用0.5%苯胺蓝染液染色, 在正置荧光显微镜Leica DM5000B下观察藻体的内部结构, 如皮层、围轴细胞个数等。
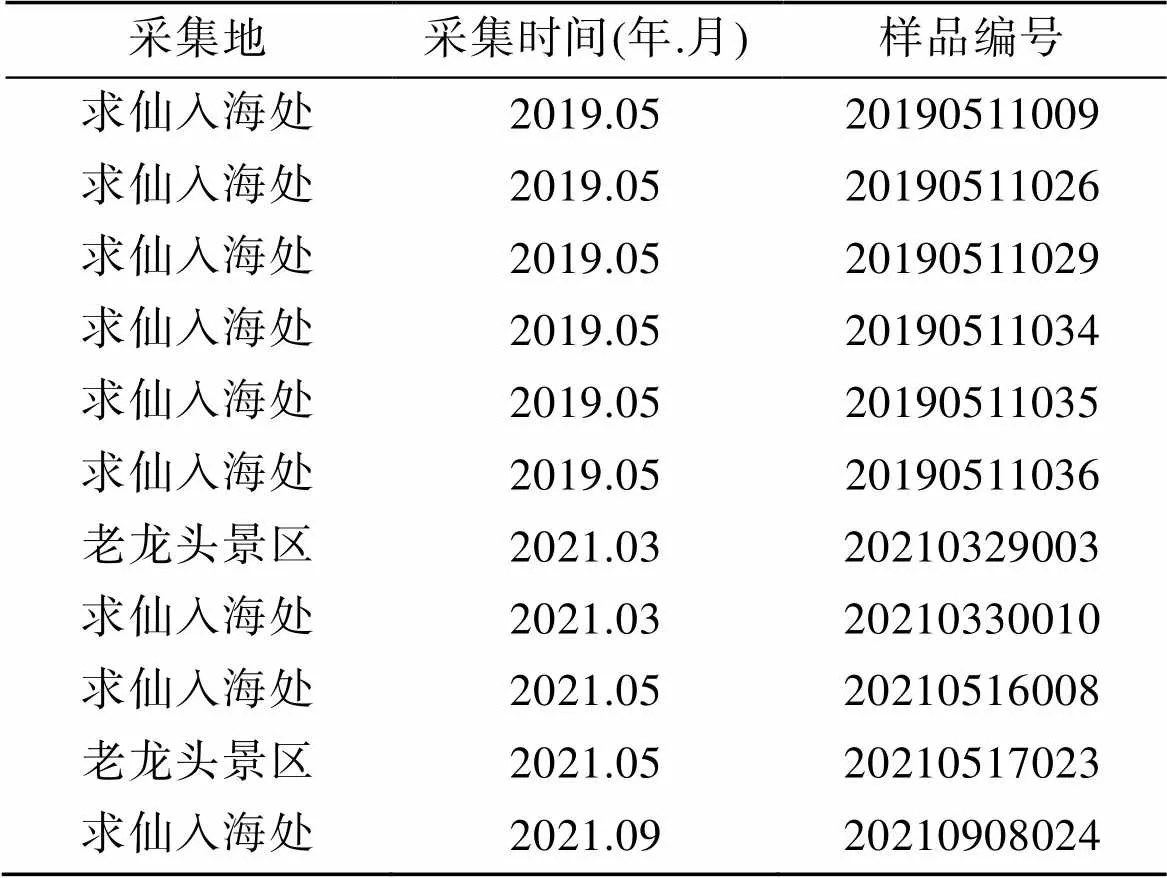
表1 样品采集信息表
1.2.3 物种鉴定
查阅多管藻类的分类学相关文献、书籍及数据等, 比对实验所得样品特征, 进行分析与物种鉴定。
2 实验结果
2.1 多管藻(图1)
多管藻(Harvey)[26], 1862, p. 169; SEGI[27], 1960, p. 616; NELSON等[28], 1996, p. 449-451, figs. 1~2; 项斯端[19], 2004, p. 89;KIM等[29], 2011, p. 118~122, figs. 1~2; 谭华强[4], 2017, p. 40~44, figs. 3~8, 3~9.
(Harvey) Kylin[30], 1941, p. 35.
Hollenberg[31], 1942, p. 774.
