
龙眼UBP 家族全基因组鉴定及其在外源胁迫下的表达分析
2023-06-07 11:17:16童宁张春渝许小琼陈晓慧申序赖钟雄
浙江大学学报(理学版) 2023年3期
童宁,张春渝,许小琼,陈晓慧,申序,赖钟雄
(福建农林大学 园艺植物生物工程研究所,福建 福州 350002)
泛素(ubiquitin)是一种普遍存在于真核生物中的由76 个氨基酸组成的多肽[1]。通过一系列酶将泛素分子与目标蛋白的特异性相结合,并引导其对蛋白酶体进行降解的过程称为泛素化。泛素化是一种常见的蛋白质翻译后的修饰方式,也是一种细胞内信号[2],在细胞分裂、自噬、消亡、DNA 修复、蛋白质降解等生物学过程中具有重要调控作用。通过蛋白酶特异地从与泛素分子结合的蛋白质底物中切割泛素分子的过程称为去泛素化,这些蛋白酶即为去泛素化酶(deubiquitinating enzymes,DUBs)。迄今为止,人类基因组中已被识别的去泛素化酶有100 余种,根据其氨基酸序列与保守结构域的不同,可分成6个家族:泛素特异蛋白酶(ubiquitin-specific protease,UBP,在哺乳动物中被称为USP)、泛素羧基末端水解酶(UCH)、卵巢肿瘤蛋白酶(OTU)、Machado-Joseph结构域蛋白酶(MJD)、MINDY 蛋白酶(MINDY)和JAMM 结构域蛋白酶(JAMM)。其中前五大家族属于半胱氨酸蛋白酶,最后一个属于锌金属蛋白酶[3]。UBP家族成员数量最多,超过50个,且结构多样化[4]。
UBP 家族成员具有2 个相似的催化残基三联体,每个三联体包含2 个短且高度保守的半胱氨酸盒和组氨酸盒,是催化位点的关键部分(半胱氨酸盒中的Cys,组氨酸盒中的His 和Asp/Asn)[5]。半胱氨酸盒与组氨酸盒的区域被称为泛素羧基末端水解酶(UCH)结构域,其在空间和结构上比较保守[6],但不同成员UCH 结构域之间的氨基酸序列不同,其氨基酸种类、数量的差异均较大[7]。目前对UBP基因的分析主要集中在单个基因的生物功能。如发现拟南芥中AtUBP1 和AtUBP2 基因可能与体内异常氨基酸代谢的调节有关[8],AtUBP12 参与了生物钟反馈调节,且可调控开花时间[9];哺乳动物中USP1 参与了胃癌的早期阶段转化[10],USP24 与细胞凋亡、细胞周期和DNA 的损伤修复有关[11];USP13 在小鼠单核巨噬细胞破骨分化中起重要作用[12]。已对UBP 家族进行全基因组鉴定的有拟南芥(27 个成员)[6]、毛竹(48 个成员)[13]、水稻(21 个成员)[14]等,尚未见对龙眼进行全基因组鉴定的报道。
龙眼(Dimocarpus longan Lour.),热带亚热带常绿经济型果树,因其假种皮富含多种磷质、氨基酸、维生素、类黄酮等[15],具有降血糖、补血健脑等作用[16]。外界环境以及自身的遗传特性和基因的表达调控对龙眼胚胎发育具有重要影响,而龙眼的胚胎发育与树体的生长发育和果实的品质产量相关。目前已知UBP 广泛存在于真核生物,能通过去泛素化作用调节生物的多种代谢过程,且已发现该家族部分成员参与胚胎生长发育,甚至会导致胚胎死亡,但关于龙眼DlUBP 基因在胚胎发育中的作用尚未见报道。因此,对DlUBP 基因进行分析有助于进一步了解DlUBP 家族在龙眼胚胎发育中的作用。通过对DlUBP 家族全基因组的鉴定,分析其基因结构、蛋白质结构、启动子顺式作用元件、进化关系,并分析龙眼体胚中DlUBP 基因在不同时间梯度的脱落酸处理和干旱胁迫处理下的表达情况,以期为UBP 基因家族在龙眼体胚或其他植物生长发育调控的研究提供参考。
