
LncRNA JHDM1D-AS1对MPP+诱导的帕金森细胞模型线粒体功能的机制研究
2023-06-07 06:41李斯琴阿日奔吉日嘎拉巴彦淖尔市医院神经内科脊柱外科内蒙古巴彦淖尔015000
现代检验医学杂志 2023年3期
李斯琴,王 兴,阿日奔吉日嘎拉(巴彦淖尔市医院.神经内科;.脊柱外科,内蒙古巴彦淖尔 015000)
帕金森病(Parkinson’s disease,PD)是老年人常见的神经退行性疾病之一,发病率呈逐年上升趋势,然而该病的发生受到社会因素、药物因素、患者因素等多种因素的影响,导致其病因及发病机制至今尚未明确[1-2]。既往研究显示,线粒体功能障碍是导致神经系统中细胞凋亡的主要原因,也被认为是引起PD 患者体内活性氧(reactive oxygen species,ROS)累积的主要原因之一,在PD 发病过程中起着重要作用[3]。因此,寻找调节神经元细胞线粒体功能的生物学标志物,并深入探讨其分子机制,对于治疗PD 疾病具有重要的临床意义。沉默调节蛋白1(SIRT1)是一种具有烟酰胺腺嘌呤二核苷酸依赖性的蛋白去乙酰基酶,可以调节细胞寿命、能量代谢及神经保护等[4]。有研究报道,SIRT1 激活剂通过激活SIRT1 依赖性PGC-1α/NRF1/TFAM 信号通路,来改善线粒体生物合成受损功能,从而预防神经发育性疾病[5]。长链非编码RNA(long non-coding RNA,LncRNA)不仅可作为遗传信息中间载体的辅助性角色,而且更多地承担了各种调控功能,已被证实在脑发育、阿尔茨海默症及PD 疾病中发挥重要调节作用[6]。JHDM1D反义RNA 1(JHDM1D antisense RNA 1,JHDM1DAS1) 是一种新发现的 Lnc RNA,定位于染色体7q34,有研究报道,JHDM1D-AS1 在神经发育、细胞分化、氧化应激等多个生物学过程中起关键作用[7]。本研究通过生物信息学预测发现,SIRT1可能是JHDM1D-AS1 的靶基因,那么,JHDM1DAS1 是否能通过调控SIRT1 在PD 细胞模型中发挥潜在作用呢?故本研究利用多巴胺能神经毒素1-甲基-4-苯基吡啶离子(MPP+)处理 SH-SY5Y 细胞构建PD 模型,并检测JHDM1D-AS1 差异表达对PD 模型细胞凋亡及线粒体功能的影响,从而深入探讨JHDM1D-AS1/SIRT1 轴在PD 进展过程中的可能分子机制。
1 材料与方法
1.1 研究对象 人神经母细胞瘤细胞SH-SY5Y 购自中国科学院细胞库,进行复苏、传代培养后,用于后续实验。
1.2 仪器与试剂 DMEM 培养基(美国Gibco 公司);1-甲基-4-苯基吡啶离子(MPP+)(美国Sigma 公司);LJHDM1D-AS1 mimic,JHDM1DAS1 siRNA 序列构建和RT-PCR 引物由上海生工生物工程有限公司设计合成;AnnexinV-FITC/PI 细胞凋亡试剂盒(美国BD 公司);ROS 试剂盒(上海通蔚生物科技有限公司);双荧光素酶基因检测试剂盒(上海碧云天生物技术有限公司);兔源SIRT1,鼠源β-actin 抗体及HRP 标记羊抗兔(鼠)II 抗(美国Abcam 公司);PCR 仪(德国Biometra);细胞培养箱(日本三洋公司);低温高速离心机(美国Thermo 公司);流式细胞分析仪(美国BD 公司)。
