
不同放牧强度对高寒草甸植被群落结构和土壤理化性状的影响
2023-06-09 03:59安生辉
畜牧兽医杂志 2023年3期
安生辉
(甘肃省甘南藏族自治州草原工作站,甘肃 合作 747000)
草地不仅是保障我国食品结构和食品安全的基础资源,其还在水土保护、养分转化、气候调节、生物多样性和基因资源保护方面发挥着重要作用。由于气候变化、过度放牧、不合理的放牧制度等综合因素作用导致草地发生了不同程度的退化。据统计,我国90%的草地发生了不同程度的退化,并且以200 km2/年的速率增加。高寒草甸是我国现有18个草原类型中面积最大的一个草原类型,约占我国草原资源面积的的16%, 也发生了不同程度的退化。
放牧是高寒草甸最基本的利用方式,能显著改变草地植被群落结构和土壤理化性状。然而很多研究结果表明过度放牧可导致草地退化、沙化、物种多样性降低和草地生态功能失衡。如何确定适度、合理和科学的放牧制度至关重要。植被和土壤的理化性状与放牧强度密切相关,是草地是否健康可持续发展最直观的表现形式。本研究分析甘肃省甘南碌曲县放牧强度对高寒草甸植物群落和土壤理化性状的影响,揭示不同放牧强度与草地之间的内在联系,为合理高效利用高寒草甸、为畜牧业可持续发展提供参考。
1 材料与方法
1.1 研究地区概况
研究地处甘肃西南碌曲县,是甘肃省重要的畜牧县, 草场总面积39.4万hm2,占全县土地总面积的83.3%。试验地位于玛曲县尕海乡(lat.34°14'N; long.102°13'E),海拔3 650 m,研究区域属青藏高原气候带高原湿润气候区,冬长无夏,春秋短促,高寒阴湿,年均气温 2.3℃,无绝对无霜期,年降水量633~782 mm,该区域主要分为冬季放牧和夏季放牧两种利用方式。土壤类型为亚高山草甸土壤;草地类型为典型的高寒草甸草地,以莎草科的嵩草属(Kobresia)和疏丛型禾本科的针茅属(Stipa)、羊茅属(Festuca)、早熟禾属(Poa)的一些草种为主,并伴有大量的杂类草,如莓叶委陵菜(Potentillafragarioide)、条叶银莲花(Anemonetrullifolia)、火绒草(Leontopdiumhatioid)等,植被盖度在80%以上。
1.2 试验设计
2016年,本试验选定一个放牧户,其家庭草场面积约为178 hm2。根据王向涛等(2010)的划分方式,以该牧户夏季草场的居住点为起点,距起点2 800~3 200 m 的范围为轻度放牧(LG)区域;离居住点1 500~1 900 m范围内中度放牧(MD)区域,距居住点200~600 m为重度放牧(HG)区域;对照区域为该牧户的冬季牧场。试验采样于2016年8月10~18日进行。
1.3 草地植被特征测定
2016年8月分别从3个放牧梯度样地和对照区随机选择10个(50 cm×50 cm)进行植物群落调查,按照禾草、莎草、豆科、杂类草归类,带回实验室于烘箱内80℃、24 h烘干,测定生物量。调查项目包括草地植被高度、盖度、群落总盖度和物种丰富度。
1.4 土壤理化性状测定方法
在三个不同放牧梯度和对照样地内随机选取5个植被群落样方,用容积为100 cm3的环刀在0~15 cm土层收集土样,迅速装入铝盒,带回室内立即称重,再将土样放至 105℃烘箱烘至恒重,测定土壤容重和土壤含水量。同时,每个样点按照0~15 cm收集土壤样品,在室温下自然风干,充分研磨过0.25 mm的尼龙筛后,以便分析土壤pH、全氮、全磷、全钾、速效氮、速效钾和土壤有机质。参照鲍士旦主编的《土壤农化分析方法》,土壤有机碳( OC) 采用重铬酸钾法测定,用凯氏定氮法测定土壤全氮(TN)(本研究中土壤速效氮为硝态氮和铵态氮之和),土壤全磷( TP) 采用钼锑抗比色法测定,用火焰分光光度计测定土壤全钾( TK) 和速效钾( AK),土壤铵态氮( AN) 和硝态氮( NN) 采用连续流动分析仪测定,土壤速效磷( AP) 采用双酸浸提钼锑抗比色法测定。