1 材料与方法
1.1 材 料
1.1.1 数据来源
第3 代龙眼基因组数据库,龙眼转录组数据库,拟南芥(Arabidopsis thaliana)UBP 家族的基因序列和氨基酸序列下载于Uniprot 数据库(https://www.uniprot.org/),甜橙(Citrus sinensis)、水稻(Oryza sativa)UBP 家族的基因序列和氨基酸序列下载于phytozome 数据库(https://phytozome.html)。
1.1.2 实验材料
龙眼体胚发生过程中的胚性愈伤组织(embryonic callu,EC)来源于福建农林大学园艺植物生物工程研究所。参照赖钟雄[17]的培养方法,以EC 为起始材料获得不完全胚性紧实结构(incomplete compact pro-embrogenic cultures,ICpEC)和球形胚(globular embryos,GE)。选择培养20 d 的生长发育良好的龙眼胚性愈伤组织,分别接种于100 μmol·L-1脱落酸(abscisic acid,ABA)、10%聚乙二醇(PEG 6000)的MS 液体培养基中,于25 ℃、110 r·min-1黑暗条件下摇床培养0,4,8,12 h,收样,液氮速冻后-80 ℃封存于冰箱,用于后续qPCR 定量分析。
参照Tripue(Transgen,China)试剂盒说明书提取龙眼在EC、ICpEC、GE 3 个阶段以及经ABA 和PEG6000 处理的RNA,再参照PrimerScript RT Reagent Kit(TaKaRa)试剂盒说明书逆转录合成qPCR 所需的cDNA。
1.2 方 法
1.2.1 DlUBP 基因的鉴定
DlUBP 基因在染色体上的定位:利用NCBI BLAST 在线网站,将龙眼基因组数据库与模式植物拟南芥AtUBP 的氨基酸序列进行同源比对,对DlUBP 家族成员进行筛选,获得18条DlUBP 候选序列。结合保守结构域数据库(conserved domains database,CDD)和SMART 对蛋白结构域的预测分析,确定龙眼UBP 基因组中存在18 条DlUBP 序列。参考拟南芥的命名方式,命名DlUBP基因。最后根据龙眼基因组注释信息和DlUBP基因的ID,用TBtools软件在染色体上绘制DlUBP基因的位置分布。
DlUBP 基因结构及蛋白质分析:基于龙眼genome 文件,用TBtools 分析18 个DlUBP 基因的结构;从ExPASy 中查询18 个DlUBP 基因的蛋白质等电点、分子质量和氨基酸数等,并列表分析;通过MEME 在线软件分析DlUBP 的保守基序,并下载XML 格式的分析结果,用TBtools 软件进行可视化绘图分析。根据DlUBP 基因的氨基酸序列,用TBtools 软件中的Batch SMART 进行蛋白质结构域预测以及可视化分析。
UBP 家族进化树的构建:先采用MEGA5.05 软件中的邻近法(neighbor-joining method)单独对DlUBP 家族进行进化分析,然后采用相同的方法构建龙眼、拟南芥、水稻、甜橙4 个物种的UBP 家族氨基酸序列系统进化树,自展系数法(Boot strap)设置1 000 次重复检验。保存nwk 格式的进化树文件,使用iTOL 在线网站(https://itol.embl.de)对进化树进行上色编辑。
