
有机废弃物对辣椒、番茄幼苗生长及根际微生物群落的影响
2023-07-17 07:51解志强赵强彪朱德颜毛振坤彭翠仙
热带农业科技 2023年3期
王 灿,解志强,孙 媛,赵强彪,朱德颜,毛振坤,彭翠仙*
(1.文山州农业科学院,云南文山 663000;2.文山州植保植检站,云南文山 663000)
蔬菜是云南高原特色农业重点产业。其中辣椒(Capsicum annuumL.)、番茄(Lycopersicon esculentumMill.)已成为云南省主要的外销亚热带果蔬作物之一。随着产业化、机械化、规模化、自动化程度的不断提高,对育苗基质的需求量也急剧上升[1]。目前市场上普遍使用的育苗基质主要成分是草炭,但其资源有限、短期内不可再生,大量开发会导致生态环境恶化,且国内草炭品质参差不齐,影响育苗效果。为此,利用一些农业有机废弃物,通过发酵腐熟、炭化、添加外源物等方式[2],开发价格低廉的新型有机复合育苗基质代替草炭已成为当务之急和研发的热点,具有重要的经济和生态意义。农业生产过程中产生的有机废弃物富含有机质、N、P、K 等营养元素[3],通过堆肥发酵、炭化处理、添加外源物料等方式制成育苗基质,一方面育苗效果显著,提高了农业废弃物利用率,另一方面保护了生态环境。Yousry 等[4]研究发现,葡萄果实废弃物堆肥发酵后与泥炭或蛭石1∶1 混合作为黄瓜育苗基质,可显著提高种子萌发、幼苗成活率及质量,同时降低育苗成本,可用于园艺苗圃的商业化生产。Perez-Murcia 等[5]利用城市污泥替代草炭作为基质观察对西兰花生长的影响,结果表明城市污泥用量为30%时西兰花生长最快。
辣椒、番茄主要采用育苗盘方式进行育苗,育苗基质是影响幼苗质量的主要因素,不仅为幼苗生长提供水分与养分,同时也起到对外界环境不利因素刺激的缓冲作用[6]。有研究表明,基质的理化性状会影响微生物类群、数量、养分释放量,进而影响植物对养分的利用情况[7-9]。王闯等[9]研究表明,椰糠+菇渣+珍珠岩制成育苗基质明显促进番茄植株生长,其产量、光合特性、根呼吸和微生物含量都显著高于对照(草炭)。宋佳泽[10]通过将不同处理的秸秆用于黄瓜种植,结果发现各处理均在一定程度上促进了根际土壤中酶活性和芽单胞菌科、鞘脂单胞菌科等土壤微生物生长繁殖,进而提高土壤养分,促进黄瓜生长发育。张凯歌等[11]发现,柠条堆肥结合适度深翻有效促进了土壤细菌群落的多样性,提高功能基因丰度与6 大碳源利用水平。
大量研究表明,有机废弃物基质对植物幼苗生长有显著促进作用,但对辣椒、番茄幼苗根际微生物群落结构的影响鲜有报道。因此,本研究采用16S/ITS rDNA 对成苗期的辣椒、番茄根际微生物进行扩增子测序,旨在探究有机废弃物基质对辣椒、番茄幼苗根际微生物群落结构的影响,为利用有机废弃物作为育苗基质提供理论依据。
1 材料和方法
1.1 供试材料
辣椒品种为‘京滇皱皮椒’,购买于云南京滇种业有限公司。番茄品种为‘305’,由云南农业大学滇台特色农业产业化工程研究中心提供。