鉴定特征: 藻体直立无皮层, 基部具匍匐枝, 假根不被侧壁切隔, 4围轴细胞, 小枝先端具芒尖, 毛丝体罕见, 四分孢子囊群直列, 具1~2个腋生四分孢子囊枝, 坛状囊果等[2, 4]。
特征描述: 藻体红棕色, 直立, 丛生, 高5~16 cm(图1a、1b)。围轴细胞4个(图1c、1d), 轴切面略方形, 无皮层, 藻体中上部围轴细胞稍螺旋(图1e)。基部具有匍匐枝及假根(图1f)。匍匐枝直径为100~ 150 μm, 关节长宽(围轴细胞的长: 围轴的宽)比为1~1.2。假根直径35~60 μm, 不被侧壁切隔, 末端有或无盘状吸胞(图1g), 在直立枝中上部的枝间有假根粘连(图1h)。直立枝细长, 基部至中下部直径为150~270 μm, 近基部关节长宽比为0.3~1, 中下部关节长宽比为5~13; 上部直径为100~150 μm, 关节长宽比为1~1.7; 末端小枝直径为20~70 μm, 关节长宽比为0.2~0.7。有限小枝顶细胞末端具芒尖, 幼枝顶细胞钝圆, 均横裂生长(图1i)。藻体直立枝基部向下产生匍匐枝(图1f), 向上每4~7个关节外生侧枝, 且下部外生侧枝具3~5个小分枝, 小分枝强烈反曲呈钩刺状(图1j、1k); 中上部为假二叉状分枝; 上部互生小枝, 且具有1~2个腋生小枝(图1e、1h、1k)。未见毛丝体及痕细胞。

图1 多管藻
注: a. 四分孢子囊株; b. 囊果株; c. 直立枝横切面; d. 分枝横切面; e. 藻体中上部; f. 匍匐枝及假根; g. 假根; h. 枝间假根(黑色箭头)及腋生小枝(白色箭头); i. 小枝; j. 直立枝下部分枝呈钩刺状反曲; k. 直立枝下部分枝及腋生小枝; l. 腋生四分孢子囊枝; m. 腋生四分孢子囊枝(黑色箭头)和四分孢子囊(白色箭头); n. 囊果枝; 比例=1000 μm (a, b, f, j); 100 μm (c、d、g、h、m); 200 μm (e、i、l、n); 500 μm (k)。
四分孢子囊球状, 直径50~80 μm, 4~7个直列于末位枝中下部, 或于1~2个腋生四分孢子囊枝上(图1l、1m)。囊果坛状, 具长颈, 长450~650 μm, 宽320~ 500 μm, 口宽120~250 μm, 生于小枝上(图1n)。本实验样品中未发现雄性植株。
习性: 生长在潮间带的岩石上或潮池中。
国内产地: 辽宁旅顺, 河北秦皇岛, 山东青岛、荣成、烟台, 浙江构杞岛、大陈岛。
标本编号: 20190511034, 20190511035, 采集于河北秦皇岛求仙入海处。
模式标本产地: 美国奥卡斯岛。
2.2 丛托多管藻(图2)
丛托多管藻(Harvey)[32], 1857, p. 331; KÜTZING[33], 1864, v. 14, t. 47 a~c; SEGI[34], 1951, p. 244; KIM等[35], 1994, p. 185~192, figs. 1~4; CURIEL等[36], 2002, p. 66~70, figs. 1~10; 夏邦美[2],2011, p. 162~164, figs. 112~113; 谭华强[4], 2017, p. 31~35, figs. 3~4, 3~5.
(Harvey) Kylin[30], 1941, p. 35.
Hollenberg[31], 1942, p. 774.
鉴定特征: 藻体直立无皮层, 基部具匍匐枝, 假根不被侧壁切隔, 4围轴细胞, 小枝先端具芒尖, 毛丝体通常早落且未留下痕细胞, 四分孢子囊群直列, 腋生四分孢子囊枝多达7~8个, 坛状囊果等[2, 4]。
特征描述: 藻体红棕色, 丛生, 直立, 高20~30 cm (图2a)。无皮层, 围轴细胞4个(图2b)。基部具匍匐枝, 直径为80~120 μm, 关节长宽比为1.5~2(图2c)。匍匐枝上生有假根, 假根直径约30~35μm, 长短不一, 自围轴细胞角端向外突出, 不被侧壁切隔, 假根末端有或无盘状吸胞(图2d、2e)。直立枝基部直径约160~250 μm, 中下部关节长宽比约3.5~5, 下部叉状或互生分枝, 分枝较稀疏, 有的枝呈外弯钩状。上部小枝互生, 直径约为50~70 μm, 复羽状排列, 上部小枝及不定枝关节短(图2f、2g)。末位枝极短, 使上部羽状枝外廓若细线状, 细胞直径约为40~55 μm, 关节长宽比约为0.7~0.85(图2h、2 i)。小枝顶细胞先端具芒尖(图2j)。偶见毛丝体, 长可达450 μm, 1~3回分叉, 通常早落且未留下痕细胞(图2k)。
四分孢子囊直径60~85 μm, 3~7个直列着生于粗短梭形的四分孢子囊枝上, 四分孢子囊枝2~7个丛生于末位小枝腋内(图2l、2m)。囊果生于藻体主轴近顶端, 呈坛状, 具颈、宽口、短柄, 未成熟囊果大小约为90~110 μm ×120~150 μm(图2n)。本实验样品中未发现雄性植株。
习性: 生长在低潮带的岩石上或潮池中。
模式标本产地: 日本函馆。
国内产地: 辽宁旅顺、大连, 河北秦皇岛(该地区为本研究新记录地区), 山东烟台、威海、青岛。
样品编号: 20210329003, 20210330010, 20210516008, 20210517023, 20210908024, 采集于河北秦皇岛求仙入海处、老龙头。
2.3 倾伏新管藻(图3)
倾伏新管藻(Segi) KimetLee[10], 1999, p. 279; 项斯端[19], 2004, p. 95; NAM等[37], 2012, p. 65~68, figs. 31~33.
Segi[34], 1951, p. 218, pl. VII, 2, and Text-figs. 17~18; LEE等[38], 1986, p. 324.
Noda[39], 1970, p. 34.
(Segi) Díaz-Tapia et Maggs[16], 2017, p. 7.