1.3 方法
1.3.1 细胞培养:采用含10g/dl 胎牛血清的DMEM培养液,加入100 U/ml 青霉素和100 μg/ml 链霉素,37 ℃,5ml/dl CO2孵育箱中培养,当细胞融合度为80%时,胰蛋白酶消化传代,待细胞长至取对数生长期后进行后续实验。
1.3.2 PD 模型构建及细胞转染:取对数生长期SH-SY5Y 细胞接种于6 孔板,在其培养液中加入MPP+试剂(终浓度为5 mmol/L),轻轻摇晃混合均匀,恒温孵育24 h,构建PD 模型;将正常培养的细胞设为对照组(control 组)。将MPP+诱导的SH-SY5Y 细胞重新接种于6 孔板中,待细胞贴壁时将JHDM1D-AS1 mimic 和JHDM1D-AS1 siRNA序列转染至SH-SY5Y 细胞中,分别记为JHDM1DAS1 mimic 组和JHDM1D-AS1 siRNA 组,培养6 h后更换新鲜培养液继续培养48 h,收集细胞检测转染效率。
1.3.3 CCK-8 法检测细胞活力:取对数生长期SHSY5Y 细胞,胰酶消化、重悬,接种于96 孔板,每孔5×103个细胞,37 ℃,5ml/dl CO2孵育箱培养24 h,加入MPP+处理24 h,每组设3 个重复孔,后分别培养1~5 天,吸出培养液,每孔分别加入CCK-8液继续孵育2 h,在450 nm 处测量各组吸光度值。
1.3.4 RT-PCR 检测转染效率:利用RNA 提取试剂盒提取各组SH-SY5Y 细胞总RNA,采用反转录试剂盒合成cDNA,反转录条件为:42 ℃孵育30 min,85 ℃灭活5 min。以反转录合成的cDNA作为模板,进行PCR 扩增,扩增条件为:94 ℃ 5 min;94 ℃ 30s,60 ℃ 30s,72 ℃ 45s,40 个循环。扩增后的样品经PCR 仪测定,采用2-ΔΔCt法计算各组细胞中JHDM1D-AS1 相对表达量,以β-actin为内参,引物序列为:JHDM1D-AS1 上游引物:5’-TGTTGTTCTGTCACCCACCC-3’,下游引物:5’-TGCATGGGTCTTTCCACTCC-3’;β-actin 上游引物:5’-CTCCATCCTGGCCTCGCTGT-3’,下游引物:5’-GCTGTCACCTTCACCGTTCC-3’。
1.3.5 细胞凋亡水平检测:取转染后各组SH-SY5Y细胞,PBS 洗涤2 次后,离心,重悬,每组加入3μl Annexin V-FITC,室温避光孵育10 μl,后再加入3μl PI 染液,室温避光孵育15 min,离心去上清,每管加入200 μl 1×Bingding buffer,混匀后在1h内行流式细胞仪检测细胞凋亡情况。
1.3.6 线粒体膜电位变化:取转染后各组SHSY5Y 细胞,重悬,分别加入JC-1 荧光染料,浓度为 1 μg/ml,37 ℃孵育30 min,离心,收集底部细胞,加入1 ml PBS 缓冲液轻轻漂洗,同等条件下离心后收集细胞,加入提前预热的500 μl PBS 缓冲液,轻轻弹起细胞使其混合均匀,经流式细胞仪检测。
1.3.7 ROS 含量测定:收集转染后SH-SY5Y 细胞悬液,加入200 μl 浓度为5 μmol/L 的 Cell Rox Red 荧光染料,轻摇混匀,37 ℃避光孵育30 min,离心去上清,加入1 ml PBS 缓冲液轻轻漂洗后,再次离心去上清,加入300 μl PBS 缓冲液,轻摇混匀后通过流式细胞仪进行检测。