DlUBP 基因启动子分析:利用DlUBP 基因组序列和基因结构注释信息文件,经TBtools 软件提取获得18 条CDS 上游碱基对为2 000 bp 的DlUBP 序列,采用PlantCARE 在线网站(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)对DlUBP基因启动子特征以及顺式作用元件的功能进行预测分析,再用Excel 软件分类整理获得的数据,最后用TBtools软件对顺式作用元件进行可视化绘图。
1.2.2 DlUBP 基因在龙眼不同体胚发生阶段的表达分析
从龙眼基因组数据库中提取18 个DlUBP 基因在龙眼体胚发生过程中EC、ICpEC、GE 3 个阶段的特异表达每千个碱基转录每百万映射读取的碎片(fragments per kilobases per million mapped fragments,FPKM),提交至TBtools 软件进行热图绘制,并分析DlUBP 基因在不同体胚发生阶段的特异性表达情况。
1.2.3 DlUBP 基因表达量的qPCR 分析
为了解DlUBP 基因在脱落酸处理和干旱胁迫处理下的表达情况,将DlUBP2、DlUBP8-1、DlUBP12-1、DlUBP12-2、DlUBP17、DlUBP19、DlUBP22-2 和DlUBP23 等8 个基因的CDS 序列通过 Primer premier 6 设计正向引物和反向引物,再通过引物检测网站Beacon Designer Free Edition(http://www.premierbiosoft.com)识别得到的引物,选出符合要求的基因,并收集其引物,见表1。20 μL qPCR 体系包括10 μL HieffqPCR SYBRGreen Master Mix(Yeasen,Shanghai,China)染料,7.4 μL dd H2O,0.8 μL 正向引物和0.8 μL 反向引物,1 μL cDNA 模板。以elF-4a 为内参基因,用Roche LightCycler 480 实时荧光定量PCR 仪(Roche Applied Science,Switzerland)检测DlUBP 基因在不同时间梯度的脱落酸处理和干旱胁迫处理下的表达量,采用2-ΔΔCT方法计算DlUBP 基因的相对表达量,用Excel 软件整理分析,并绘制柱形图。
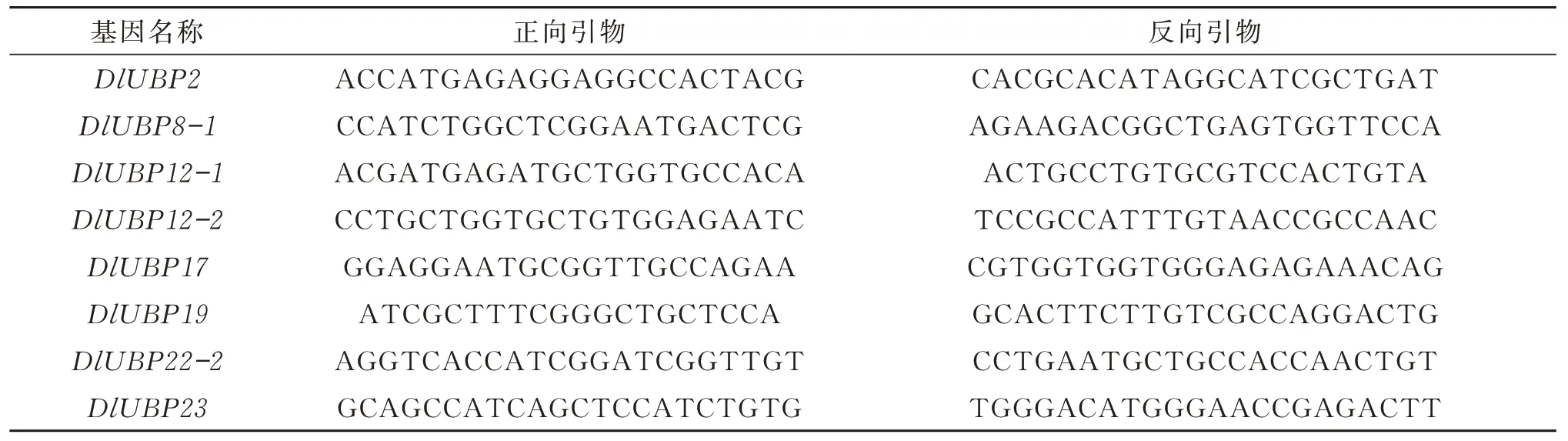
表1 实时荧光定量 PCR 引物Table 1 Real-time fluorescent quantitative PCR primers
2 结果与分析
2.1 DlUBP 全基因组鉴定与蛋白特性分析
通过与拟南芥基因组进行比对,利用CDD 和SMART 检测蛋白的结构域,共获得18 个DlUBP 基因。通过对龙眼与拟南芥、水稻和甜橙的UBP 家族成员的系统进化分析,参照拟南芥家族成员的命名方法,将DlUBP 家族成员分别命名为DlUBP2、DlUBP4、DlUBP5、DlUBP8-1、DlUBP8-2、DlUBP9、DlUBP12-1、DlUBP12-2、DlUBP12-3、DlUBP16、DlUBP17、DlUBP19、DlUBP21、DlUBP22-1、DlUBP22-2、DlUBP23、DlUBP25、DlUBP26。