有机废弃物育苗基质主要材料包括稻壳生物炭、发酵茶渣等,为上一阶段试验筛选得出的最佳配比[12]。商品育苗基质为‘绿成丰’牌,由连云港恒奥达肥料科技有限公司生产,主要成分为草炭土、蘑菇渣、珍珠岩、矿石、豆粕渣。基质理化性状由试验前期完成测定(表1、表2)。

表1 基质物理性状

表2 基质化学性状
1.2 方法
试验于2021 年4 月—2021 年12 月在文山州农业科学院进行,共设2 个处理,A 处理为有机废弃物育苗基质,CK 处理为商品育苗基质。
将发芽整齐一致的辣椒、番茄种子播种于每盘36 孔的育苗盘中,每盘为一次重复,每个处理各3 次重复。苗龄65 d 时,采用5 点取样法选取整齐一致的植株整株,拔出并抖落附着的土壤基质,用无菌刷刷取粘附在根表面的土壤基质混合于50 mL 无菌离心管中,迅速置于干冰中保存运输,后置于-80 ℃冰箱保存备用,同时收集抖落土壤用于检测理化性质;将植株保存于4 ℃用于生化检测。
植株干重用分析天平测量,株高和茎粗用刻度尺、游标卡尺测量。叶绿素采用紫外分光光度法测定[12],根系活力采用TTC 法测定[13],可溶性糖含量采用蒽酮比色法测定[14],蛋白质含量采用考马斯亮蓝G-520 染色法测定[15]。
生长函数(mg/d)=全株干重/苗龄
壮苗指数=(地下部分干重/地上部分干重+茎粗/株高)×全株干重
秧苗根际微生物DNA 提取均使用HiPure Soil DNA Kit 核酸提取试剂盒,使用Qubit® dsDNA HS Assay Kit 检测DNA 浓度。由GENEWIZ,Inc(South Plainfield, NJ)公司完成二代测序文库构建和测序。16S rDNA 上V3 及V4 区 引物 系列 如下:F:5’-CCTACGGRRBGCASCAGKVRVGAAT-3’;R:5’-GGACTACNVGGGTWTCTAATCC-3’。ITS rDNA 上的ITS2 可变区 引物如下:F:5’-GTGAATCATCGARTC-3’;R:5’- TCCTCCGCTTATTGAT-3’。通过Illumina MiSeq 进行双端测序。
1.3 数据统计与处理
数据差异显著性分析使用DPS 7.05(Duncan新复极差法,P<0.05)。测序数据质量优化使用Cutadapt 1.9.1、Vsearch 1.9.6、Qiime 1.9.1。
2 结果与分析
2.1 对幼苗生长代谢的影响
不同处理下辣椒、番茄的株高、茎粗无显著差异(表3),在干重、生长函数、壮苗指数方面,A 处理显著高于CK。在生化指标上(表4),辣椒中A处理叶绿素含量、根系活力显著高于CK,可溶性蛋白和可溶性糖含量则无显著差异;番茄中A 处理叶绿素、可溶性蛋白、可溶性糖含量、根系活力均显著高于CK。
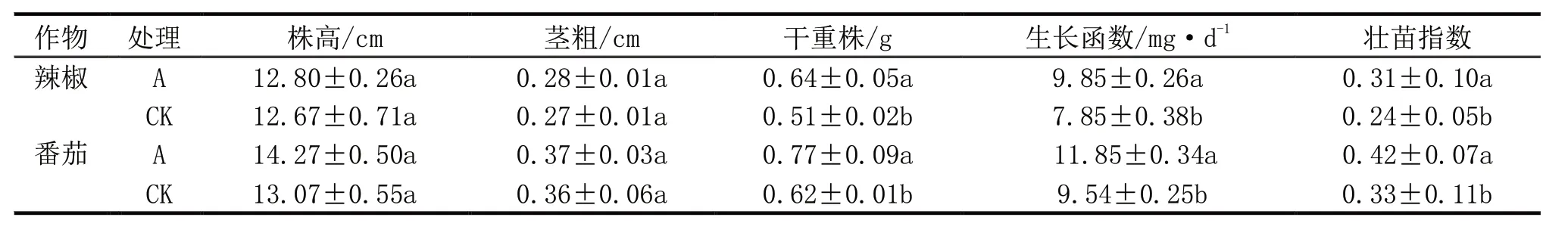
表3 不同处理对幼苗生长的影响

表4 不同处理对幼苗生化指标的影响
2.2 根际微生物群落组成
