
粘质沙雷氏菌全能核酸酶的研究进展
2023-07-28 06:50赵萍韩冬梅
北方牧业 2023年13期
赵萍,韩冬梅⋆
(河北工程大学生命科学与食品工程学院,河北邯郸 056000)
全能核酸酶又称非特异性核酸酶 (Nonspecific nuclease,NU),可降解几乎所有形式的DNA 和RNA (包括单链、双链、线状、环状、天然以及变性的核酸),生成3~5 个碱基长度的5’-单磷酸寡核苷酸。全能核酸酶来源广泛,在动物、植物、细菌、真菌和病毒均有发现,到目前为止,已鉴定出30 多种不同来源的全能核酸酶,其中来源于粘质沙雷氏菌(Serratia marcescens)的全能核酸酶活力最强(下文无特殊说明,全能核酸酶均指来源于粘质沙雷氏菌全能核酸酶),能够在非常广泛的条件下保持很高的稳定性和消化活力。全能核酸酶在兽医方面的应用主要是去除兽用疫苗、蛋白、多糖类生物制药生产过程中的核酸残留,降解动物性饲料中的病原体的核酸残留,维护养殖场的生物安全。
1 全能核酸酶结构
粘质沙雷氏菌的全能核酸酶有两种基因亚型,第一种亚型由245 个氨基酸组成,第二种亚型相对与第一个亚型其N-末端少个三个氨基酸Asp-Thr-Leu,其余氨基酸序列相同。这两种亚型生化性质几乎相同,只是在等电点处有微小的差别。全能核酸酶的一级结构、二级结构、三级结构和四级结构均已被证实,率先被解析的是2.1 欧分辨率蛋白晶体结构,随后1.7 欧分辩率和0.92 欧分辨率蛋白晶体结构依次被解析出来。全能核酸酶不同于其他核酸酶的结构,其精细的结构是由一个包含六个反平行β 链的中心层组成,在中心层的两侧有不同长度的螺旋侧翼,几个长的环段将这些区域连接起来。另外,沉降速度实验、交联研究、动态光散射实验以及X 射线晶体数据分析均证实全能核酸酶是由两条相同的单肽链连接在一起的同源二聚体。Ahrenholtz 研究发现粘质沙雷氏菌核酸酶单体的催化中心距离二聚体的接触位点较远,二聚体的分离并不影响活性位点与底物的结合。Franke 通过设计正常的核酸酶二聚体、核酸酶单体、以及含有一个突变失活亚基的核酸酶异二聚体三组实验测定酶活力,结果显示核酸酶单体、核酸酶异二聚体的活力为正常核酸酶二聚体的一半,从而进一步证明了沙雷菌核酸酶两个亚基的活性位点具有独立性。虽然全能核酸酶的二聚化结构不具有协同作用,但增加了酶与底物的碰撞几率,增强了反应效率。
2 全能核酸酶的表达调控
全能核酸酶的表达调控属于选择性表达调控而非常见的底物调控,启动子上游的操纵基因决定了全能核酸酶的分泌发生在细菌生长的平台期,胞外核酸酶的分泌量不随着细胞数量的增加而显著增加,但在细胞裂解后胞外核酸酶的量会显著增加。全能核酸酶的激活与位于多肽链的N 末端的C9 与C13 和C 末端的C201与C243 之间的两个二硫键的形成有关,二硫键的形成发生在细胞周质,因此全能核酸酶分泌到细胞周质或细胞外才具有生物学活性,从而避免了全能核酸酶对自身核酸的降解。