1.3.8 Western blot 检测SIRT1 蛋白表达情况:取转染后SH-SY5Y 细胞,胰蛋白酶消化,加入蛋白裂解液提取细胞总蛋白,BCA试剂盒检测蛋白浓度。取蛋白样品进行SDS-PAGE 凝胶电泳,后转PVDF膜,脱脂牛奶封闭1 h,加入SIRT1 和β-actin 一抗(1∶1 000),4 ℃孵育过夜,次日加入IgG-HRP二抗,室温孵育1 h,PBS 漂洗3 次,加入ECL 发光液,暗室曝光并拍照,采用Image-J 软件分析SIRT1 蛋白相对表达。
1.3.9 双荧光素酶基因报告实验:分别构建含有野生型和突变型 SIRT1 基因的荧光素酶报告基因质粒,将其与JHDM1D-AS1 mimic 及control 共转染于SH-SY5Y 细胞中,37℃孵育48 h 后,利用双荧光素酶活性检测试剂盒检测荧光强度。
1.3.10 SIRT1 激活剂处理后SH-SY5Y 细胞线粒体膜电位变化:为进一步证实JHDM1D-AS1 通过靶向作用SIRT1 调控PD 细胞线粒体功能,研究在上述转染了JHDM1D-AS1 mimic 的细胞中加入了SIRT1激活剂Resveratrol 溶液,恒温孵育24 h,后通过流式细胞仪分析SH-SY5Y 细胞线粒体膜电位变化。
1.4 统计学分析 采用SPSS 23.0 软件进行结果分析,所有数据均采用平均数±标准差(±s)表示,两组间比较采用独立t检验,多组间比较采用单因素方差分析,组间两两比较采用LSD 检验;以P<0.05 为差异有统计学意义。
2 结果
2.1 MPP+对SH SY5Y 细胞活力的影响 见图1。研究采用MPP+处理SH-SY5Y 细胞构建PD模型,经CCK-8 检测显示,MPP+处理后1,2,3,4,5 天时SH-SY5Y 细胞活力分别为1.00±0.01,0.89±0.04,0.77±0.03,0.54±0.02,0.33±0.02,随着培养时间延长细胞活力逐渐降低(均P<0.05)。
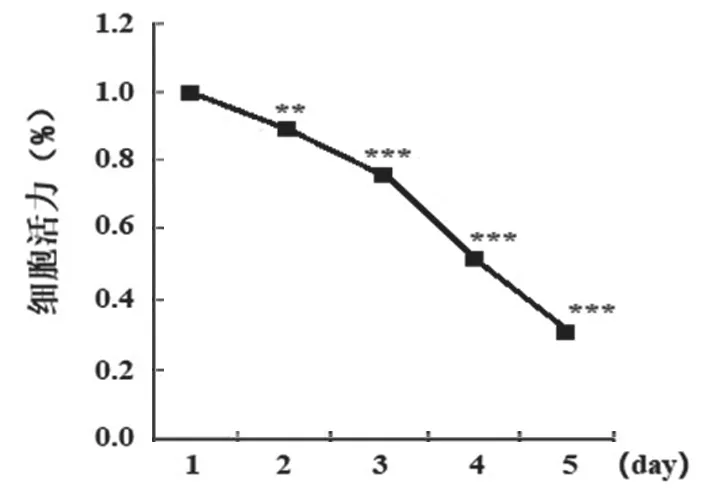
图1 MPP+处理对SH-SY5Y 细胞活力的影响
2.2 JHDM1D-AS1 在各组细胞中的表达 RT-PCR结果显示,control 组SH-SY5Y 细胞中JHDM1DAS1 相对表达水平为0.85±0.21,MPP+组细胞中JHDM1D-AS1 相对表达水平为1.25±0.33,较control 组增加,差异有统计学意义(t=62.017,P<0.05)。转染JHDM1D-AS1 mimic 和JHDM1DAS1 siRNA 序列后,两组细胞中JHDM1D-AS1相对表达水平分别为1.63±0.38 和0.72±0.17,与MPP+组相比,三组间差异亦有统计学意义(F=112.035,P<0.05),说明转染成功。
2.3 JHDM1D AS1 对MPP+诱导的SH SY5Y 细胞凋亡的影响 细胞凋亡结果显示,MPP+组SHSY5Y 细胞凋亡率为17.64%,转染JHDM1D-AS1 mimic 和JHDM1D-AS1 siRNA 序列后,SH-SY5Y细胞凋亡率分别为25.92%和10.74%,三组间比较差异有统计学意义(F=49.052,P<0.05),说明JHDM1D-AS1 过表达可促进MPP+ 诱导的SHSY5Y 细胞凋亡,沉默JHDM1D-AS1 表达则结果相反。
