
喀斯特次生林优势种群结构及数量动态1)
2023-07-10 07:54:24韦红艳徐铭柴宗政孙彩丽
东北林业大学学报 2023年7期
韦红艳 徐铭 柴宗政 孙彩丽
(贵州大学,贵阳,550025) (贵州民族大学)
植物种群结构及数量动态是植物生物学特性与环境相互影响、相互适应的结果[1-3]。研究森林群落内不同优势种群结构及数量动态,可以有效反映种群间的相互作用及种群与环境之间的联系,对揭示种群数量特征及变化规律,探究其所在群落的演替趋势、物种多样性的建成及维持至关重要[4-6]。次生林是原始森林生态系统中的一种退化类型,是指由于受人为破坏性干扰或异常自然干扰,使原始林固有林分结构、物种组成或基本功能发生显著变化后,经过天然更新或人工诱导而恢复形成的林分[7-8]。由于次生演替具有演替进程不一、群落结构多样的特点,因此次生林的发展具有双向性,若继续受到破坏,则将朝着退化的趋势发展,若能根据其结构特点采取合理经营措施,则能迅速恢复正向演替[8]。因而,科学合理保护及恢复次生林对其稳定持续发展具有重要意义。
中国喀斯特地貌区面积约占国土总面积的36%,其中以贵州省为中心的西南地区是世界上三大喀斯特地貌的典型区域之一[9-10]。特殊的地貌结构和强烈的岩溶导致喀斯特区域岩石裸露率高、土壤贫瘠且不连续、植物体发育迟缓、群落结构简单,不合理的人为干扰则造成现存森林多以片段化分布的次生林为主,次生演替进程缓慢[11-13]。因此,如何促进喀斯特次生林正向演替及实现稳定快速发展是喀斯特区域植被恢复和生态建设中优先考虑的问题[8]。目前喀斯特植物种群结构及数量动态研究大多集中于濒危或珍稀植物且多为单优种群,如对广东松(Pinuskwangtungensis)[14]、南方铁杉(Tsugatchekiangensis)[14]、掌叶木(Hadeliodendronbodinievri)[15]、单性木兰(Kmeriaseptentrionalis)[16]、黄枝油杉(Keteleeriacalcarea)[17]等的研究,对于多树种森林群落中不同优势种群结构及数量动态对比的研究较少,尤其是对喀斯特次生林这一特殊森林类型的研究尤为匮乏。
由于喀斯特次生林群落结构的不稳定性和演替方向的不确定性,对其优势种群结构及动态的研究能够有效反映各种群与环境之间的相互关系,对分析次生林群落现实状况及未来发展态势、制订合理的保护及恢复措施具有重要意义。为此,本研究实地踏查后选取贵州省喀斯特典型地区撂荒30余年后天然恢复的次生林,通过典型样地布设和系统植被调查,从龄级结构、数量动态、时间序列预测等方面,对比分析5个优势种群的结构及数量动态特征,以期为喀斯特次生林保护与恢复提供参考。
1 研究区概况