3 全能核酸酶作用机制
3.1 全能核酸酶活性中心功能基团
全能核酸酶催化功能相关的基团是位于活性中心的R57、R87、H89、N119 和E127 残基,对这些残基进行突变会导致该酶催化活性的丧失,其中H89 直接参与了磷酸二酯键的水解反应,R57和N119 残基共同起着稳定磷中间体的作用,E127 在整个过程中充当普通碱的作用,R87 作用位点在核糖3' 端的磷酸基上,这对5' 端附近的降解是至关重要的。
3.2 全能核酸酶底物的作用位点
与其他来源的核酸酶比较,全能核酸酶有较高的催化活性,大约是葡萄球菌核酸酶的4倍,是DNase Ⅰ的34 倍。其催化机理与其它核酸酶也略有不同,葡萄球菌核酸酶作用于5’-OH 与磷酸之间形成的磷酸二酯键,全能核酸酶和DNase I 作用位点相同,为3’-OH 与磷酸之间的磷酸二酯键,但全能核酸酶是从5’端切割磷酸二酯键,而DNase I 是从3’端切割磷酸二酯键。
3.3 全能核酸酶对底物的偏爱性
虽然全能核酸酶底物谱广泛,能够将单链、双链、线状、环状、天然以及变性的DNA 和RNA降解成3-5 个碱基长度的5’-单磷酸寡核苷酸。但对Poly 结构的核酸降解效率差异很大,对PolyU,PolyG , PolyC,Poly[d(A)]和Poly[d(T)]几乎不降解,对PolyA 和PolyA·PolyU 可微量降解,对Poly( I)·Poly( C) 可高效降解。另外,全能核酸酶对高GC 含量的碱基序列降解效率高于高AT 含量的碱基序列。
4 重组全能核酸酶的表达
鉴于全能核酸酶在胞内表达会对自身核酸造成降解,从而引起菌体死亡或抑制菌体生长。目前获得重组全能核酸酶以分泌性表达为主。默克公司最早利用大肠杆菌对重组全能核酸酶进行了分泌性表达,其产量仅达到104单位/升,远远不能满足生产需求。国内学者也陆续对全能核酸酶重组表达进行了研究,张开俊将全能核酸酶基因构建到分泌型载体pET-20b(+)上,在大肠杆菌BL21 进行了分泌表达,其产量为8毫克/升。陈鹏为了解决全能核酸酶胞内本底表达对宿主细胞的伤害,将全能核酸酶与麦芽糖结合蛋白融合基因构建到大肠杆菌表达载体PET22b 上,并在大肠杆菌BL21 中进行了分泌性表达,其产量达到了10 毫克/升。龚雪梅利用短短芽孢杆菌胞外分泌能力,将全能核酸酶基因重组到pNC 载体上,实现了全能核酸酶重组蛋白在短短芽孢杆菌胞外分泌表达,其产量达到30~40 毫克/升。田克恭利用酵母真核表达系统可实现外源蛋白的分泌表达优势,将全能核酸酶基因重组到pPICZɑA 载体上,实现了全能核酸酶重组蛋白在酵母胞外分泌表达,其产量达到200 毫克/升。
5 全能核酸酶的应用