试验共收集4 个样本,质控后依据97 %相似性聚类获得269 个细菌OTUs、142 个真菌OTUs(不同样本取并集所得)。A 处理下辣椒、番茄根际土壤 细 菌OTU 个 数 分 别 为186、219,CK 则 分 别 为247、227(图1a)。A 处理辣椒、番茄根际土壤真菌OTU 个数分别为71、77,CK 则分别为97、126(图1b)。A 处理作物根际土壤细菌和真菌OTU 数均低于CK。

图1 根际微生物OTU花瓣图
土壤细菌门水平组成如图2a 所示,4 个土壤样本共得到9 个类群,相对丰度从大到小依次为变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)、蓝细菌门(Cyanobacteria)、Patescibacteria、酸杆菌门(Acidobacteria)、厚壁菌门(Firmicutes)、芽单胞菌门(Gemmatimonadetes)、绿弯菌门(Chloroflexi)。A 处理辣椒和番茄根际土壤中酸杆菌门占比分别是5.97%和8.27%,低于CK(10.48%、21.92%),芽单胞菌门、绿弯菌门、Patescibacteria、蓝细菌门占比也均低于CK。但A 处理下辣椒、番茄根际土壤中厚壁菌门分 别 为 2.21% 和 3.93% ,高 于 CK(1.24% 和0.59%)。土壤细菌属水平下(Top15),黄杆菌属(Flavobacterium)、假单胞菌属(Pseudomonas)、Devosia为主要细菌属,且A 处理以上细菌属相对丰度均高于CK(图2b)。真菌门水平下,除A 处理辣椒样本外,其余样本物种数目均为4,主要菌群为子囊菌门(Ascomycota)和担子菌门(Basidiomycota)。A 处理辣椒、番茄根际土壤中担子菌门相对丰度分别为26.43%和20.52%,高于CK(1.67%和1.61%,图2c)。属 水 平 下(Top15),毛 孢 子 菌(Trichosporon)、Aspergillus、Graphium为主要菌属,且A 处理以上真菌属相对丰度均高于CK(图2d)。
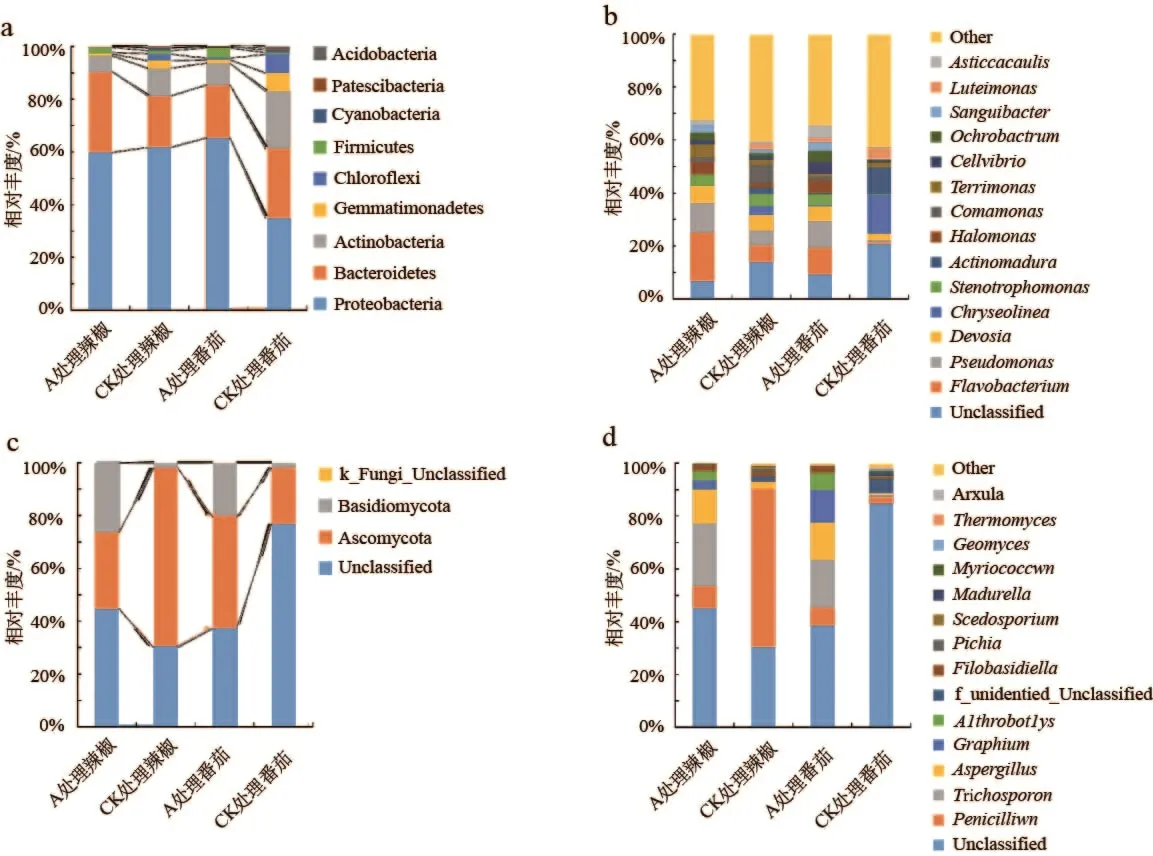
图2 根际土壤细菌、真菌门水平与属水平群落组成
2.3 根际微生物群落多样性分析
根据表5 可知,辣椒A 处理根际土壤细菌丰富度指数Ace 和Chaol(200.66 和197.04)显著低于CK(254.26 和253.38),多 样 性 指 数Shannon 和Simpson(5.03 和0.98)同 样 显 著 低 于CK(6.83 和1.10);真菌情况与细菌相似,A 处理根际土壤真菌丰富度指数和多样性指数均显著低于CK 处理。A 处理番茄根际土壤细菌丰富度指数和多样性指数均低于CK,但两者间无显著差异;真菌丰富度指数和多样性指数则显著低于CK。表明A处理显著降低了辣椒根际土壤细菌、真菌群落的丰富度和多样性,对番茄根际细菌群落多样性无显著影响,但显著影响真菌群落多样性。