鉴定特征: 藻体基部具皮层, 假根被侧壁切隔, 4围轴细胞, 小枝顶细胞钝圆, 毛丝体从顶细胞下一节发生, 四分孢子囊群螺列, 宽坛状囊果等[2]。
特征描述: 藻体红棕色, 单生或少数聚生, 质地柔软, 株高0.8~2 cm(图3a)。藻体基部具皮层, 向上分枝无皮层, 围轴细胞4个(图3b)。直立枝基部直径240~500 μm, 向顶端关节变短, 中上部分枝直径约为100~220 μm, 分枝末端直径约为30~60 μm, 整体关节较短, 关节长宽比一般为0.3~0.5(图3c、3d)。下部分枝互生或对生, 稍上为不规则叉状分枝, 末位枝呈不等钳状: 一为锥形略直, 旁边一稍细内弯(图3e~ 3g)。顶细胞钝圆, 其下一节即发生毛丝体, 毛丝体1~3回分叉(图3h、3i)。
四分孢子囊呈球形, 65~105 μm, 四面体分裂, 4~11个连续或间断地螺旋排列于末位枝及次末位枝的节上, 成熟时, 次末位枝上的先成熟, 并使小枝左右膨突扭曲(图3i、3j)。囊果侧生于直立枝中上部, 成熟囊果宽坛状, 长200~350 μm, 宽250~350 μm, 具宽口, 弯柄(图3k、3l)。本实验样品中未发现雄性植株。

图2 丛托多管藻
注: a. 藻体; b. 围轴细胞(P); c. 匍匐枝; d~e. 假根; f~g. 囊果株上部; h~i. 末位枝; j. 小枝顶细胞(箭头); k. 毛丝体(箭头); l~m. 腋生四分孢子囊枝(箭头); n. 囊果; 比例=2 cm (a); 200 μm (b, d, f, m); 500 μm (c); 100 μm (e, i, k); 250 μm (g, h, l); 50 μm (j, n)。

图3 倾伏新管藻
注: a. 藻体; b. 围轴细胞(P); c. 藻体中上部; d. 分枝; e. 藻体下部; f. 分枝; g. 末位枝; h. 毛丝体(箭头); i. 毛丝体(细箭头)及四分孢子囊(粗箭头); j. 四分孢子囊(箭头); k~l. 囊果; 比例=1 000 μm (a, g, k); 500 μm (b, d, e, f, h, i, l); 1 500 μm (c); 200 μm (j)。
习性: 附生于大型海藻上, 本实验样品附生于石花菜上。
模式标本产地: 日本岛根县美保关。
国内产地: 山东, 河北(本研究为该种新记录地区), 浙江, 福建, 广东。
样品编号: 20190511009, 20190511036, 采集于河北秦皇岛求仙入海处。
2.4 球果新管藻(图4)
球果新管藻(Børgesen) Kim et Lee[10], 1999, p. 280; 项斯端[19], 2004, p. 94.
Børgesen[40], 1918, p. 271, fig. 267~271; TAYLOR[41], 1960, p. 576; HOLLENBERG[42], 1968, p. 87~89; LEE等[38], 1986, p. 325; SILVA等[43], 1996, p. 546; ABBOTT[44]1999, p. 428, fig. 127A-G.
(Børgesen) Díaz- Tapia et Maggs[16], 2017, p. 9.
鉴定特征: 藻体无皮层, 假根被侧壁切隔, 4围轴细胞, 小枝顶细胞钝圆, 毛丝体发达, 四分孢子囊群螺列, 球状囊果等[2]。
特征描述: 藻体棕色, 直立, 丛生, 高0.8~2 cm, 下部具局限伸展的匍匐枝, 直立枝直径约300 μm,节处稍膨突(图4a)。藻体多为假二叉分枝, 主枝上互生分枝, 且具有不定小枝(图4b、4c), 上部易断, 中上部直径约为150~190 μm, 末位枝枝端多为不等叉状, 直径约为30~60 μm, 整体关节长宽比约为1~1.5(图4d)。顶细胞钝圆, 直径约为30~40 μm(图4e), 毛丝体发达, 无色, 3~4回分叉, 细胞单核(图4f、4g)。藻体无皮层, 4个围轴细胞(图4h), 主轴围轴细胞长约为330~370 μm, 宽约为80~140 μm, 中轴细胞与围轴细胞近等长。
四分孢子囊球形, 四面体分裂, 直径约为40~85 μm, 5个至多个螺旋排列于末位枝及次末位枝上, 使枝稍扭曲(图4i)。囊果呈球状, 直径约250~400 μm, 口狭呈孔状(图4j、4k)。本实验样品中未发现雄性植株。
习性: 附生于大型海藻上。
模式标本产地: 美国维尔京托马斯岛。
国内产地: 河北(本研究为该种新记录地区), 香港, 海南。
样品编号: 20190511026, 20190511029, 采集于河北秦皇岛求仙入海处。
3 讨论
本研究对采集于河北秦皇岛的样品进行了形态学观察与分析, 鉴定出多管藻类4种, 其形态特征与其他产地的记录基本相符[28, 36-37, 42, 45-53](表2)。

图4 球果新管藻
注: a. 藻体; b. 分枝; c. 不定小枝; d. 末位枝; e. 顶细胞; f~g. 毛丝体; h. 围轴细胞(P); i. 四分孢子囊(箭头); j. 囊果株; k. 囊果; 比例=1 500 μm (a); 500 μm (b, c, d, k); 100 μm (e, f, g, h); 200 μm (i); 1 000 μm (j)。

多管藻最初由HARVEY[26]于1862年报道为美国西北海岸的特有物种, 后在东亚(日本[27, 45, 54]、中国[2, 4, 19]、韩国[29, 37])、大洋洲(新西兰[28]、澳大利亚[46])和欧洲(荷兰[55])等地均有报道。其中在中国辽宁、河北、山东、浙江及广东地区均有分布[2, 4, 19, 56]。本研究样品(编号: 20190511034, 20190511035)特征与夏邦美[2]及谭华强[4]的描述基本相符, 故鉴定为多管藻。同时, 本研究样品特征也与WOMERSLEY[46]的报道相似, 但WOMERSLEY[46]报道本种的分枝模式为不规则分枝, 作者认为可能是因其藻体下部分枝没有规则、中上部不是典型的二叉分枝且生有不定枝的原因; WOMERSLEY[46]描述的囊果略小, 可能不够成熟, 或有地区差异; WOMERSLEY[46]没有描述腋生四分孢子囊枝, 但承认KYLIN[30]的内源性腋生枝的存在及其分类学意义。