2.4 JHDM1D AS1 调控MPP+诱导的SH SY5Y 细胞膜电位变化 见图2。根据JC-1 染料依赖膜电位的极性发现,JHDM1D-AS1 过表达时,JC-1 以单体形式被释放到胞质内,绿色荧光增强,线粒体膜电位降低;沉默JHDM1D-AS1 表达时,红色荧光增强,线粒体膜电位升高,三组间比较差异有统计学意义(F=57.390,P<0.05)。
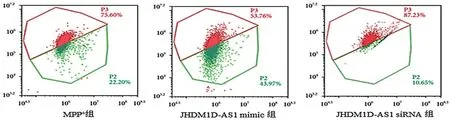
图2 JHDM1D-AS1 差异表达对MPP+诱导的SH-SY5Y 细胞线粒体膜电位的影响
2.5 JHDM1D AS1 对MPP+诱导的SH SY5Y 细胞中ROS 含量的影响 流式结果显示,MPP+组ROS相对荧光强度为27.58%±4.25%,转染JHDM1DAS1 mimic 后,SH-SY5Y 细胞中ROS 含量显著增加,其相对荧光强度为45.10%±6.05%;转染JHDM1D-AS1 siRNA 后,SH-SY5Y 细胞中ROS 含量显著减少,其相对荧光强度为14.82%±3.70%,三组间比较差异有统计学意义(F=25.794,P<0.05)。
2.6 JHDM1D AS1 与SIRT1 之间的靶向关系 Target Scan 软件预测显示,JHDM1D-AS1 与SIRT1 序列具有靶向结合位点,见图3。双荧光素酶报告基因显示,JHDM1D-AS1 过表达可降低WT-SIRT1 荧光素酶活性,而对MUT-SIRT1 荧光素酶活性无显著影响,进一步说明JHDM1D-AS1 与SIRT1 之间为负向调控关系,见表1。

表1 双荧光素酶报告基因检测结果

图3 JHDM1D-AS1 与SIRT1 的结合位点
2.7 JHDM1D AS1 对MPP+诱导的SH SY5Y 细胞中SIRT1 蛋白的影响 Western blot 结果显示,Control组SIRTI 蛋白相对表达为1.00±0.23,MPP+组SIRT1 蛋白表达下调为0.70±0.27,两组间差异有统计学意义(t=35.740,P<0.05)。转染JHDM1DAS1 mimic 后,SIRT1 相对表达为0.44±0.16,较MPP+组下调;转染JHDM1D-AS1 siRNA 后,SIRT1蛋白表达为1.34±0.22,较MPP+组上调,三组间比较差异亦有统计学意义(F=29.508,P<0.05)。
2.8 JHDM1D AS1 通过SIRT1 调控SH SY5Y 细胞线粒体膜电位 为了证实MPP+诱导的PD 模型中JHDM1D-AS1 是否通过SIRT1 调控线粒体功能,研究在MPP+诱导的SH-SY5Y 细胞中加入SIRT1激活剂Resveratrol后,SIRT1相对表达为1.46±0.34,较MPP+组0.70±0.27 明显上调,差异有统计学意义(t=3.032,P<0.05);在转染JHDM1D-AS1 mimic 组加入激活剂Resveratrol,SIRT1 相对表达为0.97±0.21,较仅转染JHDM1D-AS1 mimic 组0.44±0.16 相比明显上调,组间差异有统计学意义(t=3.477,P<0.05),表明SIRT1 激活成功。进一步经流式细胞仪检测发现,相比MPP+组,JHDM1D-AS1 过表达时线粒体膜电位明显降低,SIRT1 激活时线粒体膜电位明显升高,转染JHDM1D-AS1 mimic 并激活 SIRT1 后,线粒体膜电位水平回升,说明JHDM1D-AS1 的确可通过靶向调控SIRT1 影响MPP+诱导的SH-SY5Y 细胞线粒体功能,见图4。

