
呼伦贝尔沙化草地不同生态修复阶段植物群落特征研究
2023-07-29 13:55:28郝颢晶关潇曹明李俊生
环境工程技术学报 2023年4期
郝颢晶,关潇,曹明,李俊生,2*
1.中国环境科学研究院
2.中国地质调查局自然资源综合调查指挥中心
草地资源作为我国面积最大的陆地生态系统,对水土保持、水源涵养、气候调节等生态系统服务的维持及生物多样性保护均具有重要意义[1]。呼伦贝尔草原作为我国北方主要自然资源及生态防护屏障,具有独特的地理位置和多样的生态功能。随着全球气候变化[2]和人类活动强度的持续加深[3-4],呼伦贝尔草原面临日益严峻的沙化问题[5],威胁着草地生态系统的可持续发展[6]。近年来,呼伦贝尔市加大沙地植被修复治理力度,开展了人工植被修复等一系列草地修复和管理保护措施[7],草地生态环境得到初步修复。甘珠尔沙地位于呼伦贝尔市西南部,地处呼伦贝尔典型沙带之一,其生态区位重要,生态敏感性等级为脆弱区。自2005年呼伦贝尔市采用“围栏封育+建立草方格+补播耐旱植物”的林草、灌草结合的方式对甘珠尔沙地的流动沙丘开展修复工程后,流动沙丘被固定,植被盖度得到明显改善[8-9]。基于此,为促进沙化草地治理进程和提高治理成效,对甘珠尔不同修复年限的沙地开展植物群落研究,具有重要的现实意义。
不同立地条件下,沙化草地人工修复植物群落表现出不同的时间梯度特征。如钱洲等[10]对毛乌素沙地不同修复年限的植被修复特征的研究发现,密度、盖度及生物量有明显增长,同时植被和土壤性质之间存在极为密切的相互促进的相关性;慕宗杰[11]发现,浑善达克沙地不同治理年份的植物群落中灌木层与草本层高度、盖度、多样性的变化趋势和波动幅度存在差异;王明明等[12]对长期封育的科尔沁沙地的植物研究中发现,随着修复年限的增加植被盖度显著提高,生物量和多样性在年际间波动变化无显著趋势;栗忠飞等[13]在呼伦贝尔南部沙带植被修复过程中发现,随植被修复年限的增加地上植被生物量、盖度显著增加;金晓明等[14]对呼伦贝尔市陈巴尔虎旗人工修复后的沙化草地的研究发现,植物群落在修复初期的1~4 a 仍处于不稳定状态。综上,沙地的植被修复的动态变化受到沙地地理位置、修复年限、土壤条件及区域气候等因素的影响,在植被物种组成、结构和多样性等方面表现出差异。
目前关于不同修复年限下呼伦贝尔市甘珠尔沙地的植被研究较为鲜见,针对甘珠尔沙地在长期修复过程中植物群落特征变化、多样性变化及其影响因素的探究还较缺乏。因此,笔者采用以空代时法,以甘珠尔沙地修复区内修复年限分别为9、11、13、15 a 的沙化草地作为研究对象,结合同期气象数据与土壤养分测定数据,探讨分析不同修复年限沙地植物群落特征、多样性变化及其影响因素,有助于深入理解沙化草地植物群落对修复过程的响应,以期为沙化草地的植被修复和生态环境改善提供科学指导。
1 材料与方法
1.1 研究区概况
甘珠尔沙地位于内蒙古自治区呼伦贝尔市新巴尔虎左旗阿木古郎镇(117°55′E~119°22′E,47°59′N~48°43′N)(图1),土壤类型为沙质栗钙土[15]。平均海拔700 m,属于温带半干旱大陆性季风气候,年均气温0.3 ℃,年日照时数为3 076 h,无霜期100 d,积雪期140 d,年主导风向为西北风,年均风速2.8 m/s,年均降水量约287 mm,主要集中在7—8 月,年均蒸发量2 650 mm,雨热同期,有利于夏季植物生长[16]。

图1 研究区采样点示意Fig.1 Location map of the study area and sampling points