通过分析DlUBP 全基因组的蛋白质理化性质,发现UBP 家族氨基酸数为369~1 117 个,相差较大,除 DlUBP2、DlUBP12-1、DlUBP12-2、DlUBP16 外,其余基因的氨基酸数在1 000 个以下;分子质量为42.11~130.80 ku;等电点为5.14~9.32,见表2。

表2 DlUBP 全基因组蛋白质特性Table 2 Characteristics of DlUBP gene family protein
2.2 DlUBP 基因染色体定位分析
根据染色体定位分析结果(图1),18 个DlUBP基因不均匀地分布在10 条染色体上。龙眼基因组已组装的染色体共有15 条,其中第2,3,8,10,11 号染色体上不存在DlUBP 基因。定位在第1 号染色体上的DlUBP 基因最多,有4 个,定位在第5,6,13,15 号染色体上的DlUBP 基因各有1 个,其余5 条染色体上分别有2 个DlUBP 基因。第12 号染色体上的2 个DlUBP 基因的物理位置较接近,但超过了100 kb,HUANG 等[18]指出,染色体上物理位置不超过100 kb 的2 个及以上的同源基因为串联重复基因,因此各DlUBP 基因不存在串联重复的情况。
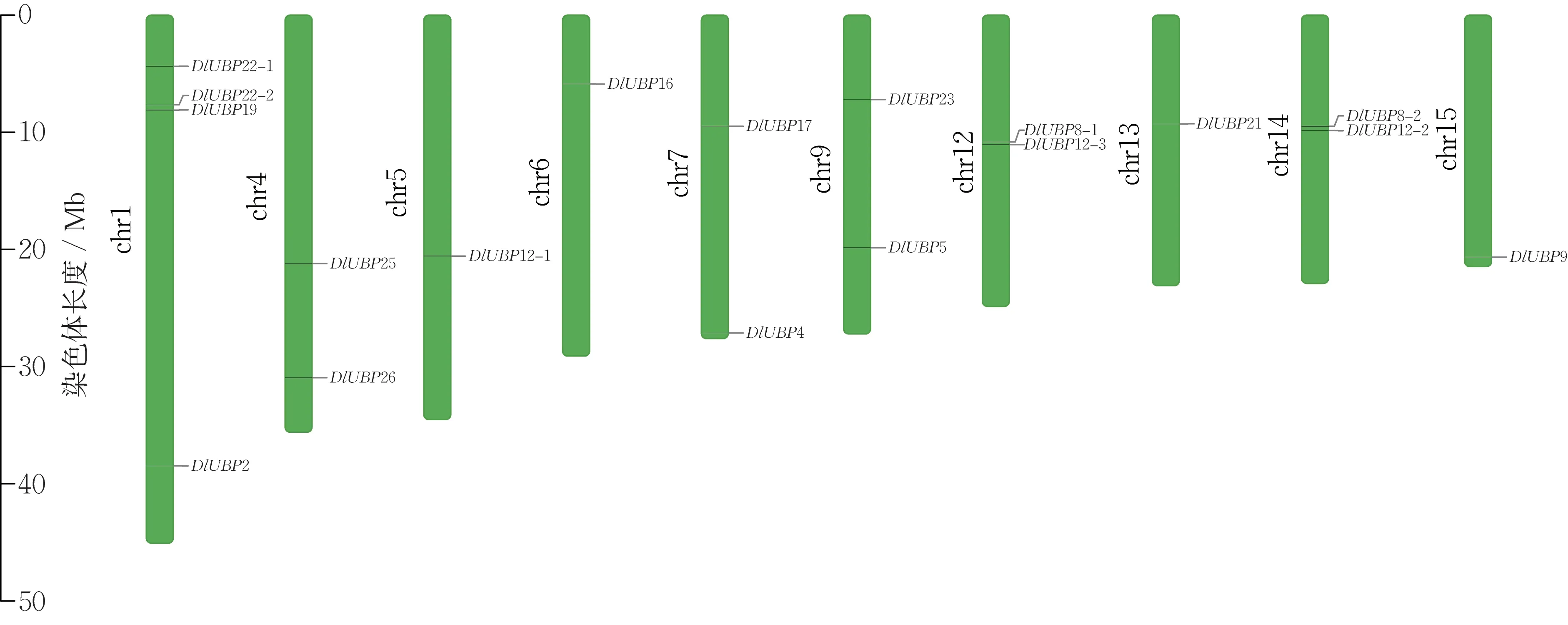
图1 DlUBP 基因在染色体上的定位Fig.1 Location of DlUBP gene on chromosome
2.3 DlUBP 基因结构与蛋白质分析
18 个DlUBP 基因的内含子数、外显子数及其位置如图2 所示,可知,DlUBP 基因内含子数为2~31个,其中DlUBP12-1 基因的内含子数最多,有31 个,其次是DlUBP12-2,有30个,DlUBP2和DlUBP22-2基因的内含子数最少,均为2个,大部分DlUBP基因的内含子数在20 以内。此外,DlUBP 基因的长度存在较大差异,DlUBP8-2 基因最长,DlUBP22-2 基因最短,这可能由内含子数差异较大引起,也可能导致各DlUBP基因之间的功能有所区别。
使用MEME 在线软件,搜索到25 个motif(图2)。从图2 中可以看出,motif 1、motif 2、motif 3、motif 5、motif 8 出现的频率较高,出现在18 个DlUBP 基因中,说明这5 个motif 在DlUBP 家族中较保守,其中motif 2 在DlUBP8-1 基因中出现了2次。其次是motif 4 和motif 6,出现在17 个基因中,特别是motif 6 在DlUBP9 基因中连续出现了2 次。motif 12 和motif 18 出现在大多数基因中。此外,在同一个小分支上,保守基序的种类和排列基本一致,这可能与其功能的保守性有关。
蛋白质结构域预测分析结果如图3 所示。由图3 可知,18 个DlUBP 基因均含有一个UCH 结构域,这体现了UCH 蛋白质结构的保守性。除共有的UCH 结构域,部分DlUBP 基因存在特异的保守区,如DlUBP2 和DlUBP22-2 均有一个ZnF-UBP 锌指结构,DlUBP12-1、DlUBP12-2 和DlUBP12-3 均有1 个 MATH 结构域,DlUBP16、DlUBP17 和DlUBP19 均有1 个zf-MYND 锌指结构,DlUBP5、DlUBP9 和DlUBP26 均有1 个DUSP 结构域。此外,同属于一个亚家族的成员在蛋白质结构域的种类和分布上比较相似,表明龙眼UBP 基因各亚家族拥有的生物学功能不同。