丛托多管藻分布于中国黄渤海海域, 并常见于韩国[35, 37, 57]及日本[34, 47, 58], 在俄罗斯[59]、意大利[36]、新西兰[60]、阿根廷[61]及不列颠群岛[62]等地也有报道。本研究样品(编号: 20210329003, 20210330010, 20210516008, 20210517023, 20210908024)特征与夏邦美[2]及谭华强[4]的描述基本相符, 并偶见毛丝体, 1~3回分叉, 长可达450 μm, 鉴定为丛托多管藻。CURIEL等[36]从1999年12月—2000年7月定期进行野外采集, 全面的观察描述了丛托多管藻的形态特征, 本研究样品包括营养植株、四分孢子囊株及囊果株, 特征也与CURIEL等的描述相近, 尤其是四分孢子囊株, 而囊果因不成熟而略小。
多管藻与丛托多管藻因受株龄和环境影响形态变化大, 常出现同物异名、错误鉴定等问题。目前被普遍接受的种级区分特征为腋生四分孢子囊枝的数量, 但该特征在藻体不同生活史阶段会发生变化。KUDO等[58]观察了不同生长周期丛托多管藻的腋生四分孢子囊枝数量变化, 少则1个, 多则7~8个, 变异程度大。因此, 针对腋生四分孢子囊枝数量这个特征, 应选择完全成熟的植株作为鉴定材料[47], 多管藻为1~3个, 丛托多管藻为7~8个。精子囊枝不育顶细胞的数量也是一个种级区分特征[36], 有报道多管藻为1~3个[2, 19]或5~7个[46], 丛托多管藻为5~8个[35]或2~6个[47]。然而, NELSON等[28]没有描述多管藻的不育顶细胞数量,但其图片中清晰可见为6个顶细胞(NELSON等, 1996, p. 450, fig. 2B), 与WOMERSLEY[46]的描述相符, 但与项斯端[19]及夏邦美[2]的描述有明显差异, 考虑因南北半球的地理差异, 生境可能对其特征产生了影响。因此, 精子囊枝不育顶细胞的数量作为分类鉴定特征还有待继续补充数据加以支持。这两个物种也会被误认为是繁枝多管藻((Mertens ex Dillwyn) Greville)或(Lightfoot ex Dillwyn) Greville, 后者目前被认为是繁枝多管藻的同物异名[63-64], 其区别鉴定特征是繁枝多管藻藻体较矮, 且成熟的四分孢子囊株不具有腋生四分孢子囊枝, 精子囊枝不育顶细胞较少[65]。与多管藻类似的淡盐多管藻(Montagne), 毛丝体长而纤细, 且通常有1个二叉, 没有腋生四分孢子囊枝及精子囊枝不育顶细胞[12]。
倾伏新管藻由KIM和LEE于1999年从多管藻属转移至新管藻属, 分布于中国[2, 66]、日本[67]及韩国[37, 68]。夏邦美[2]认为该物种与日本新管藻(Harvey) KIM等相似, 区别在于分枝方式、囊果形状、毛丝体发生位置, 以及倾伏新管藻的精子囊枝具1个不育顶细胞, 而日本新管藻不具有。而NAM等[37]记录描述了日本新管藻的精子囊枝具有1个不育顶细胞。故本研究认为精子囊枝不育顶细胞的数量作为分类鉴定特征还有待继续补充数据加以支持。同时, 本研究认为倾伏新管藻与日本新管藻在藻体大小上有明显差异, 而与细小新管藻((Hariot) KIM et LEE)及球果新管藻((Børgesen) KIM et LEE)类似, 不过后二者藻体无皮层、分枝方式不同且囊果球状, 不难区分。本研究样品(编号: 20190511009, 20190511036)特征与夏邦美[2]的描述基本相符, 故鉴定为倾伏新管藻。
球果新管藻分布广泛, 在亚洲[2, 37, 66-68]、东南亚[69-70]、太平洋[71]、北美洲[53]、大西洋[72]、欧洲[73]、非洲[73]等各地区均有报道。本研究样品与谭华强[4]描述的日本新管藻类似, 但日本新管藻基部具皮层、四分孢子囊群微螺列, 而本研究样品藻体无皮层、四分孢子囊群螺列且间断; 本样品也与AFONSO- CARRILLO[74]等描述的(Bailey) Kim, Choi, Guiry et Saunders类似, 但藻体基部具皮层、毛丝体稀少、囊果具宽口, 而本研究中藻体无皮层、毛丝体发达、囊果口狭, 且目前在日本、韩国附近海域及中国黄渤海地区缺乏形态学报道, 有待继续补充数据用以参考。本研究样品的形态特征与夏邦美[2]及NAM等[37]对球果新管藻的描述基本相符, 故鉴定为球果新管藻。
多管藻属及新管藻属物种的形态特征十分丰富, 有的种间差异极小, 且同一物种也可能因环境或成熟度不同而出现特征差异, 这都可能造成同物异名、错误鉴定等问题, 给两属海藻的物种鉴定、位置划分和数量统计等相关研究工作带来极大困难。近年来, 越来越多的学者通过分子系统发育学分析, 研究种属间的遗传距离差异, 以此理清多管藻属及新管藻属混乱的物种鉴定及位置划分问题[4, 12-15, 16]。但因物种众多而缺乏通用引物等原因, 很多物种的分子数据并不完善, 例如本研究涉及的多管藻在GenBank中只收录到3个23和1个28rRNA序列片段, 以及1个18rRNA全序列, 而丛托多管藻收录的多为L和cox1序列。不过, 近年来GenBank收录到丛托多管藻的1个18rRNA全序列, 这有助于系统发育关系的全面分析, 更多数据有待继续补充完善。DÍAZ-TAPIA[16]等基于分子及形态学数据, 将新管藻属中的33个种移入, 其中就包括本研究涉及的倾伏新管藻和球果新管藻。采集自我国沿海的以上两个新管藻属物种乃至整个属是否应移入, 仍需进一步研究, 补充相关数据加以支持验证。
4 结论
本研究利用显微观察及生物冷冻切片技术, 鉴定出中国渤海西岸多管藻类物种共2属4种, 即多管藻、丛托多管藻、倾伏新管藻和球果新管藻, 其中丛托多管藻、倾伏新管藻及球果新管藻均为渤海西岸首次记录。本研究结果补充了中国渤海西部的海藻资料, 为后续相关研究提供基础数据与参考。
