
高压静电处理菜种对小菜蛾种群及生化特性的影响
2023-06-25 01:39:20邵雅婷罗继分胡佳振徐世才
延安大学学报(自然科学版) 2023年2期
贾 丽,邵雅婷,罗继分,胡佳振,郭 娇,徐世才*,罗 坤*
(1.延安大学 生命科学学院;2.陕西省区域生物资源保育与利用工程技术研究中心,陕西 延安 716000)
延安地处黄土高原的中南部,东傍黄河,属温带半干旱区,土层深厚,日照充足,完全符合蔬菜生长的多项主要气候指标。近几年,延安地区设施蔬菜种植面积达22.09 万亩,设施蔬菜产业已成为延安农村经济发展和农民持续增长的高效产业,在脱贫攻坚和乡村振兴中发挥了巨大作用。随着种植面积的不断扩大,病虫害种群数量也随之增加,成为设施蔬菜提质发展的一道阻碍,对设施蔬菜行业健康发展造成极大的威胁。
小菜蛾(Plutella xylostellaL.)属鳞翅目(Lepidoptera)菜蛾科(Plutellidae),又名两头尖、吊丝虫,是危害上海青、白菜、甘蓝等十字花科蔬菜的重要害虫[1]。该虫年发生世代多,繁殖率高,世代间虫口数量增长快,危害极为严重,主要以幼虫形态进行危害,共计4个龄期,其中4龄幼虫食量最大[2],可在短时间内将蔬菜叶片啃食干净,所以虫害一旦暴发,菜田会遭到大面积破坏[3]。在农业生产上,对于小菜蛾的防治主要依赖于杀虫剂,但由于使用频次多且药物种类单一,导致小菜蛾抗药性发生和发展速度急剧加快[4-6]。据不完全统计,小菜蛾大约已对50 多种杀虫剂产生了抗药性[7-8]。另外,杀虫剂在造成农药残留的同时,还会对生态环境造成污染,并将自然环境中小菜蛾的天敌杀害[9],此外,随着人们健康观念的改变,使得市场对绿色无公害蔬菜的需求量逐年增加。因此,探寻绿色无公害的防治手段调控小菜蛾的群体数量,已成为新时期设施蔬菜提质发展的迫切要求。
近几十年来,随着远距离电力传输、医疗和通信设备的高速发展,导致环境中的静电场强度显著增加[10]。在静电生物学领域中最早被关注的就是高压静电场(High voltage electro static fields,HVEF)对种子的生物学效应[11]。已有研究表明,外加适当强度的静电场可以改变植物应对胁迫的能力。例如前人利用高压静电场处理作物或蔬菜的种子,由此来改变种子内部的抗氧化酶系的活性,从而提高种子的萌发率,加速植物的生长,增加产量,改变植物体内营养物质的含量[12-14]。此外,前期研究结果表明:将普通小麦种子直接暴露于强度为4 kV/cm的高压静电场处理20 min,显著增强了宿主的抗蚜性[15],为了进一步研究高压静电场处理麦种调控麦蚜的种群数量的潜在生化基础,对小麦叶片组织中抗氧化酶系的活性进行了深入的分析,研究表明,高压静电场处理主要通过提高小麦叶片的超氧化物歧化酶(Superoxide dismutase,SOD)与过氧化物酶(Peroxidase,POD)的活性以增强宿主的抗虫性[16-17]。这些研究结果为制定小菜蛾幼虫的无公害防治措施提供了重要线索。本研究将上海青种子直接暴露于强度为5.0 kV/cm 的HVEF 中于不同的处理时间,研究取食HVEF 处理后的上海青对小菜蛾种群的间接效应,以此探究该物理方法在小菜蛾无公害防治中的可行性,为发展小菜蛾的绿色防控手段提供实验依据。