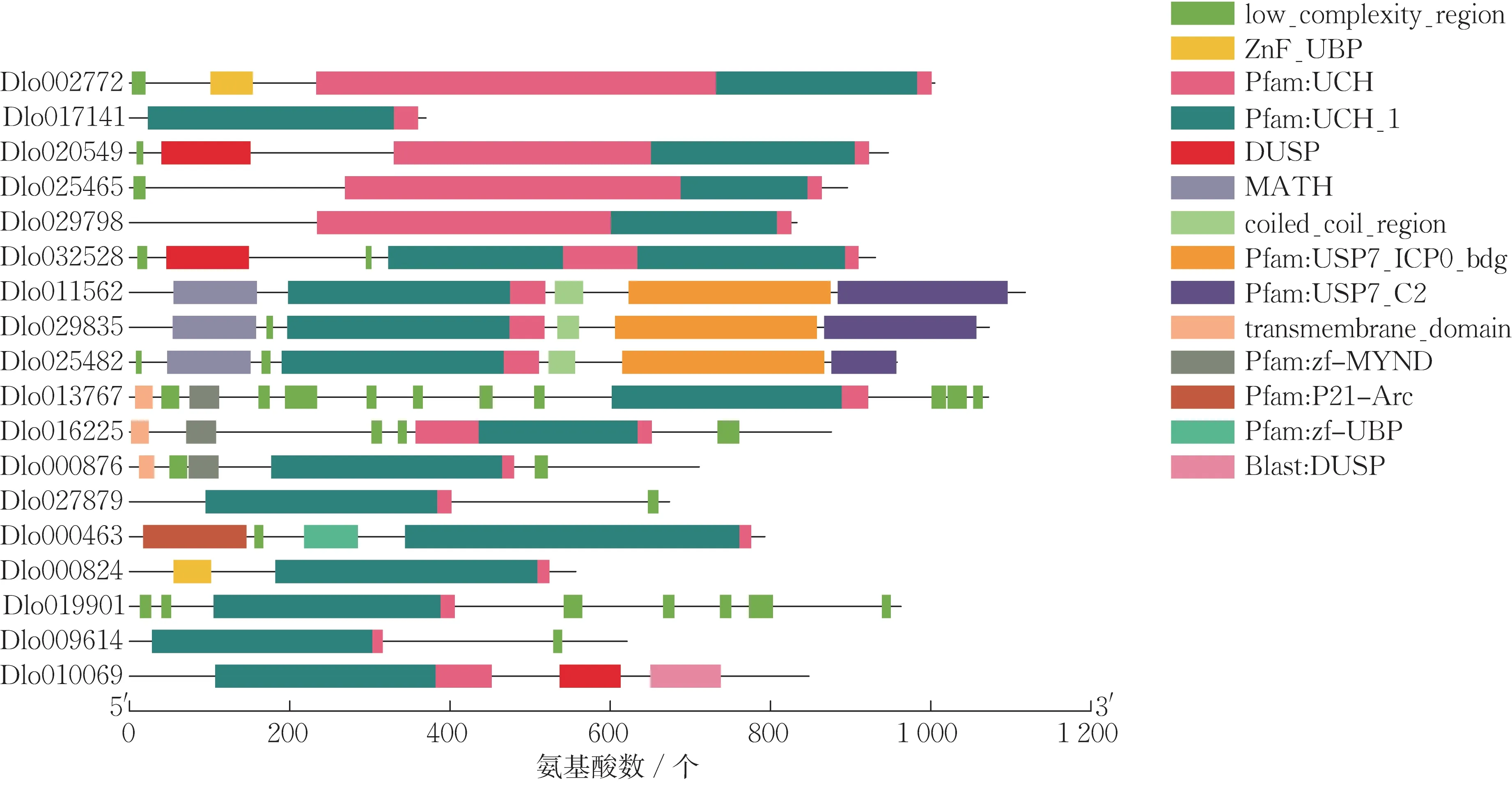
图3 DlUBP 家族成员的蛋白质保守结构域分析Fig.3 DlUBP family protein conserved domain analysis
2.4 植物UBP 家族成员的系统进化树分析
基于龙眼、拟南芥、水稻、甜橙的氨基酸序列,用MEGA5.05 软件构建了4 个物种的68 条氨基酸序列系统进化树,如图4 所示。参考拟南芥中AtUBP 基因的分类方式[8],采用SAMRT 蛋白质结构域分析,根据基因的蛋白质结构差异,将DlUBP 基因分为10 组:G1 的成员为DlUBP2,G2的成员为 DlUBP4,G3 的成员为 DlUBP5、DlUBP8-1、DlUBP8-2、DlUBP9,G5 的成员为DlUBP12-1、DlUBP12-2、DlUBP12-3,G7 的成员为DlUBP16、DlUBP17、DlUBP19,G8 的成员为DlUBP21,G9 的成员为DlUBP22-1、DlUBP22-2,G10 的成员为 DlUBP23,G12 的成员为DlUBP25,G13 的成员为DlUBP26。其中,G2 成员中包含1 个ZnF-UBP 锌指结构,G5 成员中包含1 个MATH 结构域,G7 成员中包含1 个zf-MYND锌指结构,G3 部分成员中以及G13 成员中包含1个DUSP 结构域。
2.5 DlUBP 基因启动子分析
使用PlantCARE 在线网站对DlUBP 基因启动子序列中可能存在的顺式作用元件进行预测分析,结果如图5 所示。可知,18 个DlUBP 基因的启动子序列均含CAAT-box 和TATA-box,且CAATbox 的数量比TATA-box 的多,说明DlUBP 基因都能够正常转录。18 个DlUBP 基因均含有较多的光响应元件,大部分DlUBP 基因具有厌氧诱导响应元件,响应脱落酸和茉莉酸甲酯激素应答;50%以上的基因具有干旱胁迫下MYB 转录因子的结合位点和防御与应激响应元件;44%左右的基因具有低温响应元件、光胁迫下MYB 转录因子的结合位点和MYBHv1 结合位点;33%左右的基因响应生长素、水杨酸和赤霉素应答;少数基因具有胚乳表达、缺氧特异性诱导响应元件和参与类黄酮生物合成基因调控的MYB 结合位点,参与昼夜控制和玉米醇溶蛋白的代谢调控;与种子特异调控有关的响应元件只存在于DlUBP5 基因中。由此可知,DlUBP 基因具有较多光响应、激素响应元件以及与植物生长发育有关的其他顺势作用元件,而且不同的基因具有的顺式作用元件的种类及数量不同,表明DlUBP 基因可能在胚胎发育、种子生长、植株生长发育和抗逆等过程中有重要作用,且不同的DlUBP 基因所起的作用不同。
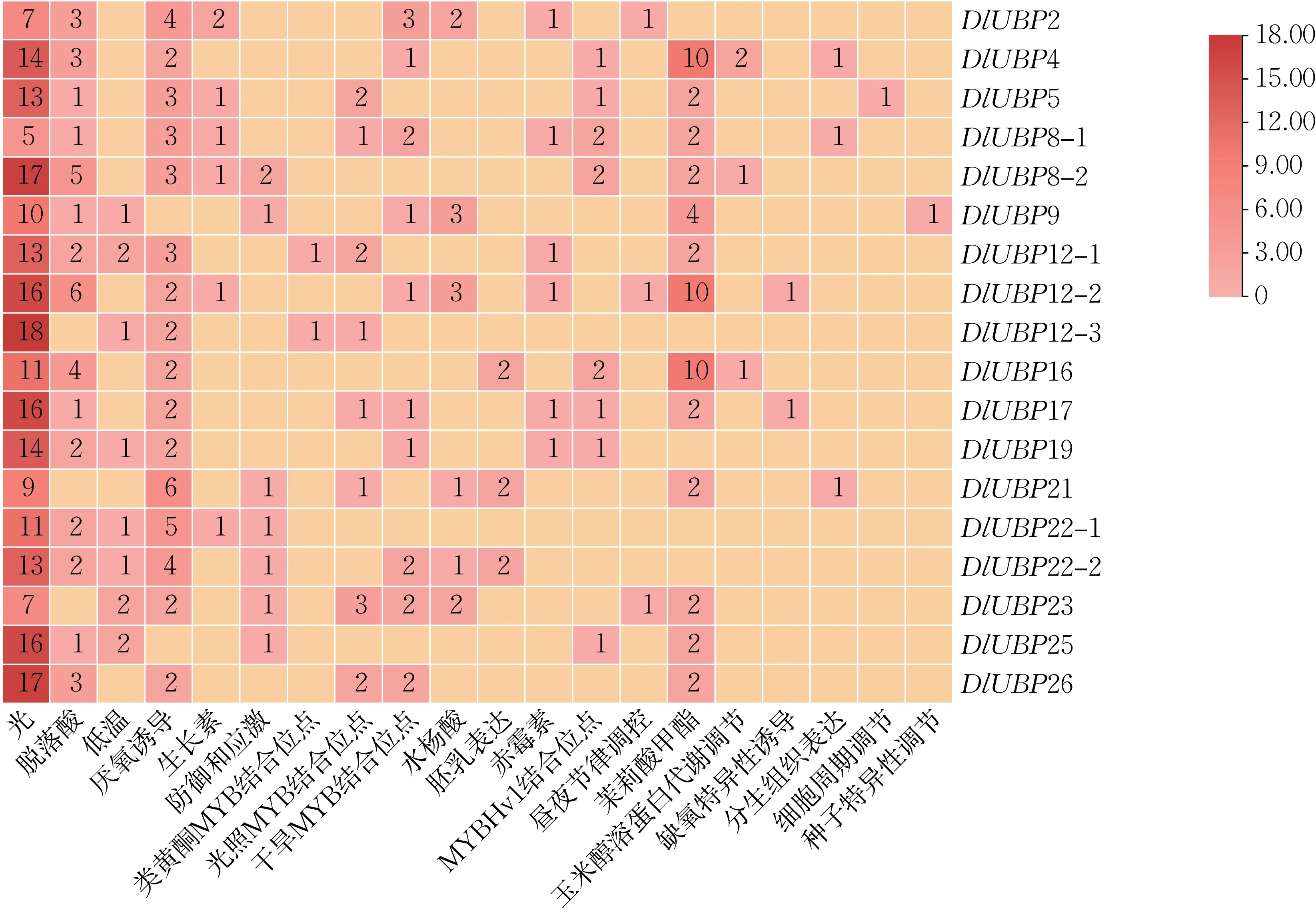
图5 DlUBP 基因启动子顺式作用元件Fig.5 Promoter cis-acting elements of DlUBP gene
2.6 DlUBP 基因在龙眼体胚发育过程中的表达分析