图4 SIRT1 激活剂处理后JHDM1D-AS1 过表达对线粒体膜电位的影响
3 讨论
据调查,PD 是仅次于阿尔茨海默病的第二大神经退行性疾病,其发病率、致残率较高,严重影响着患者的生活质量[8]。目前而言,临床治疗PD 的药物及方法主要是缓解其症状,并不能阻止病情的恶化,因此,从基因水平寻找可有效干预PD 发生发展的靶向因子,是近年来医学工作者研究的热点及难点。有研究指出,JHDM1DAS1 是一种营养饥饿反应性 LncRNA,由组蛋白去甲基化酶JHDM1D 的反义链产生,具有抗氧化应激及神经保护作用[9]。SHI 等[10]研究数据显示,上调JHDM1D-AS1 表达,可以抑制Bcl-2 蛋白水平和eIF2α 磷酸化水平,进而导致干细胞凋亡,提示JHDM1D-AS1 在ROS 诱导的细胞凋亡中起促进作用。LIU 等[11]研究显示,JHDM1D-AS1 可能作为靶向miR-101-3p 的竞争性内源性RNA,结合在DUSP1 mRNA 的3’UTR 上,从而抑制神经元凋亡,发挥神经保护作用。WANG 等[12]研究发现,JHDM1D-AS1 可以调节MPP+诱导的神经元细胞凋亡、炎症和氧化应激,其作用机制可能是JHDM1D-AS1 通过miR-134-5p/PIK3R3 轴诱导神经元损伤,进而参与PD 的发生发展,提示JHDM1DAS1 与PD 疾病进展关系密切。
PD 最主要的病理特征是中脑黑质中多巴胺能神经元细胞选择性丢失,诱发线粒体功能异常、氧化应激,最终导致细胞凋亡,引起神经元细胞不可逆损伤[13]。广泛应用多巴胺能神经毒性物质建立体外PD 细胞模型是目前研究的重要手段。MPP+是多巴胺能神经元相对选择性神经毒素,是1-甲基-4-苯基-1,2,3,6-四氢吡啶(MPTP)的活性代谢产物,其能对中脑黑质多巴胺能神经元产生选择性破坏作用,研究证实MPP+诱导可产生与PD 相似的神经细胞氧化应激和凋亡等病理变化,是目前制备PD细胞模型的常用诱导剂[14-15]。而神经母细胞瘤细胞SH-SY5Y具有与多巴胺能神经元相似的生化特征,MPP+处理的SH-SY5Y 细胞被认为是高度模拟神经细胞线粒体功能障碍导致氧化应激的PD 细胞模型[14]。本研究中采用终浓度5 mmol/L MPP+诱导处理SH-SY5Y 细胞后发现细胞活力明显降低,细胞凋亡增加,ROS 含量增加,符合PD 病理改变,表明PD 细胞模型构建成功。进一步探究发现,MPP+诱导PD 细胞模型中JHDM1D-AS1 表达显著增加,转染JHDM1D-AS1 过表达序列后,促进了MPP+诱导SH-SY5Y 细胞凋亡、ROS 含量增加及线粒体膜电位降低。线粒体膜电位的稳定是线粒体及细胞发挥正常生理功能的基本保证,线粒体功能障碍是引起多巴胺能神经元损伤的重要机制[3]。大量研究已证实,不同刺激下细胞发生凋亡时常伴随线粒体膜电位的下降[16]。本研究中观察到转染JHDM1DAS1 过表达时线粒体膜电位降低,沉默JHDM1DAS1 表达则结果相反,说明JHDM1D-AS1 可能通过诱导线粒体功能异常,引起神经元细胞损伤,在PD 疾病发展过程中发挥重要作用,有望成为PD 诊断治疗的潜在靶点,但其具体的作用机制还有待进一步探讨。