主坐标分析中,样本点距离的远近代表了样本中微生物群落的相似性,距离越近,相似度越高;聚集在一起的样本由相似的微生物群落构成。如图3a,在细菌中第一主坐标揭示了样本间72.26%的差异并将A 处理和CK 处理区分开,第二主坐标揭示了样本18.12 %的差异。真菌中(图3b)第一主坐标揭示了样本间61.5%的差异并将A处理和CK 处理区分开,第二主坐标揭示了样本35.33%的差异。

图3 根际微生物PCoA分析
2.4 微生物群落与基质性状相关性分析
冗余分析中选择基质土壤相关理化性状与属水平下丰度前10 的微生物进行分析。根据图4a,持水孔隙度、通气孔隙度、速效P、速效K、pH 及腐殖质均对A 处理辣椒、番茄的根际细菌群落结构产生显著影响,是导致细菌群落组成改变的影响因子。而速效P、速效K、pH 及有机质对A 处理真菌群落结构组成有显著影响(图4b)。

图4 细菌(a)、真菌(b)与土壤理化性质的冗余分析
土壤细菌群落与理化性质的Spearman 分析中(图5a),A 处理优势菌厚壁菌门相对丰度与pH值、有机质、速效K 含量显著正相关,与速效N 含量显著负相关(P<0.05)。而CK 处理中的优势菌酸杆菌门、芽单胞菌门、绿弯菌门则正好相反。酸杆菌门相对丰度与pH 值、有机质、速效K 含量显著负相关;芽单胞菌门相对丰度与pH 值、有机质、通气孔隙度显著负相关,与速效N 含量显著正相关;绿弯菌门丰度与pH 值、有机质显著负相关,与EC 值、速效N 显著正相关(P<0.05)。在土壤真菌与理化性质的相关性分析中(图5b),A 处理的优势菌担子菌门相对丰度与pH 值、通气孔隙度、有机质、速效K 含量显著正相关,与EC 值、速效N 含量显著负相关(P<0.05)。

图5 细菌(a)、真菌(b)与土壤理化性质的Spearman分析
3 讨论
本试验发现,与传统的商业育苗基质相比,有机废弃物基质处理并未降低辣椒和番茄的生长质量,反而提高了叶绿素含量、根系活力等指标。表明有机废弃物基质具有与商品基质同样育苗效果的潜力,这与前人研究结果一致[16]。通过16S/ITS rDNA 扩增子测序进一步对两者间根际微生物群落结构进行分析,发现A 处理辣椒、番茄根际土壤细菌和真菌的OTU 数均低于CK,表明A 处理降低了根际微生物的OTU 个数。在A 处理下辣椒和番茄根际土壤中酸杆菌门、芽单胞菌门、绿弯菌门、Patescibacteria 和蓝细菌门的相对丰度均低于CK,但厚壁菌门的相对丰度则高于CK。真菌中主要菌群为子囊菌门和担子菌门,且A 处理辣椒、番茄根际土壤中担子菌门相对丰度显著高于CK。表明有机废弃物基质处理改变了根际微生物群落结构,使辣椒和番茄根际中厚壁菌门、黄杆菌属、假单胞菌属和Devosia、毛孢子菌、Aspergillus、Graphium等显著富集。这可能是废弃物基质中添加有生物炭,而研究表明生物炭可直接(提供易分解的有机碳和营养元素)或间接(改变土壤基质物理性质增加孔隙度等)影响微生物群落[17-18],同时基质中的茶渣含有丰富的有机质和腐殖质,也为微生物的繁殖提供了营养物质。厚壁菌门中的许多成员都是植物益生菌,具有固氮和纤维素降解作用,能够分解相关营养物质供作物生长利用,从而促进作物生长[20]。