利用龙眼基因组数据库中DlUBP 基因在龙眼体胚发生过程中EC、ICpEC、GE 3 个阶段的特异表达FPKM 值绘制聚类分析热图(图6),发现DlUBP基因在体胚发生阶段的表达模式共有6 种:(1)在EC 和GE 阶段表达下调,在ICpEC 阶段上调(DlUBP2、DlUBP5、DlUBP8-1、DlUBP8-2、DlUBP9、DlUBP12-1、DlUBP12-2、DlUBP17、DlUBP26);(2)在ICpEC 和GE 阶段上调,在EC 阶段下调(DlUBP22-2、DlUBP19、DlUBP23);(3)在EC 和ICpEC 阶段上调,在GE 阶段下调(DlUBP12-3、DlUBP21、DlUBP25);(4)在EC 阶段上调,在ICpEC 和GE 阶段下调(DlUBP22-1);(5)在EC 和GE 阶段上调,在ICpEC 阶段下调(DlUBP4);(6)在EC 和 ICpEC 阶段下调,在 GE 阶段上调(DlUBP16)。整体来看,多数DlUBP 基因的表达在EC 阶段和GE 阶段下调,在ICpEC 阶段上调,推测DlUBP 基因可能有助于龙眼胚性愈伤组织的分化。
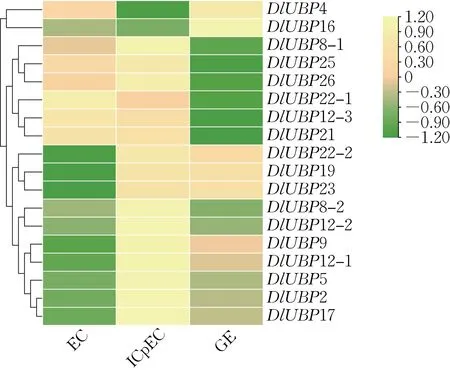
图6 DlUBP 基因在龙眼体胚发生各阶段的特异表达Fig.6 The specific expression of different stages of somatic embryogenesis in the DlUBP gene
2.7 DlUBP 基因在脱落酸和干旱胁迫处理下的表达量分析
DlUBP 基因启动子中含有较多的脱落酸顺式作用元件,为探究DlUBP 基因在脱落酸处理下的表达情况,筛选含脱落酸响应元件的6 个基因:DlUBP2、DlUBP8-1、DlUBP12-1、DlUBP12-2、DlUBP17、DlUBP19,采用qPCR 方法分析它们在不同时间梯度(0,4,8,12 h)脱落酸处理下的表达量,如图7 所示。可知,DlUBP2、DlUBP12-1、DlUBP17基因在脱落酸处理下表达量先升高后降低,处理4 h时表达量达最高;DlUBP8-1 基因在脱落酸处理下的表达量明显降低,均低于0 h 对照组;DlUBP19 基因的表达量随脱落酸处理时间的增加逐渐升高,在处理12 h 时表达量急剧升高;DlUBP12-2 基因的表达量在脱落酸处理4 h 时无明显变化,在处理8 h 达到最高,然后显著下降,在处理12 h 时最低,且低于0 h 对照组。综上所述,脱落酸能影响DlUBP 基因的表达,且多数DlUBP 基因的表达量在处理4 h 时较高,在处理12 h 时最低,猜测脱落酸处理时间较长会抑制多数DlUBP 基因的表达。

图7 6 个DlUBP 基因家族成员在不同脱落酸处理时间下的表达情况Fig.7 Expression of six DlUBP gene family members under different abscisic acid treatment times
为探索DlUBP 基因在干旱胁迫处理下的表达水平,从18 个DlUBP 基因中随机挑取含干旱胁迫响应元件的6 个基因:DlUBP2、DlUBP8-1、DlUBP12-2、DlUBP17、DlUBP22-2、DlUBP23,用qPCR 方法分析它们在10%聚乙二醇(PEG6000)渗透调节剂模拟的不同时间梯度(0,4,8,12 h)干旱胁迫处理下的表达量,结果如图8 所示。可知,随着处理时间的增加,DlUBP2、DlUBP22-2、DlUBP23 基因的表达量先降低后升高;DlUBP12-2 基因的表达量先降低再升高再降低,在处理8 h 时表达量最高。