进行人工植被修复前,甘珠尔沙地的地貌特征为流动沙丘。自2005年起,甘珠尔沙地开展人工植被修复工作,以灌草混播的方式为主,辅以少量樟子松(Pinus sylvestris)固定流动沙丘并在人工播种后对甘珠尔沙地修复区实施围栏封育。各样地均保持独立封育,期间可排除家畜采食、践踏、施肥等人工干扰,使土壤内养分贮存从而促进修复区内植被自然更新和生产力恢复。开展本研究时,修复区已分布着大量不同修复年限的人工植被,灌木有羊柴(Hedysarum laeve)、小叶锦鸡儿(Caragana microphylla)、盐蒿(Artemisia halodendron),其下草本以羊草(Leymus chinensis)、 披碱草(Elymus dahuricus)、糙隐子草(Cleistogenes squarrosa)、地稍瓜(Cynanchum thesioides)、 虫实(Corispermum hyssopifolium)等植物为主。
1.2 试验设计与数据采集
1.2.1 样地设置
通过实地调查,分别选取甘珠尔沙地修复区内地形地貌、成土母质等土壤环境条件相近,修复起始年限不同的4 个修复样地(2006年、2008年、2010年和2012年),开展植物群落调查及土壤样品的采集。按照修复年限的不同将样地命名为15、13、11、9 a。为克服边缘效应对结果的影响,各样地均距离围栏边界20 m 以上,各样地内自西向东沿植被种植时样带走向,均匀设置5 条平行样带(相邻样带间隔50 m 以上),每条样带上设置3 个重复,每个重复包含5 m × 5 m 灌木样方和1 m × 1 m 草本样方各1 个,共计草本植物样方60 个,灌木样方60 个。
1.2.2 植被调查与土壤采集
2021年8 月下旬,在沙地植被生长旺盛期分别对15、13、11、9 a 人工修复的植被进行采样调查,包括植物群落特征调查和土壤样品采集。野外调查与数据计算参考任继周[17]草原生态调查方法,利用钢卷尺进行测量并记录样方内出现的草本及灌木植物种类、株数、株高、多度、基径、冠幅、分盖度、群落总盖度、频度。齐地刈割样方内的草样,分装后带回室内105 ℃杀青30 min,65 ℃烘干至恒重再称取其地上生物量。计算相对密度(RD)、相对盖度(RV)、相对频度(RF)及重要值(IV)。在刈割后的草本样方内,用直径3 cm 的土钻采集0~20 cm 深度的土壤样品,每个样方内采用对角线五点取样法采集5 个土壤样品并混合均匀,带回室内风干磨碎并过筛。
1.2.3 气象数据
研究区2006—2012年的年均降水量、蒸散发、风速和温度等气象数据来源于国家地球系统科学数据中心(http://www.geodata.cn)。
1.3 测定方法
1.3.1 主要植物群落结构特征值测定
根据已有研究[18-21]得到的生物量模型计算灌木物种地上生物量:
式中:W1、W2 、W3、W4 分别 为羊柴、黄柳、小叶锦鸡儿及盐蒿物种的地上生物量,kg/hm2; DH为断面周长,m;V为冠幅体积,m3;A为冠幅面积,m2。
式中:IV 为各物种重要值;RD 为相对密度,%;RV为相对盖度,%;RF 为相对频度,%。
物种多样性指数主要包括Shannon-Wiener 指数( Sw)、Simpson 指数(D)、Margalef 指数(M)以及Pielou 指数(J),计算公式如下:
式中:Pi为第i物种密度占群落总密度的比例;S为样方物种总数;T为样方中所有物种的个体总数。
1.3.2 土壤测定
参考鲍士旦[22]的土壤分析方法,测定土壤含水量、容重、pH、电导率、总氮含量、总钾含量、总磷含量和有机碳含量。采用烘干称重法测定土壤含水量;环刀法测定土壤容重;电极电位法测定土壤pH 和电导率;凯氏定氮法测定土壤总氮含量;NaOH熔融-原子吸收法测定土壤总钾含量;NaOH 熔融-钼锑抗显色-紫外分光光度法测定土壤总磷含量;重铬酸钾容量法-外加热法测定土壤有机碳含量。