[1] ARESCHOUG J E. Phycearum, quae in maribus Scandinaviae crescunt, enumeration: sectio prior Fucaceas continens[J]. Nova Acta Regiae Societatis Scientiarum Upsaliensis, 1847, 13: 223-382.
[2] 夏邦美. 中国海藻志. 第2卷. 红藻门. 第7册, 仙菜目松节藻科[M]. 北京: 科学出版社, 2011.
Xia Bangmei. Flora algarum marinarum sinicarum Tomus II Rhodophyta No. VII Ceramiales Rhodomelaceae[M]. Beijing: Science Press, 2011.
[3] GUIRY M D, GUIRY G M. AlgaeBase: World-wide electronic publication[DB/OL]. Galway: National University of Ireland. [2022-10-05]. http://www.algaebase. org.
[4] 谭华强. 基于形态特征与分子数据的中国广义多管藻属分类学研究[D]. 汕头: 汕头大学, 2017.
TAN Huaqiang. A taxonomic study on(Rhodomelaceae, Rhodophyta) in China based on morphological characters and molecular data[D]. Shantou: Shantou University, 2017.
[5] 赵小亮. 利用糖芯片研究海洋多糖及其衍生物与蛋白质的相互作用[D]. 青岛: 中国海洋大学, 2013.
ZHAO Xiaoliang. The interactions of marine polysaccharides and their derivatives with proteins by carbohydrate microarray[D]. Qingdao: Ocean University of China, 2013.
[6] ZHAO X L, JIAO G L, YANG Y, et al. Structure and immunomodulatory activity of a sulfated agarose with pyruvate and xylose substitutes fromHarvey[J]. Carbohydrate Polymers, 2017, 176: 29-37.
[7] WANG P R, CHEN J P, CHEN L J, et al. Characteristic volatile composition of seven seaweeds from the Yellow Sea of China[J]. Marine Drugs, 2021, 19(4): 192.
[8] WILLIAMS S L, SMITH J E. A global review of the distribution, taxonomy, and impacts of introduced seaweeds[J]. Annual Review of Ecology Evolution and Systematics, 2007, 38: 327-359.
[9] MCIVOR L, MAGGS C A, PROVAN J, et al.L sequences reveal multiple cryptic introductions of the Japanese red alga[J]. Molecular Ecology, 2001, 10(4): 911-919.
[10] KIM M S, LEE I K.gen. et sp. nov. with a taxonomic reassessment of the genus(Rhodomelaceae, Rhodophyta)[J]. Phycological Research, 1999, 47: 271-281.
[11] 夏邦美, 王永强. 中国多管藻属一新种[J]. 海洋与湖沼, 2012, 43(5): 963-966.
XIA Bangmei, WANG Yongqiang. A new species of(Rhodophyta) from China[J]. Oceanologia Et Limnologia Sinica, 2012, 43(5): 963-966.
[12] MAMOOZADEH N R, FRESHWATER D W.(Ceramiales, Florideophyceae) species of Caribbean Panama includingsp. nov. andsp. nov.[J]. Botanica Marina, 2012, 55(4): 317-347.
[13] DÍAZ-TAPIA P, KIM M S, SECILLA A, et al. Taxonomic reassessment of(Rhodomelaceae, Rhodophyta) and similar species, including, a newly introduced species in Europe[J].European Journal of Phycology, 2013, 48(4): 345-362.
[14] KIM B, KIM M S. Three new species of(Rhodophyta) based on the morphology and molecular evidence[J]. Algae, 2014, 29(3): 183-195.