4 个基因的表达量均在处理4 h 时达最低水平,低于0 h 对照组,推测干旱胁迫对这4 个基因的表达产生影响,且在处理4 h 时抑制其表达,在处理8 和12 h时促进其表达(DlUBP23 的表达在处理8 h 时被抑制)。此外,在干旱胁迫处理下DlUBP17 的表达量均高于0 h 对照组,但在处理8 h 后即使增加处理时间,表达量的升高幅度也非常小,推测干旱胁迫对DlUBP17 的表达有促进作用,且表达量可能在处理8 h 后逐渐趋于稳定。结果表明,干旱胁迫处理影响DlUBP 基因的表达,且处理时间不同,对表达量的影响亦不同。
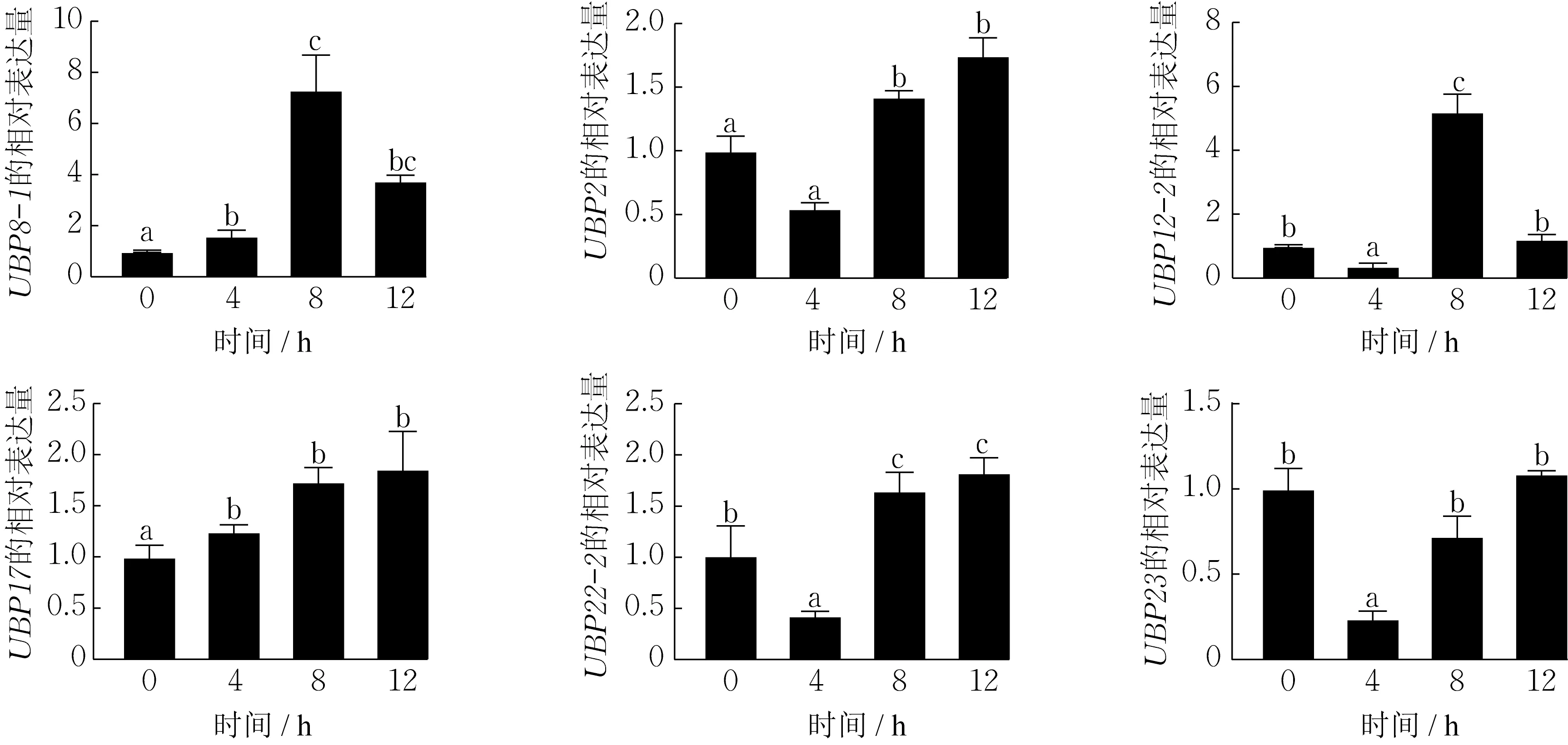
图8 6 个DlUBP 基因家族成员在不同干旱胁迫处理时间下的表达情况Fig.8 Expression of six DlUBP gene family members under different drought stress treatment times
3 讨论
3.1 DlUBP 基因的功能可能具有多样性
去泛素化酶广泛存在于真核生物体内,它们在细胞内分工明确,同时相互协调,共同维护细胞内泛素化水平的稳定[19]。目前对于去泛素化酶功能的研究尚不够深入,已知的生理功能主要有激素调控与应激反应[20]、肿瘤与癌症的发生[21]等,在植物体内主要调控植物的形态发生、免疫应激和信号转导和程序性细胞死亡等,在动物体内参与癌症与肿瘤[22]的发生、神经和生殖细胞分化、DNA 损伤修复等[23]。泛素特异蛋白酶是去泛素化酶的一种,其生物学功能与去泛素化酶相似,可调节植物生长发育和对环境响应,并介导多种信号转导途径。参照拟南芥的基因分类鉴定方法,对龙眼、拟南芥、甜橙、水稻4 个物种进行进化树分析,将基因分为14类。龙眼中有18 个DlUBP 基因,拟南芥中有27 个AtUBP 基因,由于龙眼与拟南芥的亲缘关系较近,猜测其生物学功能也可能较相似[24]。DOELLING等[25]的研究指出,缺乏AtUBP14 基因会阻止拟南芥胚胎的发育甚至导致胚胎死亡,AtUBP13 基因在调节根分生组织发育中起关键作用[26]。此外,AtUBP2 可能参与了体内异常氨基酸的代谢,AtUBP12 能调节开花时间,AtUBP26 在种子发育过程和花期中起重要调控作用等。龙眼中也有相同的UBP 基因,且其蛋白质结构的相似程度较高,如DlUBP2 和AtUBP2 都有1 个ZnF-UBP 锌指结构,DlUBP12 和AtUBP12 都有1 个MATH 结构域,DlUBP26 和AtUBP26 都有1 个DUSP 结构域,因此推测龙眼中的UBP 基因有类似的功能。LEE等[27]通过分析小鼠胚胎阶段的Northern 印迹发现,USP22 在多数发育的小鼠胚胎中普遍表达,且在各种成人组织和胚胎早期均有广泛表达,于是猜测其与胚胎发生有关。不仅如此,分析发现,DlUBP基因启动子含有20 余种顺式作用元件,包括光响应元件、厌氧诱导响应元件、缺氧特异性诱导元件、干旱胁迫响应元件、多种激素响应元件等,且不同的DlUBP 基因所含的顺式元件的种类和数量不同。DlUBP12-1 和DlUBP12-3 基因含有参与类黄酮生物合成基因调控的MYB 结合位点,表明DlUBP 基因可能参与龙眼体胚发生过程中花青素、黄酮类化合物等次生代谢产物的合成[28];DlUBP5 基因含有细胞周期调控元件,猜测DlUBP基因可能与龙眼体胚细胞周期的调控有关。总之,18 个DlUBP 基因在功能上既有特异性也有互补性,共同维护龙眼正常生长发育并提高其抵御逆境胁迫的能力。