1.5 数据分析
所有数据进行正太分布检验。试验数据统计分析使用SPASS19.0软件,用One-way ANOVA进行单因素方差分析,图表用Excel 2003绘制。
2 结果分析
2.1 不同放牧强度下植被高度、盖度、物种数和地上生物量的变化
从表1可以看出,不同放牧强度下的植被高度、盖度和地上生物量均显著低于零放牧区 (P<0.05);植被高度、盖度和地上生物量均表现为:对照(NG)>轻牧(LG)>中牧(MG)>重牧(HG)。物种数在中度最高,可达29种,其次依次为轻度、重度,对照区域物种数最低,只有19种(P<0.05);

表1 不同利用方式下草地植被高度、盖度、物种数和地上生物量的变化
2.2 不同放牧强度对4个功能群的影响
从图1中可以看出,对照区域的禾草生物量所占比例显著高于莎草、豆科牧草和杂草(P<0.05);轻度放牧区域的禾草生物量所占比例与莎草差异不显著,但禾草和莎草均显著高于豆科牧草和杂草(P<0.05);中度放牧区域的莎草生物量所占比例显著高于禾草、豆科牧草和杂草(P<0.05);重度放牧区域的杂草生物量所占比例显著高于禾草、莎草和豆科牧草,但莎草和禾草均显著高于豆科牧草(P<0.05)。
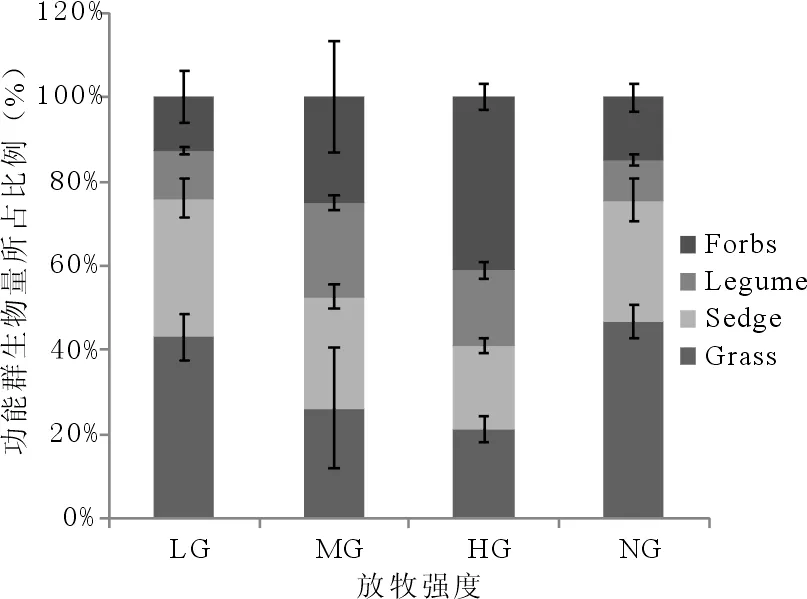
图1 不同放牧强度下4种功能群地上生物量所占比例
2.3 不同放牧强度对土壤物理性状的影响
从表2可以看出,与对照相比,中度放牧区域的土壤容重虽然略有下降,但差异不显著(P>0.05);轻度和中度放牧区域的土壤容重显著低于对照和重度放牧区域,且二者之间差异显著(P<0.05)。重度放牧区域的土壤含水量显著低于中度、轻度和对照,但中度、轻度和对照三者之间差异不显著(P>0.05)。

表2 不同利用方式下土壤理化性状的变化
2.4 不同放牧强度对土壤化学性状的影响
从表2可以看出,与对照相比,不同放牧强度下的土壤pH、全P无显著性差异变化(P>0.05)。随着放牧强度的增加,全氮、全钾和速效钾表现为先增加后降低的趋势,中度放牧下最高,分别可达4.72 g/kg,76.8 g/kg和131.6 mg/kg。速效氮在中度放牧下达到最高,可达54.6 mg/kg,显著高于轻度和重度放牧区域(P<0.05)。速效磷表现为随放牧强度的增加而增加,但中度和重度区域无显著性差异(P>0.05)。土壤有机质随放牧强度的增加而减小,重度放牧条件下最小,为111.8 g/kg。
3 讨论
适度放牧能促进高寒草甸正向演替,然而,过度放牧可导致高寒草甸发生逆向演替,草原生产力下降,稳定平衡的生态系统遭到破坏,引起草原发生不同程度的退化。本试验研究发现,不同放牧强度下植物群落高度、盖度和地上生物量显著降低,轻度放牧能有效增加植物群落高度、盖度和地上生物量;同时,适度放牧能有效增加植被群落结构中莎草所占的比例。这从一定程度上验证了“中度干扰假说”中关于适度放牧可以维持草地较高的生产力。地上生物量在放牧区显著低于对照区域,主要是由于家畜长期践踏和采食造成的。
毒杂草在重牧强度时生物量达到最大,这与很多研究结果相一致,由于放牧强度的增加,草地群落结构发生改变,禾本科和莎草科的优良牧草减少,而中等和劣质牧草的毒杂草增加。