1.4 数据处理
采用Excel 软件对不同修复年限的植物调查数据、环境因子数据进行预处理,基于R 4.1.2 语言进行多样性指数计算及单因素方差分析,并基于Tukey HSD 检验对其进行多重比较。利用R 4.1.2 语言的randomForest 包对物种多样性与环境因子之间的相互关系进行随机森林模型回归分析,基于rfPermute包检验模型中各环境因子的显著性[23],A3 包检验整个模型的显著性[24],通过部分依赖图展现具有显著性的环境因子与生物多样性指数之间的关系。采用Origin 8.0 软件制图。
2 结果与分析
2.1 不同物种组成及优势度
2.1.1 植物科、属、种和生活型组成
研究区域内不同修复阶段的植物群落中,共发现42 种植物,隶属于17 科35 属。其中灌木物种共3 种,分别为羊柴(Hedysarum fruticosum)、小叶锦鸡儿(Caragana microphylla)和盐蒿(Artemisia halodendron);草本植物39 种,隶属于17 科33 属,物种组成以菊科(Asteraceae)、禾本科(Poaceae)、蔷薇科(Rosaceae)和豆科(Leguminosae)为主,占总数的56.4%。藜科(Chenopodiaceae)、蓼科(Polygonaceae)、石竹科(Caryophyllaceae) 及苋科(Amaranthaceae)共8 个物种,占总数的20.5%。单属种9 种,占总数的23.1%。按生活型划分,可分为一年生草本9 种、一/二年生草本2 种和多年生草本28 种。多年生草本物种数明显高于一年生和一/二年生,占比为43.8%~71.4%。
不同修复阶段,群落物种数存在一定差异。草本物种数整体呈现先下降后上升的趋势,表现为26 种(15 a)、23 种(13 a)、16 种(9 a)、13 种(11 a),在各修复年限中,一/二年生草本占草本物种总数的占比与其他生活型相比最小;灌木物种数以豆科为主,在不同修复阶段均有分布。菊科的盐蒿物种仅在9 和11 a 中出现。
2.1.2 物种重要值变化
植物物种的重要值可反映出各修复年限草地植被的整体情况。结果显示,各物种重要值在年际间波动剧烈(图2、图3)。草本植物中地梢瓜(Cynanchum thesioides)、虫实(Corispermum hyssopifolium)、披碱草(Elymus dahuricus)、 糙隐子草(Cleistogenes squarrosa)、 羊草(Leymus chinensis)与狗尾草(Setaria viridis)的重要值明显高于其他物种。9 a 时草本优势度较高的物种依次为地梢瓜(18.3%)、虫实(14.8%)、披碱草(11.9%) 和羊草(11.0%)。同期糙隐子草(3.69%)并未表现出优势。11 a 时地梢瓜(29.4%)与披碱草(22.0%)重要值显著高于其他草本物种。13 a 时地梢瓜被糙隐子草(15.6%)和狗尾草(15.1%)取代,与此同时黄花蒿(Artemisia annua)、羊草与虫实的重要值均超过10.0%,与糙隐子草和狗尾草共同组成该年的优势群落。15 a 时披碱草与狗尾草成为草本最优势物种,重要值分别为18.1%和10.5%。

图2 不同修复年限研究区草本植物重要值雷达图Fig.2 Radar map of important values of herbaceous plants in the study area with different recovery years