研究区位于贵州省喀斯特地区——紫云苗族布依族自治县(25°21′43″~26°2′30″N,105°55′14″~106°29′56″E),处于贵州高原和广西丘陵之间过渡的天然边界区域。平均海拔1 185 m,年均降雨量1 337.1 mm,年均气温15.3 ℃,属于亚热带季风性湿润气候,气候垂直变化及季风交替明显,太阳辐射能量低,无霜期长。紫云县境内的格凸河国家级风景名胜区中洞景区喀斯特地貌类型丰富,石漠化严重,土壤类型属于非地带性石灰土,土壤贫瘠,山体母岩主要为沉积岩与碳酸盐岩。研究地点在上世纪80年代中期前为农耕地,主要种植作物为玉米,弃耕后开始形成大面积的撂荒区域,现已有30多年,人为干扰较少,群落自然恢复状况良好[18]。
2 研究方法
2.1 样地设置与种群调查
在对研究区多次踏查的基础上,选取紫云县境内具有典型喀斯特地貌的撂荒地次生林地段作为样地,以热带森林中心(CTFS)样地建设方法为依据,设置面积为16 800 m2的标准矩形样地(140 m×120 m),将样地划分成42个样方,每个样方长、宽为均为20 m。以西南角为起点,对所有样方内胸径(D)>0的木本植物进行每木检尺,并记录林木物种名、胸径、树高、冠幅、枝下高、生长状态等信息[18],同时,使用GPS对样地进行定位,记录海拔、经纬度等地理信息,调查记录坡度、坡向、坡位、郁闭度等生境内容。
2.2 龄级划分
在生境条件相似的情况下,环境对林木径级、龄级的影响具有相对一致性。由于林业上在不破坏树木情况时很难获取种群具体树龄,故大多研究采用径级代替龄级[5,19-20]。本研究中,5个优势种群龄(径)级划分结果如下:Ⅰ龄级(D<1 cm)、Ⅱ龄级(1 cm≤D<3 cm)、Ⅲ龄级(3 cm≤D<5 cm)、Ⅳ龄级(5 cm≤D<7 cm)、…、Ⅻ龄级(D≥21 cm)。基于样地调查数据分析,将光皮桦(Betulaluminifera)、枫香(Liquidambarformosana)各划分为12个龄级;马尾松(Pinusmassoniana)、化香(Platycaryastrobilacea)均划分为11个龄级;山杨(Populusdavidiana)划为10个龄级。
2.3 种群数量变化动态指数


2.4 种群时间序列模型预测
根据时间序列分析中一次平均推移模型,分别对马尾松、枫香、山杨、化香、光皮桦等5个优势种群年龄结构进行动态模拟和推测[22]。公式如下:
式中:Mt为种群植株存活数量;n为需要预测的时间;t为龄级;Xk为第k龄级中的植株存活数量。
2.5 数据处理
采用R语言进行数据分析和处理。其中,种群数量变化动态指数和时间序列模型采用自编函数计算,图形采用ggplot2程序包绘制。
3 结果与分析
3.1 次生林树种组成及优势种群确定