中度干扰下草地具有较高的生物多样性,即中等放牧强度能增加物种丰富度,这与江小雷等(2004)的研究结果一致。放牧强度较低时,高大禾草的光利用率高,为优势植物,种间的光竞争强度增大,对低矮植被的光利用率具有抑制作用;放牧强度适中时,由于家畜采食降低了群落中禾草优势植物的高度,导致种间光竞争强度减小,有利于那些竞争力弱的植物建殖。当放牧强度持续增加,适口性高、植株高的禾草优势物种减少,食物资源短缺后,家畜进而采食那些适口性差的植物,从而使物种多样性降低;除此而外,低矮的植物通常是毒草,这些有毒植物根系分泌物能够抑制其它牧草种子萌发和牧草生长,这也是造成物种多样性降低的原因之一。
土壤含水量是影响草地生产力的重要因子。重度放牧区域由于家畜的踩踏使表层土壤紧实度增加,同时由于植被盖度降低,阳光直射地面导致蒸发量增加,进而导致土壤持水能力降低,土壤含水量降低。本研究中的土壤含水量在对照组最高,轻度和重度放牧区域虽有下降,但差异不显著,在重度放牧强度下土壤含水量最低。放牧引起土壤容重变化最直接的原因就是家畜不同程度的践踏导致的。然而,本研究发现土壤容重从小到大依次为对照>中牧>轻牧>重牧,毫无规律可言,还需更长时间的观测才能发现是否放牧强度对土壤容重存在规律性的响应。
氮、磷和钾的补给对土壤肥力有着重要作用。很多有关青藏高原高寒草甸的研究表明,植物生长季适度放牧能有效提高土壤中全氮和全钾的含量。本研究也证实了这一点,适度放牧能有效提高土壤养分含量。然而,全磷含量差异不显著,这可能是因为高寒草甸土壤磷含量少,同时还受气候环境等因素的影响,所以导致放牧强度对磷元素的影响不大。土壤中速效养分的含量反应了土壤实际提供养分能力。速效养分与温度、含水量和土壤微生物活动密切相关。本试验研究表明,高寒草甸土壤速效钾含量随放牧强度增加而降低。这可能是因为随放牧强度的增加,家畜采食量增大,牧草不断进行补偿性生长,增加了对这些速效养分的利用,导致土壤中速效钾含量降低。速效氮和速效磷随放牧强度增加呈先增加后降低的趋势。这很有可能是家畜适宜的采食和践踏能改变土壤微生物群落结构,进而引起氮和磷向其他组分的转换。高寒草甸气候寒冷,土壤微生物活动弱,土壤中有机物分解速率缓慢,因而放牧是改变土壤中碳元素的主要因素之一。我们的试验表明,高寒草甸土壤有机质含量随放牧强度增加而降低,与其他的研究结果一致。有机质的这种变化可能是因为放牧活动使植物不断进行补偿性生长,进而从土壤中吸收大量养分,减少了土壤有机质含量;也有可能是采食和践踏使植物枯落物含量减少,导致土壤中碳输入量减小。
4 结论
(1)植被盖度、高度和地上生物量均随放牧强度增加而降低,物种在中度放牧下最高;适度放牧能显著增加莎草比例,杂草随放牧强度增加而增加。(2)全氮、全钾和速效钾随放牧强度增加呈先增加后降低的趋势;速效磷随放牧强度增加而增加。(3)适度放牧能显著改良植被群落结构和土壤理化性质,有利于高寒草甸的可持续性利用。
猜你喜欢
疯狂英语·新读写(2023年4期)2023-05-10
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
黑龙江工程学院学报(2020年5期)2020-10-21
水土保持研究(2019年6期)2019-10-19
活力(2019年21期)2019-04-01
现代园艺(2017年21期)2018-01-03
绿色科技(2016年16期)2016-10-11
防护林科技(2015年5期)2015-06-10
海南热带海洋学院学报(2015年5期)2015-03-14