[15] BUSTAMANTE D E, WON B Y, CHO T O.sp. nov. andsp. nov.: two new species of(Rhodomelaceae, Rhodophyta) from Korea[J]. European Journal of Phycology, 2015, 50(3): 330-342.
[16] DÍAZ-TAPIA P, MCIVOR L, FRESHWATER D W, et al. The generaBornet & Falkenberg andS.F. Gray constitute well-defined clades of the red algal tribe Polysiphonieae (Rhodomelaceae, Ceramiales)[J]. European Journal of Phycology, 2017, 52(1): 1-20.
[17] TSENG C K. Marine algae of Hong Kong. VI. The genus[J]. Papers of the Michigan Academy of Sciences, Arts and Letters, 1944, 29: 67-82.
[18] NODA M. Marine algae of north-eastern China and Korea[J]. Science Reports of Niigata University Series D (Biology), 1966, 3: 19-85.
[19] 项斯端. 中国多管藻属及新管藻属的研究[J]. 浙江大学学报(理学版), 2004, 31(1): 88-97.
XIANG Siduan. Study onandin China[J]. Journal of Zhejiang University (Science Edition), 2004, 31(1): 88-97.
[20] 刘美媛, 闫盼竹, 王雪聪, 等. 河北秦皇岛仙菜科Ceramiaceae红藻的分类研究[J]. 广西科学, 2022, 29(1): 147-157.
LIU Meiyuan, YAN Panzhu, WANG Xuecong, et al. Classification study of family Ceramiaceae (Rhodophyta, Ceramiales) from Qinhuangdao, Hebei[J]. Guangxi Sciences, 2022, 29(1): 147-157.
[21] 丁兰平, 刘美媛, 闫盼竹, 等. 红藻门仙菜科Ceramiaceae的分类现状与展望[J]. 广西科学, 2022, 29(1): 131-146.
DING Lanping, LIU Meiyuan, YAN Panzhu, et al. Classification status and prospects of the family Ceramiaceae, Rhodophyta[J]. Guangxi Sciences, 2022, 29(1): 131-146.
[22] 王艺晓, 姜晶晶, 丁兰平, 等. 中国海洋仙菜科(Ceramiaceae)新记录属——盖氏藻属(红藻门)的分类研究[J]. 海洋与湖沼, 2020, 51(2): 328-334.
WANG Yixiao, JIANG Jingjing, DING Lanping, et al. Study on taxonomy of marine red algal, a new record genus of family Ceramiaceae from Chinese coasts[J]. Oceanologia Et Limnologia Sinica, 2020, 51(2): 328-334.
[23] DING L P, TAN H Q, ZHANG Q L, et al. Morphological study of the genus(Rhodophyta, Rhodomelaceae) on the coast of eastern Guangdong, China, with a description ofsp. nov.[J]. Chinese Journal of Oceanology and Limnology, 2016, 34(2): 271-282.
[24] 丁兰平, 钟丽敏, 谭华强, 等. 海洋红藻岩生多管藻绒毛变种var.形态学研究[J]. 热带海洋学报, 2015, 34(1): 42-48.
DING Lanping, ZHONG Limin, TAN Huaqiang, et al. A morphological study on marine red algalvar.[J]. Journal of Tropical Oceanography, 2015, 34(1): 42-48.
[25] TAN H Q, LIU W H, LI P. Taxonomic reassessment of(Rhodomelaceae, Rhodophyta) based on molecular and morphological analyses[J]. Journal of Oceanology and Limnology, 2021, 39(5): 1077-1084.
[26] HARVEY W H. Notice of a collection of algae made on the northwest coast of North America, chiefly at Vancouver Island, by David Lyall, Esq, M D R N, in the years 1859-1861[J]. Journal of the Proceedings of the Linnean Society of London Botany, 1862, 6(24): 157-177.
[27] SEGI T. Further study offrom Japan (II)[J]. Report of the Faculty of Fisheries, Prefectural University of Mie, 1960, 3: 608-626.
[28] NELSON W A, MAGGS C A. Records of adventive marine algae in New Zealand:,(Ceramiales, Rhodophyta) and(Dictyosiphonales, Phaeophyta)[J]. New Zealand Journal of Marine and Freshwater Research, 1996, 30: 449-453.
[29] KIM B H, NAM K W. First record ofHarvey (Ceramiales: Rhodophyta) in Korea[J]. The Korean Society of Fisheries and Aquatic Science, 2011, 14(2): 118-122.
[30] KYLIN H. Californische Rhodophyceen[J]. Acta Universitatis Lundensis, 1941, 37(2): 1-71.
[31] HOLLENBERG G J. An account of the species ofon the Pacific coast of North America. I.[J]. American Journal of Botany, 1942, 29: 772-785.
[32] HARVEY W H. Algae[C]//GRAY A. Account of the botanical specimens. Washington: Senate of the Thirty- third Congress, Second Session, Executive Document, 1857: 331-332.
[33] KÜTZING F T. Tabulae phycologicae; oder, Abbildungen der Tange. Vol. XIV[M]. Nordhausen: Gedruckt auf kosten des Verfassers, 1864.
[34] SEGI T. Systematic study of the genusfrom Japan and its vicinity[J]. Journal of the Faculty of Fisheries, Prefectural University of Mie, 1951, 1: 167-272.
[35] KIM M S, LEE I K, BOO S M. Morphological studies of the red algaHarvey on the Korean coast[J]. Korean Journal of Phycology, 1994, 9: 185-192.