本研究中,调查木本植物共计41种,3 251株(表1),其中,幼苗446株,占总数量的13.7%;幼树1 291株,占总数量的39.7%;成年树1 514株,占总数量的46.5%。调查木本植物中,光皮桦1 038株,占总数量的31.9%;化香793株,占总数量的24.4%;马尾松344株,占总数量的10.6%;枫香198株,占总数量的6.10%;山杨183株,占总数量的5.60%。5个树种共计2 556株,占总调查植物总数的78.60%,为群落中的优势种群。从不同生长阶段来看,5个优势种群中幼苗101株(22.6%)、幼树1 081株(83.7%)、成树1 374株(90.7%)。随着林分发展,优势种群的株数占比逐渐增大,优势度越明显。总体来看,该次生林以光皮桦、化香、马尾松、枫香、山杨为优势种群。

表1 次生林树种组成及数量特征
3.2 优势种群龄级结构特征
优势种群龄级结构如表2显示。5个优势种群龄级结构相对完整,且各种群个体数量随龄级的增大总体呈先增加后降低的单峰变化趋势。5个优势种群Ⅲ~Ⅴ龄级个体数量占比较大,分别为74.2%、86.7%、65.4%、52.0%、79.2%,径级大多为3~9 cm,可见该地区优势种群以小龄级个体为主。5个优势种群Ⅰ~Ⅱ龄级的个体数偏少且波动较大,优势种群总体表现为更新不足,种群稳定性较差。
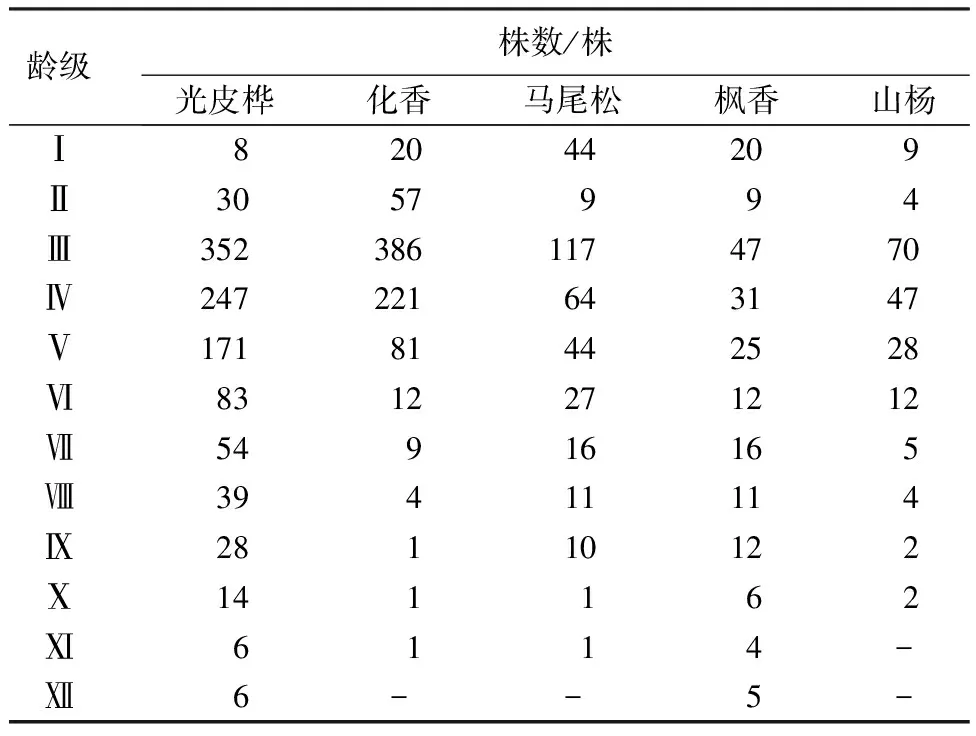
表2 5个优势种群龄级结构
3.3 优势种群数量动态分析
优势种群动态变化指数结果显示(表3),5个优势种群相邻龄级的动态指数随着龄级的增加呈不同的变化规律,5个优势种群相邻龄级的动态指数级划分见表3。光皮桦、化香种群动态指数V1均小于0,表明光皮桦、化香种群Ⅰ龄级个体数量少于Ⅱ龄级个体数量,在Ⅰ~Ⅱ龄级生长过程中,这2个优势种群的结构动态表现为衰退型。马尾松、枫香、山杨种群动态指数V1均大于0,种群结构动态表现为增长型。5个优势种群动态指数V2均小于0,种群出现衰退现象。光皮桦、化香、马尾松、山杨种群动态指数V3~V11均大于0,表现为个体数量呈增长态势。枫香种群在Ⅵ~Ⅶ、Ⅷ~Ⅸ、Ⅺ~Ⅻ龄级出现3次衰退(V6<0、V8<0、V11<0),表明在该种群在第Ⅵ、Ⅷ、Ⅺ龄级出现衰退。此外,5个优势种群整体动态指数均大于零,表明各优势种群动态受外部环境影响,种群数量出现波动,但总体仍呈增长态势。5个优势种群增长潜力由大到小依次为马尾松、化香、山杨、光皮桦、枫香。5个优势种群的种群数量动态变化指数也均大于0,其中,马尾松的最大,枫香最小。马尾松种群结构动态在面对外部环境随机干扰时,敏感指数最大,这与种群受外界干扰时其数量动态变化指数表现一致。这一现象说明,马尾松的增长潜力最大,受干扰的概率也最大,种群动态不稳定。光皮桦种群随机干扰风险概率极大值最小,反映出其受外部干扰的概率低,种群稳定性好。综上分析可知,5个优势种群数量动态呈现不同的变化规律,但是各种群总体表现为进展型,Ⅰ~Ⅱ龄级个体数量不足可能会阻碍种群稳定持续发展。