图3 不同修复年限研究区灌木植物重要值雷达图Fig.3 Radar map of important values of shrub plants in the study area with different recovery years
灌木物种小叶锦鸡儿在各修复年限间整体表现出具有较高的优势度,其在9 a 时重要值最大为74.8%。11 a 时盐蒿物种分别超过羊柴(26.2%)和小叶锦鸡儿(21.0%)达到最大值(52.8%)。13 a 时小叶锦鸡儿重新成为最优势物种,重要值达到73.3%。15 a 时,未发现盐蒿物种存在,2 种豆科植物羊柴(51.7%)和小叶锦鸡儿(48.3%)共同构成当年的灌木群落。
2.2 盖度、生物量变化
随着修复年限的增加,草本植被盖度整体呈波动增加的趋势,但差异不显著(P>0.05) 〔图4(a)〕。其中修复年限最短的(9 a)总盖度最小(30.5%),在11 a 时植被盖度升至43.0%,在13 a 时降至34.9%,在修复年限最长(15 a)时达到最大值(43.3%)。与修复9 a 相比,11、13 和15 a 草本总盖度分别高出41.2%、14.4%和42.2%。草本地上生物量在不同修复年限之间无显著差异(P>0.05),但与盖度呈现相似的波动趋势。其最大值出现在15 a(33.9 g/m2) ,与9 a的最小值(18.1 g/m2)相比增加了87.0%〔图4(b)〕。
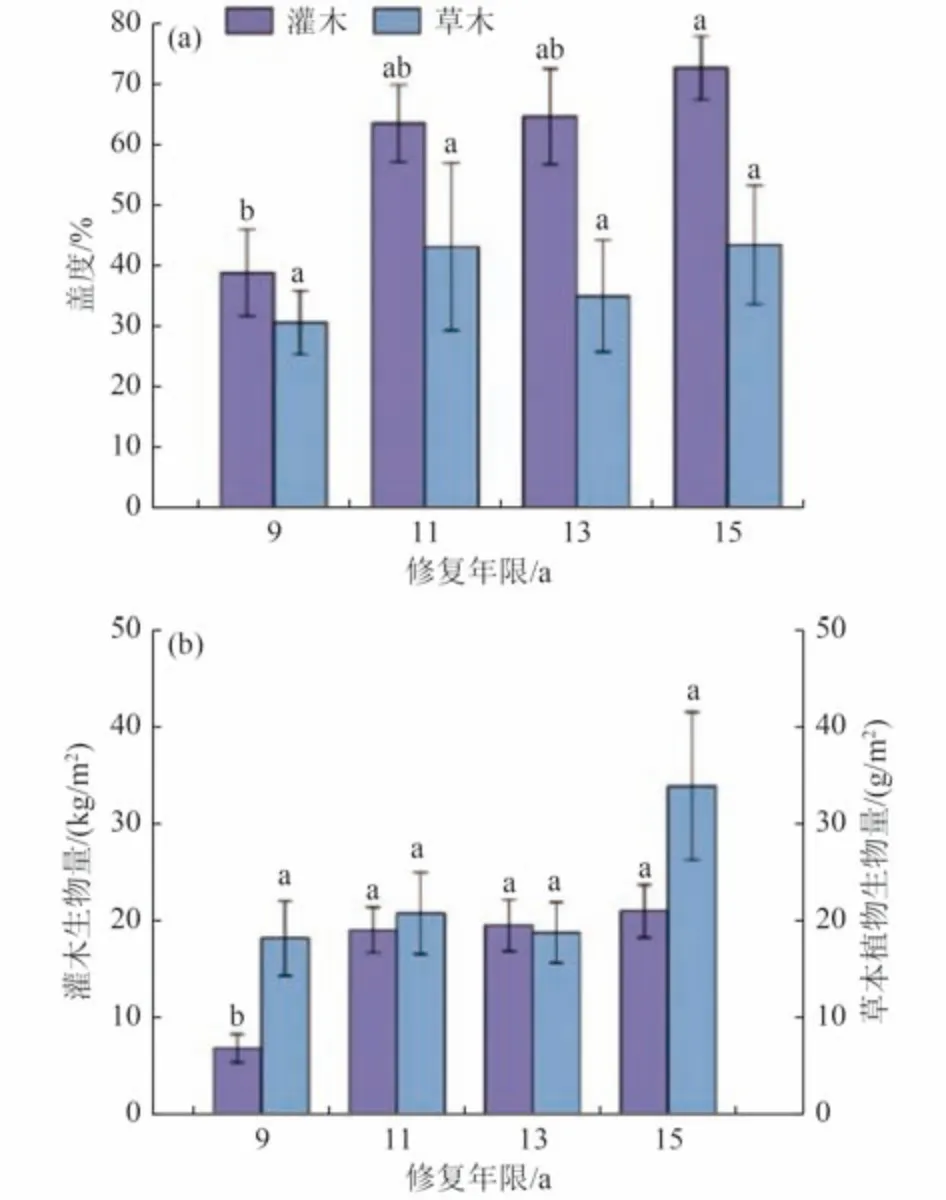
图4 甘珠尔沙地不同修复年限植被盖度、地上生物量比较Fig.4 Comparison of vegetation coverage and aboveground biomass in different restoration years in Ganzhuer sandy land
灌木总盖度随着修复年限的增加而增加,在修复年限最短的9 a(38.7%)和最长的15 a(72.6%)之间存在显著性差异(P<0.05),在其他修复年限之间差异不显著〔图4(a)〕。灌木地上生物量在修复9 a 与其他修复年限(11、13、15 a)之间均存在显著性差异(P<0.05),而11、13、15 a 之间差异不显著(P>0.05)〔图4(b)〕。总体来看,灌木地上生物量随修复年限的增加呈现持续增加的趋势。9 a 为最小值(6.75 kg/m2),11、13 、15 a 相比于9 a 分别显著增加了1.81 倍、1.88 倍和2.10 倍(P<0.05),15 a 为最大值20.9 kg/m2〔图4(b)〕。
2.3 物种多样性变化
不同修复年限的沙地草本植物多样性指数无显著差异(P>0.05)(图5)。草本植物的4 个多样性指数均在11 a 为最小值。Shannon-Wiener指数在9 a 为1.11,随后在11 a 降至最小(0.570)。与9 a 相比,13、15 a 分别高出0.14% 和0.710%〔图5(a)〕。Simpson指数在9 a 时最大(0.570),在11 a 降为最小(0.300),13 a 与15 a 基本稳定在0.570 左右〔图5(b)〕。Margalef 指数在9 a时为0.910,在11 a 时降至最小(0.500),13 a 时升至最大(0.930),15 a 时为0.860,比13 a降低了8.27%〔图5(c)〕。Pielou指数与Margalef指数的变化趋势相似,在9 a 时为0.710,11 a 时降至最小(0.440),13 a 时升至最大(0.760),15 a 为0.690,比13 a 降低了9.89%〔图5(d)〕。