1 材料和方法
1.1 供试植物及处理
本研究中使用的上海青种子由山东晟远种业公司提供。挑选大小均匀,籽粒完整的上海青种子,将其随机分成7组,每组100粒种子,进行3次重复,其中1组为空白对照(未用电场处理),其余放置于强度为5.0 kV/cm的高压静电场中进行处理,处理时间分别为5、10、15、20、40、60 min。
1.2 供试昆虫处理
小菜蛾采自陕西省延安市宝塔区正大宏园林绿化有限公司,将虫源带回试验室稳定繁殖5 代后作为供试虫源。虫源饲养于温度25±1 ℃,相对湿度55%±10%,光周期12 L∶12 D 的人工气候箱内。将HVEF 处理之后的上海青种子进行种植。待上海青成熟后采集新鲜叶片饲养小菜蛾同日卵孵化出的一龄幼虫,为了保证小菜蛾的生长和发育不受食物的影响,每天更换一次新鲜菜叶。饲养一龄幼虫直至全部成虫羽化,在此过程中收集小菜蛾的4 龄幼虫,使用液氮冷冻后放置于-80 ℃冷冻保存用于酶活测定。将羽化后的成虫按照雌∶雄=1∶1配对后饲养于一次性塑料杯中,饲养条件同上。每隔24 h 补充10%的蜂蜜水,更换产卵纸并记录成虫产卵量、产卵周期、前期及雌雄虫存活天数,每个处理30 对成虫。
1.3 仪器与试剂
WJ-Ⅱ高压静电发生器(江苏省无锡博雅电子技术有限公司);全波长酶标仪(Tecan,Sunrise),2X-600低速离心机(海门市其林贝尔仪器制造有限公司);HC-2518 高速离心机(安徽中科中佳科学仪器有限公司)。
BCA 蛋白浓度测定试剂盒、过氧化氢酶(Catalase,CAT)、总超氧化物歧化酶(T-SOD)、过氧化物酶(POD)活性和丙二醛(Malonaldehyde,MDA)含量测定试剂盒均购自于南京建成生物工程研究所有限公司。
1.4 抗氧化酶活性测定
粗酶液制备:将10 头初发育为4 龄幼虫的小菜蛾放入2 mL离心管中使用液氮速冻,然后使用电子天平准确称取重量,按照质量体积比为1∶9 加入生理盐水,冰水浴条件下使用研磨器进行机械匀浆,制成10%的组织匀浆,使用低速离心机2 500 r/min离心10 min,用移液枪吸取上清液转移至新的离心管中,放入超低温冰箱备用。
酶活性测定:总蛋白浓度、CAT、T-SOD、POD活性和MDA含量均根据试剂盒说明书进行操作,并略作调整。
1.5 数据处理
从软件中导出的产卵前期(Adult preoviposition period,APOP)、产卵期(Oviposition period,OP)、产卵天数(Oviposition days,OD)、逐日产卵量(Daily fecundity)、每雌产卵量等文件后和酶活数据使用TWOSEX-MSChart 软件进行生殖相关参数的统计计算。使用26.0 版SPSS 软件的单因素方差进行方差分析,各处理的比较采用Student-Newman-Keuls(SNK)法进行多重比较,各处理间的显著性差异均设为P<0.05水平。使用Graph Pad 8.0软件作图。