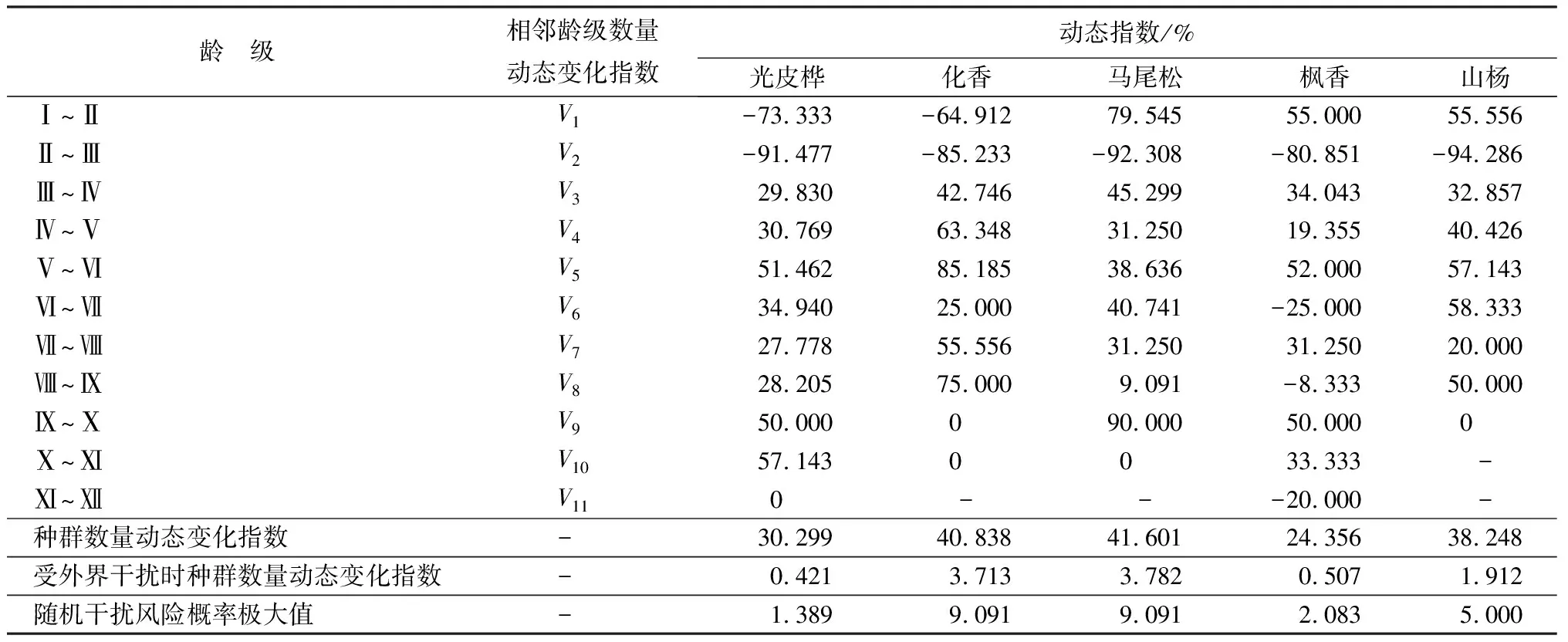
表3 5个优势种群动态变化指数
3.4 优势种群时间序列预测
时间序列预测结果表明(图1),5个优势种群数量呈大致相同的变化趋势,表现为种群数量前期骤减而后期稳定增长。各优势种群在未来龄级均出现种群个体数量优势减弱的现象,此后,随着龄级的增加,个体数量衰退现象逐渐消失,之后呈增加趋势。其中,5个优势种群在经过6个龄级后个体数量增幅最为明显。就整个种群的发展趋势进行综合分析,经过2、4、6、8个龄级时间以后,5个优势种群各龄级个体数的最大值与预测相比,逐渐往后移动,中、大径级的林木株数均呈现不同幅度的增长,种群总体发展较好。经过6个龄级后,各种群结构逐渐趋于合理,呈稳定增长态势。

图1 5个种群数量动态时间序列预测
4 讨论
林木种群龄级结构是树种生物学特性在环境影响下选择和适应的结果[23],可以反映种群内部个体数量配置状况和种群数量动态变化规律[24-25]。幼龄阶段尤其是幼苗期是植物种群更新发展过程中最重要、最敏感的时期[26-28],缺乏可更新的幼苗个体将阻碍种群稳定、持续发展[29-31]。本研究中,喀斯特次生林5个优势种群的龄级结构分析结果显示,光皮桦、化香、马尾松、枫香、山杨种群龄级结构均呈不规则金字塔型,其中第Ⅰ龄级、第Ⅱ龄级个体数极度匮乏,表明各种群潜在更新能力较弱。出现该现象的主要原因是该次生林经撂荒演替后,保存恢复良好,目前主林层已郁闭,林分密度过大,林下光照严重不足,而5个优势种群均为喜光阳性树种,种子萌发及幼苗生长对光照要求较高,所以这2个龄级的幼苗成活率较低。此外,由于喀斯特地区生境条件差,土层浅薄,岩石裸露程度高,因此种子萌发、幼苗形成及早期生长受到制约,从而导致第Ⅰ龄级、第Ⅱ龄级个体数量有限,成为各优势种群更新、发展的瓶颈。
种群数量特征可以有效地反映植物对生存环境的适合度和受干扰程度,并能够预测其未来数量动态及发展趋势[32-34]。种群数量动态分析结果表明,5个优势种群均属于进展型种群,但数量动态存在明显差异。光皮桦、化香种群在Ⅰ~Ⅱ龄级时,种群结构动态表现为衰退型,二者均为中性偏阳性树种,种子均表现出“体小质轻量大”的特征,成熟后主要以风为媒介进行传播。群落形成后,当种子停落在不利于种群更新的生境中,会出现种子难以萌发或萌发后的幼苗因难以适应环境而出现死亡的现象。马尾松、枫香、山杨种群在Ⅰ~Ⅱ龄级时,结构动态表现为增长型,原因主要是在喀斯特特殊生境的长期选择下,马尾松幼苗已经可以适应环境中的胁迫机制,萌发并建成。