图5 甘珠尔沙地不同修复年限植被多样性指数比较Fig.5 Comparison of diversity index of different restoration years in Ganzhuer sandy land
灌木层的多样性指数随修复年限的增加表现出的差异不同(图5),4 个多样性指数在9 a 与11、15 a均呈现出显著差异(P<0.05);而13 a 与其他修复年限差异均不显著(P>0.05)。整体上看,各多样性指数变化趋势一致,即上升—下降—上升。Shannon-Wiener指数在9 a 为最小(0.130),11 a 显著增加了3.34 倍后达到最大(0.560)(P<0.05)。而13 a 降至0.320,在15 a 有所回升(0.520)。修复年限最长(15 a)与修复年限最短(9 a)相比,显著提高了3.03 倍(P<0.05)〔图5(a)〕。Simpson 指数9 a 为最小(0.080 0),15 a 达到峰值0.360 且显著高于9 a(P<0.05)〔图5(b)〕。Margalef 指数在9 a 为最小(0.100),显著低于11 a 的最大值0.450(P< 0.05)。11、13、15 a 与9 a 相比,均提高了1.24 倍以上〔图5(c)〕。Pielou 指数在9 a 为最小(0.190),但随着修复年限的增加出现明显升高,11 和15 a 分别提高了2.44 倍和3.03 倍,差异达到显著水平(P<0.05)。13 a 与9 a 相比提高了1.48 倍,但差异不显著(P>0.05)〔图5(d)〕。
2.4 草本多样性与环境因子的关系
为直观表达环境因子与植被修复之间的相互关系,找出沙地生态系统修复过程中影响植物多样性变化的潜在驱动因素,并量化环境因子对植被多样性的贡献程度。本研究选用随机森林模型(RF)方法,将选取的环境因子土壤酸碱度(pH)、电导(EC)、含水量(SMC)、有机质含量(SOC)、总氮含量(TN)、总磷含量(TP)、总钾含量(TK)、年降水量(AP)、年蒸散量(AE)、年均温度(AAT)、年均风速(MAWS)、容重(UW))作为解释变量,草本植物多样性指数作为响应变量,进行筛选及模型回归。分析确定包含土壤养分因子与环境因子在内的相对重要性排序图(图6)。随机森林模型的结果显示,并非所有环境因子对多样性指数做出了相似的贡献。其中土壤总氮含量、总钾含量、有机质含量、年降水量和年均风速为影响沙地植被Shannon-Wiener 和Simpson 指数的关键性因子(P<0.01) 〔图6(a)、(b)〕。同时对于Shannon-Wiener 指数来说,pH 与土壤含水量也对其具有较高的解释度(P<0.01) 〔图6(b)〕。土壤总氮含量、有机质含量、土壤含水量和年降水量依次为显著影响Margalef 指数的环境因子(P<0.01)〔图6(c)〕。尽管环境因子对Pielou 指数影响均不存在显著性〔图6(d)〕,但总钾含量、土壤含水量、有机质含量与总氮含量仍为影响Pielou 指数的重要指标。部分依赖图(图7)结果表明,土壤总氮含量、总钾含量、有机质含量,土壤含水量及年降水量等环境因子与植被多样性指数呈显著正相关,且随着环境因子的提高而逐渐增加,后稳定在较高的水平。年均风速与植物多样性呈显著负相关,当风速超过2.27 m/s 时,沙地生境Shannon-Wiener 和Simpson 指数将会出现显著下降。

图6 环境因子与草本植物多样性指数随机森林模型重要性排序Fig.6 Importance ranking chart of random forest model of environmental factors and herb diversity index