2 结果与分析
2.1 对小菜蛾种群繁殖参数的影响
以强度为5.0 kV/cm的HVEF中处理上海青菜种,可显著的影响小菜蛾的繁殖与发育参数。如表1所示,随着处理时间延长,小菜蛾的产卵前期和产卵期均逐渐延长,且不同处理时间下小菜蛾产卵前期均显著长于对照组。在所有不同处理时间的比较中,10、20和40 min处理下小菜蛾的产卵前期显著延长(F=6.293,d.f.=6,P<0.05),其中10 min处理下达到最大值6.26 d。不同处理时长并未显著影响成虫的产卵期(F=1.514,d.f.=6,P<0.05),但相比对照组,处理组的产卵期均有所延长。在不同处理时间下产卵天数在短时处理下有所上升,5 min 处理下产卵天数甚至高于对照组,但当处理时间大于10 min后,产卵天数呈现下降趋势,最低降至4.91 d,但处理组与对照组无显著差异(F=2.038,d.f.=6,P<0.05)。这也说明上海青种子处理时间越久,对小菜蛾生殖影响越大。主要体现在小菜蛾逐日产卵量随时间延长而逐渐减少,在15 min处理下逐日产卵量达到最低值7.93粒。另外,平均产卵量在不同处理下均呈下降趋势,最低达到55.43 粒/雌,其中短时处理比长时处理下降更为剧烈。
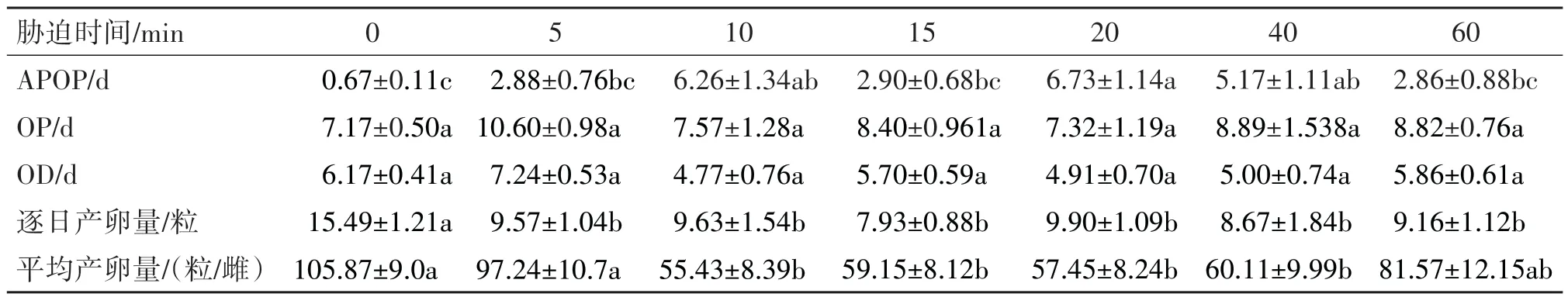
表1 取食HVEF处理的上海青之后小菜蛾的繁殖参数
2.2 小菜蛾体内抗氧化酶系的活性变化
单因素方差分析结果表明,当小菜蛾取食不同时间HVEF 处理的上海青之后,体内SOD 活性显著高于对照组(F=60.960,d.f.=6,P<0.05)(图1)。并且随着处理时间的延长,SOD 活性呈现出逐渐上升的趋势,其中60 min 处理时间下SOD 达到最大值68.768 7 U/mg。这表明HVEF 的间接作用对小菜蛾产生了一定的影响,因此小菜蛾体内的超氧化物歧化酶作出应答,从而使多余的超氧阴离子歧化产生H2O2在体内聚集。各处理中,10 min 处理组的响应最弱,不能及时分解多余活性氧,从而在虫体内积累。
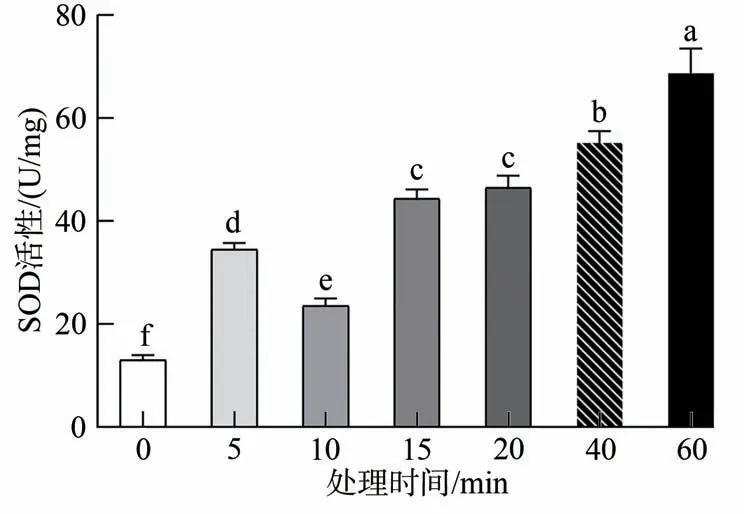
图1 取食HVEF处理的上海青之后小菜蛾体内SOD活性变化
通过单因素方差分析可知,当小菜蛾取食HVEF处理15、20 和40 min 的上海青之后,虫体内POD 活性显著低于对照组(F=46.618,d.f.=6,P<0.05),其中15 min 下降最为显著,POD 活性为14.408 3 U/mg(图2),这表明在这几个处理组中小菜蛾体内保护酶系统中POD 活力和平衡受到破坏,进一步使得机体内氧自由基大量积累,同时在SOD 酶帮助下歧化的超氧阴离子产生的H2O2无法及时被进一步分解,导致小菜蛾的生殖受到影响,产卵天数减少并且每雌产卵量大幅度下降。多重比较结果可知,60 min 长时处理下与其他处理时间均有差异,POD 活性显著上升,且达到最大值25.323 3 U/mg,在分解H2O2时起到重要作用。
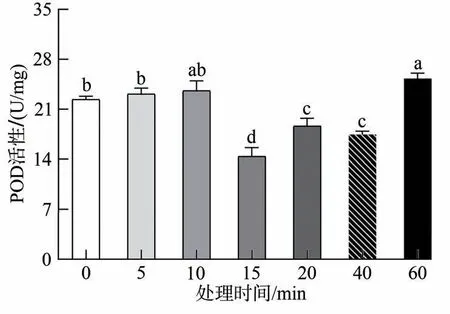