α 多样性指数中,Ace 和Chao1 反映菌群丰富度,数值越小表明丰富度越低,Shannon、Simpson反映菌群多样性,其数值越小表明群落多样性越低。A 处理显著降低了辣椒根际细菌、真菌多样性与丰富度,显著降低番茄根际真菌群落多样性,对番茄根际细菌丰富度无显著影响。废弃物基质中添加了适量壳聚糖,研究表明壳聚糖可显著降低环境中微生物群落丰富度及多样性[19]。董春娟等[19]分析认为,壳聚糖可促进某些可分解利用壳聚糖的微生物繁衍,抑制其它微生物生长,导致相关微生物富集。主坐标分析表明,不同处理下的辣椒、番茄根际微生物群落结构差异较大,A 处理下辣椒和番茄幼苗间根际细菌、真菌群落结构差异较小,CK 处理下辣椒、番茄细菌、真菌群落结构差异均较大。表明农业废弃物基质定向改变了辣椒、番茄根际土壤细菌、真菌群落结构。
冗余分析和Spearman 分析表明有机质含量、孔隙度等是改变微生物群落丰度的主要因素。土壤基质不仅为幼苗生长提供水分与养分,同时也为微生物生长与繁殖提供适宜的环境,其物理化学性质影响了基质中微生物群落组成[21]。进一步说明了农业废弃物有机质含量、孔隙度等对根际微生物群落结构有一定影响。向羽等[22]研究表明,向种植土壤中添加腐熟菜籽饼肥可显著促进烟株生长,提高烤烟根际土壤微生物对氨基酸、聚类化合物碳源的利用能力,其根际微生物代谢功能及多样性指数显著高于对照。农业有机废弃物中含有丰富的营养物质,如有机酸,氨基酸、糖类等其它的大分子有机化合物,对土壤微生物群落组成、功能和碳源利用效率均有显著影响[23-25],同时有机废弃物增加了土壤孔隙度,为好氧微生物提高了良好生存环境。Dombrowski等[26]研究多年生黄花亭荠在自然和控制条件下根际细菌群落组成,发现土壤性质能够解释根内细菌15%的群落变异性,而环境条件和植物基因型最多解释了11%的群落变异性,且土壤类型对根内细菌门分类水平上的影响更加显著。因此,可以通过调控作物根际生长环境,定向改变其根际微生物群落,抑制某些有害微生物,让相关有益微生物富集,从而促进作物生长。
4 结论
本研究结果表明,在相同基质土壤环境中,同科不同属(辣椒属、番茄属)的作物幼苗根际微生物群落结构存在一定差异,但有机废弃物基质可通过改变环境因子如增加适量有机质含量和孔隙度等来调控它们之间的差异,达到定向改变微生物群落,促进有益微生物富集,进而协助作物生长或抑制相关病原微生物。这为有机废弃物作为育苗基质的应用提供了新的见解。
猜你喜欢
英语文摘(2022年6期)2022-07-23
中国土壤与肥料(2021年5期)2021-12-02
军民两用技术与产品(2021年10期)2021-03-16
世界博览(2020年19期)2020-10-30
中国比较医学杂志(2020年4期)2020-05-26
水生生物学报(2019年4期)2019-07-20
生物安全学报(2019年3期)2019-02-15
川北医学院学报(2019年6期)2019-02-10
天然产物研究与开发(2018年3期)2018-05-07
山西教育·幼教(2017年2期)2017-04-23