图7 环境因子与草本植物多样性指数部分依赖图Fig.7 Partial dependence of environmental factors on herb diversity index
3 讨论
3.1 植物群落特征及优势度变化
群落的结构特征是研究生态系统的动态变化规律及其维持机制的基础。群落物种组成对群落动态变化规律以及植被恢复策略等方面的研究具有重要作用[25]。本研究中,不同修复年限的沙地植物群落的组成存在一定差异,草本植物种数呈现先减少后增加的趋势,这与一些学者在修复迹地的研究结果相似[26-27]。研究发现,草本群落主要集中在菊科和禾本两大科,其中对草地沙漠化环境具有一定忍耐性的菊科植物物种数最大,这与草地植被退化的逆向演替趋势恰恰相反[28]。以披碱草、狗尾草、糙隐子草和羊草为主的禾本科植物的数量较稳定,有较高的优势度,且随着修复年限的增加逐渐突显其在群落中的作用和功能。从生活型组成来看,在各修复年限的草本层组成中多年生草本占优势,且随着植被修复的进行,抗逆性强的多年生植物种类增加,并且优势度明显超过一年生及一/二年生植物,对群落生态功能的维持起主要作用。灌木群落中的优势科豆科的优势度及多年生草本优势度随着修复过程逐渐上升,这与张继义等[29]的研究结果相同。多年生植物具有比一年生植物更强的抵抗环境扰动和保持种群稳定的能力,因此物种生活型组成上的这种变化也反映了植被修复过程生态系统结构与功能的变化特征,群落结构趋于稳定,生态功能增强。群落总盖度和生物量从侧面反映了植物光合能力强弱及繁茂程度,是草地植物重要的数量特征。植被群落总盖度的增加能够显著减少水土流失和改善地表小气候,是植被修复的关键性指标。赵啸龙等[30]在科尔沁沙地自然修复过程中发现修复20 a 的草本植物群落总盖度和生物量显著大于修复12 与17 a。而本研究并未发现9~15 a 草本植物群落总盖度和生物量存在显著差异,与赵啸龙等[30]的研究结果并不一致,推测与修复地位置、修复年限、修复措施以及灌草植物的相互作用有关。
本研究发现,在9 a 时灌木群落保持着较低的群落总盖度与生物量,根据实地调查也发现该年限样地中灌木植株相较其他年限的灌木更矮小,根系相对不发达,所以根际对养分的截留能力较弱。根际是土壤水分和矿物质进入根系参与生物循环的门户[31],在半干旱荒漠区,土壤养分含量较低,因此根际微域内的有效养分决定着植物养分实际吸收的养分量,其截留效应的影响相比于农田和森林更明显[32]。由于受到养分可获得性的影响,该年限样地中草本与灌木植物的群落总盖度和生物量均处于较低水平。修复11 与9 a 相比,草本植物物种数出现降低,为恢复年限中的最小值。同年的样地中盐蒿物种成为最优势种,灌木总盖度大幅度提高,生物量也出现显著增加。研究表明,盐蒿同小叶锦鸡儿相比,具有更强的抗旱能力及适应性[33],因此盐蒿物种的优势度变化也反映了该修复阶段可能受到极端天气的影响使土壤中养分含量波动剧烈,使竞争能力较强的盐蒿物种脱颖而出。13 a 样地中灌木群落维持着较高的群落总盖度与生物量,且保持着持续稳定增长的趋势,其灌丛下分布的草本植物丰富度也有显著增加。较多的草本植物的出现可能与土壤中的养分富集有关。灌木群落在13 a 呈现出以豆科的小叶锦鸡儿为优势种,该旱生灌木的根瘤固氮作用可为植株生长提供氮元素,在缺水、贫瘠、没有人工干预的沙化草地上具有强烈的适应性[34],因此更多草本植物出现、定居。本研究发现15 a 样地中无盐蒿物种分布,调查时仅有小叶锦鸡儿与羊柴2 种豆科植物。考虑到盐蒿是一种好氧植物,适宜在修复年限早期较疏松的半固定沙地上生长,优势的盐蒿种群很快使土壤坚实化并向淡栗钙土化发展,使土壤通气性降低,造成生境条件转变从而抑制了自身生长[35]。同年的样地中由于缺少强抗旱的盐蒿作为竞争对手,小叶锦鸡儿与羊柴灌木保持较高的群落总盖度和地上生物量且随修复年限表现出显著性上升,其生态优势度逐渐突显,且朝着高大植物的趋势发展。在整个修复年限中,物种数、群落总盖度与地上生物量表现出波动增加,这与其他地区植被修复过程中变化规律一致[36-37]。