[36] CURIEL D, BELLEMO G, ROCCA B L, et al. First report ofHarvey (Ceramiales, Rhodophyta) in the Mediterranean Sea[J]. Botanica Marina, 2002, 45(1): 66-70.
[37] NAM K W, KANG P J. Algal flora of Korea. Volume 4, Number 4. Rhodophyta: Ceramiales: Rhodomelaceae: 18 genera including[M]. Incheon: National Institute of Biological Resources, 2012.
[38] LEE I K, KANG J W. A check list of marine algae in Korea[J]. Korean Journal of Phycology, 1986, 1(1): 311-325.
[39] NODA M. Some marine algae collected on the coast of Iwagasaki, Prov. Echigo facing the Japan Sea[J]. Science Reports of Niigata University, Series D (Biology), 1970, 7: 27-35.
[40] BØRGESEN F. The marine algae of the Danish West Indies. Part 3. Rhodophyceae (4)[J]. Dansk Botanisk Arkiv, 1918, 3: 241-304.
[41] TAYLOR W R. Marine algae of the eastern tropical and subtropical coasts of the Americas[M]. Ann Arbor: The University of Michigan Press, 1960.
[42] HOLLENBERG G J. An account of the species ofof the central and western tropical Pacific Ocean. I.[J]. Pacific Science, 1968, 22: 56-98.
[43] SILVA P C, BASSON P W, MOE R L. Catalogue of the benthic marine algae of the Indian Ocean[J]. University of California Publications in Botany, 1996, 79: 1-1259.
[44] ABBOTT I A. Marine red algae of the Hawaiian Islands[M]. Honolulu, Hawaii: Bishop Museum Press, 1999.
[45] KUDO T, MASUDA M. Taxonomic notes onHarvey andHollenberg (Ceramiales, Rhodophyta)[J]. Japanese Journal of Phycology, 1988, 36: 138-142.
[46] WOMERSLEY H B S. The marine benthic flora of southern Australia-Part IIID Ceramiales-Delesseriaceae, Sarcomeniaceae, Rhodomelaceae[M]. Canberra & Adelaide: Australian Biological Resources Study & State Herbarium of South Australia, 2003.
[47] KUDO T, MASUDA M. Taxonomic features ofHarvey (Ceramiales, Rhodophyta)[J]. Korean Journal of Phycology, 1992, 7(1): 13-26.
[48] KIM M S, YANG E C, MANSILLA A, et al. Recent introduction of(Ceramiales, Rhodophyta) to Punta Arenas, Chile[J]. Botanica Marina, 2004, 47: 389-394.
[49] PIÑEIRO-CORBEIRA C, VERBRUGGEN H, DÍAZ- TAPIA P. Molecular survey of the red algal family Rhodomelaceae (Ceramiales, Rhodophyta) in Australia reveals new introduced species[J]. Journal of Applied Phycology, 2020a, 32(4): 2535-2547.
[50] KIM M S. Taxonomic reassessment of the genus(Rhodomelaceae, Rhodophyta): morphology and reproduction ofand[J]. Algae, 2003, 18(2): 107-119.
[51] GUIMARÃES S M P B, FUJI M T, PUPO D, et al. Reavaliação das características morfológicas e suas implicações taxonômicas no gênero(Ceramiales, Rhodophyta) do litoral dos Estados de São Paulo e Espírito Santo, Brasil[J]. Revista Brasileira de Botanica, 2004, 27(1): 163-183.
[52] N′YEURT A D R, PAYRI C E. Marine algal flora of French PolynesiaⅢ. Rhodophyta, with additions to the phaeophyceae and Chlorophyta[J]. Cryptogamie Algologie, 2010, 31(1): 3-2.
[53] MAMOOZADEH N R, FRESHWATER D W. Taxonomic notes on Caribbeanand(Ceramiales, Florideophyceae): five species from Florida, USA and Mexico[J]. Botanica Marina, 2011, 54(3): 269-292.
[54] YOSHIDA T. Marine algae of Japan[M]. Tokyo: Uchida Rokakuho Publishing Co., Ltd, 1998.
[55] STEGENGA H, MOL I, PRUD'HOMME VAN REINE W F, et al. Checklist of the marine algae of the Netherlands[J]. Gorteria Supplement, 1997, 4: 3-57.
[56] 曾呈奎. 中国黄渤海海藻[M]. 北京: 科学出版社, 2009.
ZENG Chengkui. Seaweeds in Yellow Sea and Bohai Sea of China[M]. Beijing: Science Press, 2009.
[57] BOO S M, KO Y D. Marine plants from Korea[M]. Seoul: Marine & Extreme Genome Research Centre Program, 2012.
[58] KUDO T, MASUDA M. A taxonomic study ofHarvey (Rhodophyta, Ceramiales)[J]. Japanese Journal of Phycology, 1981, 29: 263-272.
[59] PERESTENKO L P. The seaweeds of Peter the Great Bay[M]. Leningrad: NAUKA Leningradskoe Otdelenie, 1980.
[60] D'ARCHINO R, NEILL K F, NELSON W A. Recognition and distribution of(Rhodomelaceae, Rhodophyta) in New Zealand[J]. Botanica Marina, 2013, 56(1): 41-47.
[61] RAFFO M P, GEOFFROY A, DESTOMBE C, et al. First record of the invasive red algaHarvey (Rhodomelaceae, Rhodophyta) on the Pategonian shores of the Southwestern Atlantic[J]. Botanica Marina, 2014, 57(1): 21-26.