图2 取食HVEF处理的上海青之后小菜蛾体内POD活性变化
由图3 可知,取食HVEF 处理的上海青之后小菜蛾体内CAT活性的变化与POD 有相似的趋势,不同的是,在15 min 处理时间下CAT 显著上升,这也表明在此处理时间下CAT 在清除H2O2中起到重要作用。随着处理时间增加,处理组CAT 活性呈现先上升后下降的趋势,其中20、40 和60 min 显著下降(F=58.673,d.f.=6,P<0.05),在60 min 处理下有POD 活性达到最低值113.547 5 U/mg,因此在上述处理时间下H2O2无法被CAT 酶分解。上述结果表明,CAT 酶也参与抗氧化系统调控,但与SOD 酶不同,短时处理组中CAT活性显著高于长时处理组。
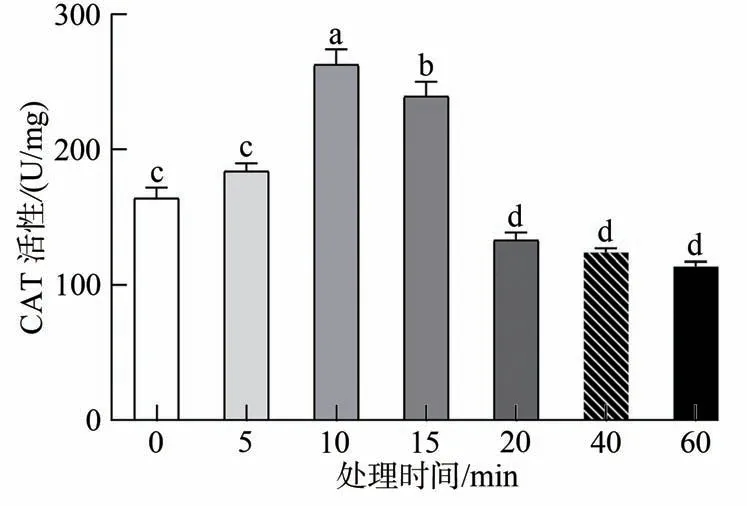
图3 取食HVEF处理的上海青之后小菜蛾体内CAT活性变化

图4 取食HVEF处理的上海青之后小菜蛾体内MDA含量变化
在取食不同处理时间的上海青之后小菜蛾体内MDA 的含量不同,其中10 和20 min 处理时间下小菜蛾MDA 含量显著高于对照组(F=19.020,d.f.=6,P<0.05),这表明在使用这两个处理时间下的上海青饲喂小菜蛾后机体内有细胞损伤,对其产生了不利影响。在取食5和15 min处理下的上海青之后小菜蛾体内MDA 含量与对照相比无显著差异(P>0.05),这主要是因为经SOD 歧化产生的H2O2被CAT 和POD 正常分解了,极大程度上减少了对虫体造成的损伤,对生殖的影响也随之减弱。
3 讨论和结论
有诸多研究表明,经过静电处理后,植物种子的萌发和幼苗的生长都会受到影响,并且这种影响会延续至植物幼苗中同时对昆虫产生间接影响。早在20 世纪70 年代,国内学者就已开始探索高压静电对种子萌发活力、生理生化变化、植株生长和抗逆能力的影响[18-19]。研究证明,高压静电能使黄瓜种子的发芽率,生长力得到增强[20]。高压静电场作用下,小麦种子抗冻害能力显著提升,植物中营养物质的含量亦发生了一定的改变[21]。此外,已有研究证明静电场处理可以促进菠菜新、陈种子的萌发及苗期生长,其中50 kV/5 cm×20 s 处理显著提高了菠菜新种子的发芽势、发芽率和萌发指数,同时高压静电也可提升陈化水稻种子活性指数,降低电导率,提升抗氧化酶活性[22]。本研究中利用高压静电处理上海青的种子,研究表明在适宜的处理时间下,能提升种子的发芽率,减少种子萌发的时间,在农业生产中可以减少种子在土壤中被虫害侵害的概率。
最初,进行电场生物学效应的研究主要是为探究环境电场改变能否对作物的生长乃至经济产量有促进作用,确定最佳的电场处理方式,诱发最大的生物学效应,从而开发出行之有效的农业物理新技术[23]。随着研究的不断深入,研究人员发现高压静电的效应不仅作用于植物自身,而且该效应还能间接作用于昆虫。研究表明,将小麦种子直接放置于4 kV/cm 的高压静电场处理后对麦长管蚜的性能产生不利影响[15]。另外,有研究通过测定小麦及麦长管蚜体内3 种抗氧化酶活性(SOD、POD、CAT)的变化,并对麦长管蚜的种群数量动态和模型进行分析,明确高压静电对小麦及蚜虫的作用[16]。