3.2 物种多样性变化
植物多样性是维持草地生态系统发展和生产力的最关键因素之一[38]。时间梯度上植物群落物种多样性的变化,在一定程度上可以反映出植被的演替变化[39]。本研究中,草本植物各项多样性指数随着修复时间的增加整体表现出增加的趋势,具体为先下降后上升的不显著波动。这种变化可以一定程度上反映出草本植物群落逐渐达到一个相对稳定的阶段,物种丰富程度和复杂程度相似,沙地生态系统保持着稳定的生态效应。草本植物Shannon-Wiener 指数、Simpson 指数、Margalef 指数与Pielou 指数在11 a均低于其他修复年限,反映出11 a 草本植物群落中物种数目少,群落中各物种分布较均匀,最优势种地梢瓜与披碱草的优势地位不突出,推测可能由于灌木群落在11 a 表现出较高的优势度与丰富度,其复杂的群落结构导致其下草本群落结构倾向于简单化发展。随着灌木群落郁闭度的持续增加以及其根系越来越发达,所吸收的养分随之增加,草本植物物种受到光资源与灌木竞争资源空间能力的逐渐减弱,同时优势灌木物种占据的生态位也对下层草本植物的生长起到一定限制作用[40]。本研究中15 a 草本植物与灌木植物的丰富度与多样性指数与9 a 相比有显著提高,15 a 样地中灌木的均匀度较高,这与单桂莲等[37]认为14 a 是较适宜的封育年限的结果有所不同,可能与不同物种对沙化生态系统的适应能力及程度的差别有关。盐蒿物种的退出使得沙地仅存在小叶锦鸡儿和羊柴2 种固沙能力强的灌木,由于竞争作用让二者及对土壤水分、养分的吸收处于相对较稳定的状态。
3.3 物种多样性与环境因子的关系
草本植物相较于灌木植物更新速度较快,对环境变化的响应更敏感。其多样性是生物与环境因子长期作用和适应的结果[41],体现了生物资源的丰富性与环境之间的复杂关系[42-43]。自然因素对沙化土地变化的影响是通过土壤、植被、降水、温度、风速等多方面共同造成的,而植物多样性[44]和植物物种分布[45]与气候、土壤养分密切相关,直接影响着生物多样性格局[46]。土壤是植物生长发育的基质[47],沙地土壤环境的改善直接影响植被演替进程[48-49],其养分的积累是影响植被盖度、植物群落组成和植物多样性的重要因素[50-56]。喀斯特地区[57]、黄山丘陵[58]和毛乌素沙地[59]等土壤养分匮乏的地区,全氮、有机质等土壤养分成为群落分布的决定性因子;在干旱地区水分也是植被恢复的限制因子,土壤含水量为调控森林-草原连续体及冻原生态系统的物种多样性的主要因子[60-61];pH 是湿地和半干旱草原植被分布的调控因子[62-63];全氮、pH 和全钾是绿洲植被结构的主要影响因子[64];有机质、全氮、土壤质地和湿度是荒漠生态系统植被分布的制约因子[65];全盐量、Cl-、K+、Na+、Mg2+等的含量是绿洲荒漠过渡带群落多样性的主要影响因子[66]。此外,对于同一的生态系统不同研究区而言影响植被群落分布与多样性的影响因子也有所不同。