已有研究表明,枫香种群光补偿点较低,在林下弱光的生境中具有一定的阴生叶特性,可以忍耐较低的光强度[35];山杨种群幼苗生长力旺盛,可以最大程度地利用林分内的营养空间。因此,马尾松、枫香、山杨幼苗在林下更具竞争优势,成活率相对较高。此外,枫香种群在Ⅱ~Ⅲ、Ⅵ~Ⅶ、Ⅷ~Ⅸ、Ⅺ~Ⅻ龄级出现4次衰退。这是由于枫香的种子为蒴果具窄翅,风力传播后其着落点有很大的随机性,萌芽具有不确定性;此外,枫香个体生理性衰退也会造成种群个体消亡。实地调查发现,部分枫香植株经采伐后,其伐桩着生大量萌条,其幼苗和幼树对环境的适应能力较差,生长表现的差异也较大,因此导致了该种群龄级结构波动较大。时间序列预测显示,5个优势种群未来经过2、4、6、8个龄级后,前期种群数量锐减,但很快能够得到补充,后期种群数量趋于稳定。经过6个龄级后,各种群结构逐渐趋于合理,呈稳定增长态势。5个种群的发展趋势呈现不同规律。山杨在发展过程中由于缺乏中大径级个体,所以被逐渐淘汰,难以持续;化香由于本身属于小乔木,在林分发展过程中,不具竞争优势,也将被逐步淘汰;光皮桦、马尾松、枫香都属于高大乔木,作为群落的先锋树种,在喀斯特自然环境的长期选择下,种群个体数量相对稳定,在森林群落演替过程中长期处于优势层。因此可以推断,随着次生林群落的自然演替,枫香、光皮桦、马尾松的数量优势将越来越明显。
目前,研究区次生林自撂荒以后恢复效果良好,群落总体呈进展演替趋势,但由于缺乏科学的经营管理措施,林木干形不良、郁闭度过高、密度过大、天然更新匮乏的问题依然严重。通过探究该地区优势种群结构特征及数量动态,建议进行适度疏伐以减小林分密度、伐除病害木及干形质量差的林木以提升林分质量。采伐造成的林间空隙也会促进幼苗更新发展。研究区可结合封育保护,禁止乱砍滥伐、过度放牧、火灾等人为干扰活动,避免次生林出现逆行演替,有效实现其稳定及可持续发展。
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11 03:44:34
乡村地理(2022年2期)2022-05-30 10:48:04
环境与生活(2021年5期)2021-06-24 18:09:52
河南农业大学学报(2020年2期)2020-05-22 02:56:30
山西林业科技(2019年1期)2019-05-27 12:24:52
现代农业研究(2017年11期)2018-01-12 21:03:03
辽宁林业科技(2017年4期)2017-06-22 10:23:36
辽宁林业科技(2017年4期)2017-06-22 10:23:36
现代园艺(2017年20期)2017-02-03 18:49:29
当代贵州(2016年23期)2016-07-21 08:55:34