SIRT1 是目前研究最深入的 Sirtuin 蛋白,它以组蛋白和多种非组蛋白为底物,通过其去乙酰化调节自噬、凋亡、衰老、炎症反应、氧化应激、基因转录等关键生物过程,在线粒体功能及能量代谢等多种过程中发挥关键作用[17-18]。CAMPOREZ等[19]研究表明,在阿尔茨海默症动物模型中SIRT1活性降低,究其原因可能是它通过去乙酰化上调PGC-1α,而后者激活线粒体生物发生。LI 等[20]研究指出,SIRT1 在PD 动物模型中表达下调,给予SIRT1 激活剂干预后,可以抑制小胶质细胞活化、减轻氧化应激和炎症反应,避免多巴胺神经元线粒体损伤。基于上述报道,推测JHDM1DAS1 是否通过靶向作用SIRT1 而影响PD 细胞模型的线粒体功能呢?本研究通过生物学软件预测发现JHDM1D-AS1 与SIRT1 存在结合位点,证实JHDM1D-AS1 可以负向调控SIRT1,说明JHDM1D-AS1 通过SIRT1 轴调节PD 模型细胞的生物学行为。其次研究通过转染SIRT1 激活剂上调SIRT1 表达,经流式细胞仪分析,SIRT1 过表达时线粒体膜电位升高,在转染 JHDM1D-AS1 过表达并激活 SIRT1 后,线粒体膜电位水平回升。研究结果提示,JHDM1D-AS1 过表达可以导致神经元细胞线粒体功能障碍,加速PD 疾病的发展进程,激活SIRT1 后,则可以保护线粒体功能,抑制MPP+诱导的SH-SY5Y 细胞凋亡,发挥神经元细胞保护作用。此外,前期有大量文献证实活性氧(ROS)来源于线粒体,并且通常情况下,细胞内的ROS处于较低水平状态,若ROS 水平过高,将会对细胞和基因结构造成严重损伤[21-22]。本研究证实,JHDM1D-AS1 过表达后ROS 含量增加,反之沉默其表达后ROS 含量减少,与既往研究报道的ROS大量累积,可导致线粒体代谢紊乱,细胞氧化应激反应增强,最终引起细胞的凋亡,导致神经元细胞不可逆损伤的结论相一致[23]。
综上所述,MPP+诱导的PD 细胞模型中,沉默JHDM1D-AS1 表达可以负向调控SIRT1 蛋白水平,抑制SH-SY5Y 细胞发生凋亡,减轻氧化应激反应,增加线粒体膜电位水平,保护线粒体功能,发挥了神经元细胞保护作用,在一定程度上为临床诊断和治疗PD 病提供新的思路及理论基础。但本研究还存在一定的局限性,后期可结合体内动物实验验证JHDM1D-AS1 在PD 疾病形成过程中的可能作用机制。
猜你喜欢
世界科学技术-中医药现代化(2021年7期)2021-11-04
海洋通报(2021年1期)2021-07-23
生物学通报(2021年4期)2021-03-16
教育教学论坛(2019年19期)2019-06-17
中成药(2017年9期)2017-12-19
中成药(2017年5期)2017-06-13
华南农业大学学报(2015年5期)2015-12-04
中国药理学通报(2014年2期)2014-05-09
癌变·畸变·突变(2014年1期)2014-03-01
微循环学杂志(2011年2期)2011-03-19