通过对影响沙地修复区草本植物多样性的关键因子的研究,发现人工植被修复9~15 a 的沙地生态系统中,土壤总氮含量、总钾含量、有机质含量、年降水量、土壤含水量及年均风速等环境因子与植物多样性显著相关,其中土壤养分(如土壤总氮含量、总钾含量和有机质含量)与该地区的物种多样性显著正相关,这一结果与以往研究中发现由于土壤养分含量低导致植物物种多样性下降的结果相似[67-68],另外推测与半干旱地区典型的灌丛“肥岛”现象有关[69-71],灌丛肥岛的发育有利于根系养分的利用,产生较多的根际沉积,而根系的活动又会促进肥岛的发育[32],进一步加剧土壤养分胁迫。除土壤养分的影响,本研究还表明风速和降水成为制约植物的主要因素,其年际间的变化对物种多样性也有显著影响。对于荒漠程度下的沙化草地,年均风速的增加与植物多样性的显著负相关关系,表明更高的风速带来土壤表面更大的风蚀,使植物多样性降低。这与基于全球环境变化的长期监测研究发现风速、温度与降水影响着多年生物种多样性变化的结论一致[72]。与土壤养分因子相比,本研究中甘珠尔修复区的植被受到水分的影响表现出弱于土壤总氮含量、总钾含量和有机质含量等因子。推测其原因为研究区曾分布有旧河道,因此地下水对土壤仍存在一定的补给作用。另外,在沙质草地多为草本植物,密布的根系层对降水存在滞留作用[73],草本植物相比于灌木耗水相对较少,且较高的植被盖度产生的地表遮荫效果也能有效减弱土壤蒸发[74],这些因素的综合作用使其土壤水分含量提高,进而使得水分对植物多样性的限制作用弱于土壤养分。同时本研究发现,在本研究区中沙地土壤含水量对植被生长存在重要影响,这是因为在固定沙丘时的优势物种主要为根系较深、耗水强烈的盐蒿、小叶锦鸡儿与杨柴3 种灌木,使得土壤水分消耗强烈,因而导致草本植物受到一定程度的土壤水分的限制。综上所述,甘珠尔沙地修复区植被多样性受到气候、水分、土壤养分特征等因素的共同影响,因此需要对不同修复阶段的沙化草地植被群落进行调查与研究,并根据养分需求对其实施合理的修复措施。
4 结论
(1)甘珠尔沙地调查共发现植物42 种,以菊科、禾本科、蔷薇科、豆科为主。草本群落在不同修复年限样地中表现为菊科及禾本植物数量稳定且占有高优势度,群落总盖度和生物量变化不显著,但随修复年限的增加呈现先增后减再增加的趋势。物种分布均匀性良好,丰富度指数和多样性指数在各修复年限间出现小幅波动,但无显著差异,年际间变化稳定。灌木群落整体呈现出以豆科为优势种,随修复年限保持着较高的群落总盖度和地上生物量。灌木物种组成较单一,但多样性指数整体上随着修复年限的延长存在显著增加的趋势。
(2)甘珠尔沙地修复区草本植物多样性主要受土壤养分含量(土壤总氮、总钾、有机质)和修复当年的自然条件(年降水量、年均风速)影响。在沙化草地植被修复过程中,适当补充土壤养分含量(土壤总氮、总钾、有机质)和水分,有助于干旱区域荒漠草原生态系统的群落结构的重建和生物多样性的恢复。
(3)未来应持续开展沙地修复后的植被生态调查,掌握修复后生态系统群落结构及生物多样性情况,分析生态系统修复的影响因素,以了解沙化地区植被修复成效,为沙化区域植被修复策略的制定提供科学参考。
猜你喜欢
军事文摘(2021年18期)2021-12-02 01:27:58
现代畜牧科技(2021年4期)2021-07-21 06:13:08
内蒙古林业(2021年6期)2021-06-26 10:42:32
幼儿智力世界(2021年4期)2021-05-17 17:10:29
科普童话·百科探秘(2018年7期)2018-09-13 11:12:08
乡村科技(2018年8期)2018-02-13 12:16:18
中成药(2017年6期)2017-06-13 07:30:35
诗潮(2017年2期)2017-03-16 10:42:30
湖南林业科技(2017年1期)2017-02-06 05:29:17
动物营养学报(2015年3期)2016-01-07 11:06:10