植物体内抗氧化酶活性以及与抗性相关物质含量的变化改变了植物体内原本的微环境。当昆虫取食HVEF处理的植物后,高压静电作为一种胁迫因子使昆虫体内产生大量活性氧(ROS),正常生理状态下,机体内ROS产生和消除处于动态平衡,但当ROS积累过多时,就会引起脂质过氧化,从而对生物体造成损害,因此酶活力的变化能够反映生物体所受环境胁迫的状态和氧化损伤的程度[24-25]。与此同时,机体内的氧自由基能够攻击生物膜中的多不饱和脂肪酸(Polyunsaturated fatty acid,PUFA),引发脂质过氧化后形成脂质过氧化物,如醛基(MDA)、酮基等,这些物质会引起细胞代谢及功能障碍,甚至导致细胞死亡[26]。因此MDA 含量的变化常常作为细胞膜质过氧化程度的衡量指标,与抗氧化酶系的活性水平一起反映机体清除自由基的能力和受到胁迫的严重程度。本研究中在取食不同处理时间的上海青之后小菜蛾体内MDA 的含量不同,其中10、20 min处理时间下小菜蛾MDA 含量显著上升并且高于对照组,该结果与酶活结果保持一致,在10、20 min 处理时间下,SOD 酶响应胁迫活性显著上升,SOD 分解超氧阴离子后产生的H2O2在机体内积累,但机体内分解H2O2的CAT和POD活性却呈现下降趋势,导致H2O2在机体内大量积累,从而攻击细胞膜,造成脂质过氧化,因此脂质过氧化标志物MDA 含量提升,进一步反映出虫体受到严重的损伤,最终对小菜蛾幼虫的繁殖产生不良影响,使小菜蛾种群得到控制。在本研究中40 min 处理时间下MDA 含量的结果是出乎意料的,在此时间下CAT和POD 活性都是下降的,无法分解机体内积累的H2O2,导致细胞受损,因此MDA含量应该上升,但结果却与之相反,这值得我们进一步探究机体内其他的与ROS 分解相关的物质。
因此,利用高压静电场对上海青种子进行适当时间的处理,可以提高种子的萌发率,并能有效的抵抗小菜蛾,使其生长和发育受到不利影响,从而为调控小菜蛾的种群数量,制定生态防控小菜蛾的策略提供新的思路。在生产上减少农药的使用频次,降低生产成本,减少3R 问题的发生,更加绿色高效地提高上海青的产量和品质。
猜你喜欢
军民两用技术与产品(2022年6期)2022-08-06 07:31:44
植物保护(2022年1期)2022-02-10 23:33:07
农业技术与装备(2022年11期)2022-01-01 11:31:38
中学生数理化(高中版.高考理化)(2020年11期)2020-12-14 07:36:00
上海农业科技(2020年5期)2020-10-24 00:46:38
中学生数理化(高中版.高二数学)(2020年1期)2020-02-20 13:22:30
新高考·高一物理(2016年7期)2017-01-23 13:28:18
中学生数理化·高二版(2016年9期)2016-05-14 21:55:05
山西农业大学学报(自然科学版)(2015年3期)2015-04-19 02:46:41
河南科技(2014年24期)2014-02-27 14:19:40