由于全能核酸酶可在广泛的条件下能保持较高的稳定性和消化活力,高效地降解所有形式的RNA 和DNA,已广泛地应用于生物医药领域。全能核酸酶最典型的用途是除去疫苗、多糖、蛋白等工业生物制品中核酸,美国FDA 对治疗用每剂量的生物制品的核酸含量要求低于10 pg,国内相应的要求为低于100 pg。全能核酸酶可以使这些生物制品中的核酸含量符合要求,提高生物制品功效。其次,在蛋白纯化过程中,细胞裂解产生的大量核酸增加了裂解液黏度,使下游的生产程序复杂化,全能核酸酶通过降解核酸可有效降低细胞裂解液的黏度,从而提高蛋白得率,改善分离效果。另外,在细胞来源的产物纯化过程中,核酸易粘附在细胞产物上,使其颗粒大小或电荷发生改变,引发细胞产物聚集,如外周血单细胞(PBMC)存放过程中的结块现象,添加全能核酸酶可避免此类情况的发生,有利于提高细胞产物纯化效率。在ELISA、色谱分析、双相电泳和足迹分析等生化分析样品的制备过程中,用全能核酸酶处理含核酸的蛋白样品,可提高蛋白的分辨率和回收率。PCR 实验室核酸污染是造成检测结果假阳性的主要因素,全能核酸酶通过降解核酸实现实验室的净化。在医学方面,由于核酸酶具有干扰细胞复制的功能,具有明显的抗肿瘤特性,并且对纤维性囊肿、系统性红斑狼疮,儿童气喘病等疾病也有明显的疗效。
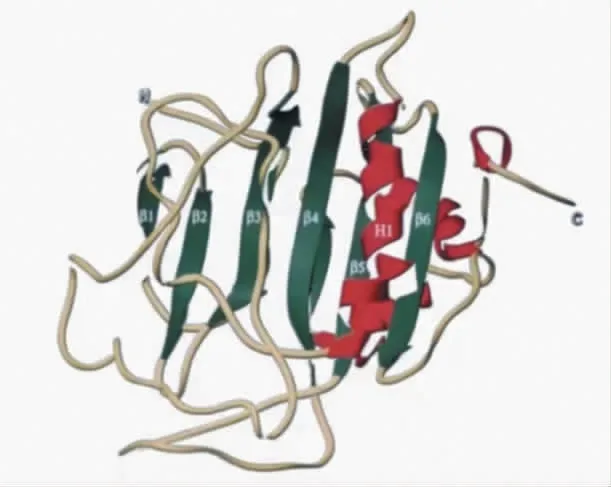
图1 全能核酸酶单聚体结构
6 小结与展望
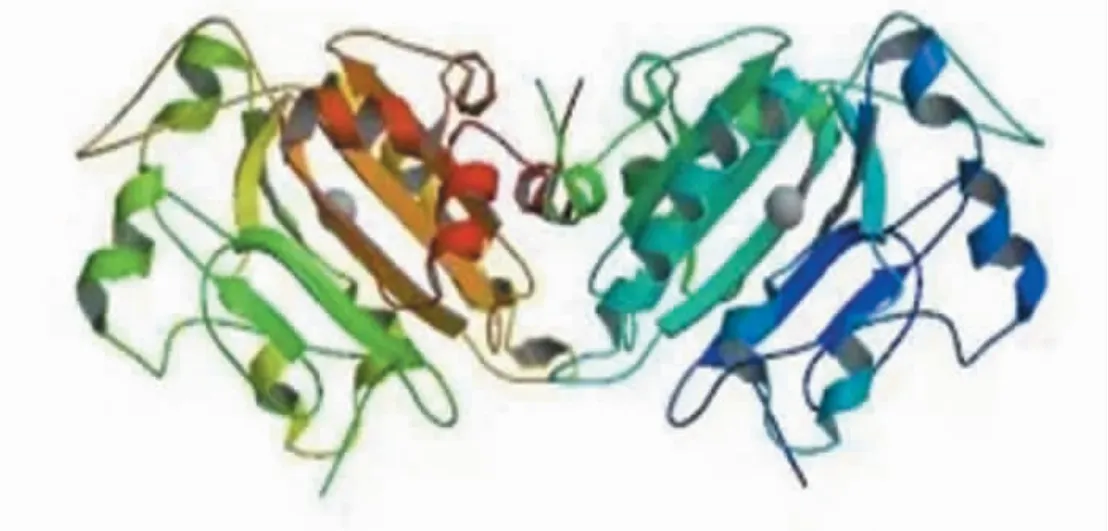
图2 全能核酸酶二聚体结构

图3 DNase I,葡萄球菌核酸酶以及SMNE 的切割位点
全能核酸酶应用范围广泛,作用效力强,不仅在工业、生物医药领域有突出表现,也可以在畜牧生产中派上用场,全能核酸酶可降解猪血浆蛋白粉携带的病原微生物核酸,尤其是非洲猪瘟病毒核酸,可提高猪血浆蛋白的饲用安全,降低动物疫病传播风险,维护猪血浆的畜牧生产价值。另外全能核酸酶与蛋白酶联合应用,有望成为新型的环保生物消毒剂,首先通过蛋白酶降解病原微生物的外膜蛋白使其核酸暴露,全能核酸酶进一步发挥降解核酸作用,从而达到消毒效果,与传统化学消毒药相比,对机体无毒副作用,不污染环境,是一种潜在的理想绿色消毒制剂。
猜你喜欢
幼儿画刊(2022年8期)2022-10-18
幼儿画刊(2022年10期)2022-10-14
幼儿画刊(2022年9期)2022-09-14
幼儿画刊(2022年5期)2022-06-06
环境污染与防治(2022年5期)2022-05-29
无机化学学报(2020年7期)2020-07-20
中国预防兽医学报(2020年2期)2020-06-01
三农资讯半月报(2020年8期)2020-05-13
食品与发酵工业(2018年11期)2018-12-15
生物学杂志(2017年6期)2017-12-19