[62] PIÑEIRO-CORBEIRA C, MAGGS C A, RINDI F, et al. Molecular assessment of the tribes Streblocladieae and Polysiphonieae (Rhodomelaceae, Rhodophyta) in the British Isles reveals new records and species that require taxonomic revision[J]. Cryptogamie Algologie, 2020b, 41(8): 55-72.
[63] KIM M S, MAGGS C A, MCIVOR L, et al. Reappraisal of the type species of(Rhodomelaceae, Rhodophyta)[J]. European Journal of Phycology, 2000, 35(1): 83-92.
[64] MAGGS C A, HOMMERSAND M H. Seaweeds of the British Isles. Volume 1. Rhodophyta. Part 3A. Ceramiales[M]. London: HMSO, 1993.
[65] DÍAZ-TAPIA P, BÁRBARA I. Seaweeds from sand- covered rocks of the Atlantic Iberian Peninsula. Part 1. The Rhodomelaceae (Ceramiales, Rhodophyta)[J]. Cryptogamie Algologie, 2013, 34(4): 325-422.
[66] 刘瑞玉. 中国海洋生物名录[M]. 北京: 科学出版社, 2008.
LIU Ruiyu. Checklist of Marine Biota of China Seas[M]. Beijing: Science Press, 2008.
[67] YOSHIDA T, SUZUKI M, YOSHINAGA K. Checklist of marine algae of Japan (Revised in 2015)[J]. Japanese Journal of Phycology, 2015, 63: 129-189.
[68] LEE Y, KANG S. A catalogue of the seaweeds in Korea[M]. Jeju: Cheju National University Press, 2001.
[69] ABBOTT I A, FISHER J, MCDERMID K J. Newly reported and revised marine algae from the vicinity of Nha Trang, Vietnam[C]//BBOTT I A, MCDERMID K J. Taxonomy of economic seaweeds with reference to some Pacific species. Vol. VIII. Vietnam: California Sea Grant College, 2002: 291-321.
[70] ANG P O, LEUNG S M, CHOI M M. A verification of reports of marine algal species from the Philippines[J]. Philippine Journal of Science, 2013, 142(3): 5-49.
[71] HUISMAN J M, ABBOTT I A, SMITH C M. Hawaiian reef plants[M]. Honolulu: A publication of the University of Hawai'i Sea Grant College Program, 2007.
[72] SCHNEIDER C W. An annotated checklist and bibliography of the marine macroalgae of the Bermuda Islands[J]. Nova Hedwigia, 2003, 76: 275-361.
[73] GÓMEZ GARRETA A, GALLARDO T, RIBERA M A, et al. Checklist of the Mediterranean seaweeds. III. Rhodophyceae Rabenh. 1. Ceramiales Oltm[J]. Botanica Marina, 2001, 44(5): 425-460.
[74] AFONSO-CARRILLO J, SANSÓN M, REYES J, et al. Morfología y distribución de la rodófita alóctona, y comentarios sobre otras algas marinas probablemente introducidas en las Islas Canarias[J]. Revista de la Academia Canaria de Ciencias, 2002, 14: 83-98.
Study on the morphological taxonomy ofComplexfrom the west coast of the Bohai Sea
YAN Pan-zhu, CHU Yue, DING Lan-ping, HUANG Bing-xin, LIU Mei-yuan, WANG Xue-cong, QIN Jia-nan
(Tianjin Key Laboratory of Animal and Plant Resistance, College of Life Science, Tianjin Normal University, Tianjin 300387, China)
Through microscopic observation and biological frozen section techniques, the morphological taxonomy of theComplex collected from Qinhuangdao, Hebei, was studied. The samples were identified as two species of, namelyHarvey andHarvey, and two species of, namely(Segi) Kim et Lee and(Børgesen) Kim et Lee. Among them,,, andwere first recorded on the west coast of the Bohai Sea. This paper describes the external morphology and internal structure characteristics of the samples and presents clear pictures describing the features. The research results supplement the data on algae resources in Bohai Bay and provide a taxonomic reference for the related research on theComplex.
;; Qinhuangdao; taxonomy; red algae
Oct. 5, 2022
Q949.29
A
1000-3096(2023)5-0066-13
10.11759/hykx20221005001
2022-10-05;
2023-01-13
国家自然科学基金面上项目(31970216, 32270219, 31670199)
[National Natural Science Foundation of China, Nos. 31970216, 32270219, 31670199]
闫盼竹(1997—), 女, 天津人, 硕士研究生, 主要从事大型海藻分类学研究, E-mail: yanpanzhu@126.com; 黄冰心(1974—),通信作者, 福建莆田人, 博士, 硕士生导师, 主要从事大型海藻资源及海洋生物技术研究, E-mail: skyhbx@tjnu.edu.cn
(本文编辑: 谭雪静)
猜你喜欢
中国海洋大学学报(自然科学版)(2022年8期)2022-08-15
语文周报·教研版(2021年28期)2021-08-19
现代农业科技(2020年21期)2020-12-14
生物学通报(2019年7期)2019-07-08
科技创新与应用(2017年20期)2017-07-15
数学理论与应用(2017年2期)2017-06-27
数学理论与应用(2017年2期)2017-06-27
新疆农业科学(2016年11期)2016-11-26
数学理论与应用(2016年1期)2016-02-28
西北植物学报(2015年5期)2015-07-05