3.2 DlUBP 基因可能参与脱落酸的调控并对干旱胁迫有应激反应
组织特异性和应激反应基因表达情况主要取决于启动子中的顺式元件,顺式元件与多种应激反应基因密切相关。分析发现,DlUBP 基因启动子含有多种植物激素的响应元件,如脱落酸响应元件、赤霉素响应元件、茉莉酸甲酯响应元件、水杨酸响应元件等,这表明龙眼中的DlUBP 基因可能在多种激素胁迫响应中起作用。MOON 等[13]指出,毛竹中含有与脱落酸对应的顺式元件,同时证明了有些PeUBP 基因具有较高的脱落酸敏感性,推测PeUBP 基因在植物发育和脱落酸胁迫响应中发挥了作用。在本研究中,随着脱落酸处理时间的增加,6 个DlUBP 基因中的5 个表达量呈下降趋势,有些甚至低于0 h 对照组,仅DlUBP19 的表达量逐渐上升,由此推测DlUBP 基因可能参与脱落酸的调控,且基因表达情况与处理时间有很大关系。
在胁迫条件下,脱落酸快速产生,调节气孔关闭,影响胁迫响应基因的表达。JINFENG 等[29]指出,AtUBP24 是拟南芥中脱落酸信号传导和盐胁迫耐受性的负调控因子,若缺少AtUBP24 基因,可能导致拟南芥在脱落酸诱导的气孔调节中对干旱胁迫的敏感性增加。不仅如此,文献[30-31]指出,在干旱胁迫下,MYB 受脱落酸依赖,可通过调节胁迫增加植物的耐旱性[30]。对龙眼DlUBP 基因启动子的分析发现,50%以上的DlUBP 家族成员启动子含有在干旱胁迫下MYB 转录因子结合位点,且其中一半成员具有1 个以上该结合位点,初步猜测DlUBP 基因可能对干旱胁迫有响应。通过分析干旱胁迫处理不同时间梯度下6 个DlUBP 基因的表达量发现,在干旱胁迫处理不同时间后,6 个DlUBP基因的表达量均呈上升或下降趋势,大部分基因的表达量在干旱胁迫处理4 h 后呈上升趋势,因此推测DlUBP 家族对干旱胁迫有应激反应,且适当时间的干旱胁迫处理有利于基因表达。
4 结论
对龙眼泛素特异蛋白酶(UBP)进行全基因组鉴定,对体胚早期不同阶段以及在外源胁迫下的表达进行了分析。在龙眼中共鉴定出18 个DlUBP 基因,分析了DlUBP 基因的结构、蛋白质理化性质、染色体定位、启动子顺式元件、在不同时间梯度的脱落酸处理和干旱胁迫处理下的表达情况。结果表明,DlUBP 基因可能参与龙眼体细胞胚胎发育、脱落酸和干旱胁迫响应过程,可为后续对龙眼UBP 家族单基因的功能验证以及外源胁迫的响应机理研究提供参考。
猜你喜欢
作文大王·低年级(2019年4期)2019-05-13 01:44:10
临床医药文献杂志(电子版)(2017年77期)2017-08-15 00:55:30
岭南音乐(2016年5期)2017-01-17 07:44:58
学苑创造·A版(2016年9期)2016-10-10 11:36:01
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15 12:47:42
军事文摘·科学少年(2015年6期)2015-05-30 20:39:04
军事文摘(2015年12期)2015-05-07 12:52:26
中国医学科学院学报(2015年5期)2015-03-01 04:03:46
现代检验医学杂志(2015年2期)2015-02-06 02:01:01
四川生理科学杂志(2014年3期)2014-02-28 14:09:38
