
功能磁共振成像技术在织物刺激脑感知中的研究进展
2023-05-30 05:36翟淑娜娄琳王其才苑洁
现代纺织技术 2023年3期
翟淑娜 娄琳 王其才 苑洁


摘 要:针对织物舒适度领域中大脑感知机制尚不明确的问题,功能磁共振成像技术凭借其超高的空间分辨率在织物刺激脑感知表征领域表现出良好的技术优势。根据织物刺激的感知过程以及功能磁共振成像技术的表征原理,总结来自织物触觉刺激、视觉刺激、视-触觉跨模式刺激的大脑感知规律,并提出将此技术深入运用于脑感知表征研究时需要突破的一些难题和方向,期待以其客观、即时的优势为构建织物舒适度脑感知理论体系和满足纺织服装产品设计舒适性要求提供新思路与新方法。
关键词:舒适度;脑感知;功能磁共振成像;触觉;视觉
中图分类号:TS941.19
文献标志码:A
文章编号:1009-265X(2023)03-0274-11
基金项目:国家自然科学基金项目(52003245);浙江省自然科学基金项目(LQ18E030007);先进纺织材料与制备技术教育部重点实验室和浙江省纤维材料和加工技术研究重点实验室(浙江理工大学)开放基金项目(2019QN05);浙江省教育厅一般科研项目项目(113129A4F21075);浙江理工大学科研启动基金项目(11313132612042)
作者简介:翟淑娜(1997—),女,山东临沂人,硕士研究生,主要从事织物压力舒适度脑感知方面的研究。
通信作者:苑洁,E-mail:yuanjie@zstu.edu.cn
关于织物舒适度的表征技术,传统的心理评价虽然简便易行,但存在个体差异、随机性、不稳定性、不能实现感知评价定量化的缺陷。而以物理机械信号与主观评价规律形成对织物舒适度的物理评价,虽以数据为基础,但评价的对象却是织物,而非感知的主体——人体。随着医学生物技术的不断发展和跨学科渗透,生理学评价应运而生。在织物多种刺激作用下,对人体的心率[1]、皮肤温度[2]、血流量[3]、等生理特征与人体舒适度感知的关系进行了大量的研究。其中,脑电图[4](Electroencephalography,EEG)、事件相关电位技术[5](Event-related potentials,ERPs)和功能磁共振成像技术[6](functional Megnetic resonace imaging,fMRI)均可用于探究在织物舒适度的脑感知研究。EEG的某些节律可以有效评价人体穿着纺织品的舒适性狀态,例如α 波比例差值与织物的舒适感知呈正相关[7],而 β 波、θ波、δ波与大脑的紧张不适感知有关[4,8]。除此之外,基于EEG 技术的丝织物触感试验证明,β波越大,织物的接触冷暖感越强,而θ、α波越大,刺痒感越小[9]。在ERPs技术监测下,诱发电位成分P50、P100、P200、P300可作为脑生理反应指标表征织物轻微触感刺激[5, 10-13],P300、N450表征织物接触压力刺激[14],P100、N100、P200、N200、P300表征织物视觉刺激[15-18]。两种技术在研究中都表现出了百微秒级超高时间分辨率的优势,但由于受限于颅骨不匀且有个体差异、容积导体效应以及脑电源间电场开放性的差异,其空间分辨率均较低,远不及fMRI技术,只是定性地表达织物舒适程度,尚未实现舒适度表征的定量化。
相较而言,fMRI技术是目前可实现织物舒适度原位表征的最优方法。故此,本文以fMRI技术的表征原理为基础,从织物触觉刺激、视觉刺激、视觉-触觉跨刺激3个领域出发,综述fMRI技术的织物刺激脑感知研究现状,提出此技术目前需要突破的方向,期待以其客观、精准的优势为织物舒适度脑感知理论体系和评价标准的研究突破提供新的契机。
1 fMRI技术表征原理
fMRI技术可以对织物触觉、视觉、视觉-触觉跨模式刺激相关的激活脑区进行有效的实时监测,可以精准地捕捉刺激过程中人体产生的心理、生理变化。基于血氧水平依赖信号(Blood oxygenation level dependent,BOLD)的功能磁共振成像技术是以新陈代谢的血液动力学方法为基础的[19]。大脑在各种不同刺激下某区域的神经激活时,局部脑组织血流量、血流容积以及血氧消耗均会增加,但大脑中的血氧消耗量仅轻微增加,导致血流量增加程度超出了氧耗量的增加程度,最终导致脑激活功能区的静脉血氧浓度升高[20],所以氧合血红蛋白(逆磁性物质)含量增加,脱氧血红蛋白(顺磁性物质)相对减少,从而表现为弱信号,其浓度越低,宏观磁共振信号强度降低幅度越大,而组织的磁共振信号强度则会上升,从而产生BOLD效应,BOLD信号增加,激活脑区最终在磁共振图像上呈现为亮信号[21-22]。基于fMRI的织物刺激脑感知表征原理如图1所示。
首先,常用的fMRI实验序列由任务刺激状态和静息状态两部分组成,在设计的实验序列中,人体感官受到织物材料或单纤维的触觉和视觉单模式以及视-触觉跨模式这类特定刺激后,各效应器接收刺激信号经脊神经传入脊髓后到达大脑皮层中的神经细胞,神经系统会对织物材料的各种刺激产生大脑反应的BOLD,由1.5 T至3 T的标准头颅线圈医用磁共振系统监测特定磁场中的脉冲信号变化,并采集受试者的脑部结构图像和功能图像[21-22]。
其次,在数据预处理阶段,利用以MATLAB为载体的SPM12(Statistical parametric mapping)插件,其可以完成 fMRI 数据的预处理、统计分析、结果报表及可视化查看、计算大脑特定功能区的功能连接,以及数据处理效果检查等其他功能。MRIConvert可以将原始的 DICOM 格式转化成SPM 软件可使用的 NIFTI 格式,DPARSF可以进行数据预处理的批量操作。具体操作为,首先将采集到的功能成像进行头动校正,利用最小二乘法原理和六参数包括三维平移和三维旋转刚体模型的空间变换,即:先把时间序列内的除第一组全脑图像之外的其他全脑图像都通过匹配算法做刚性旋转到第一组全脑图像的位置,后用内插算法来对每个像素进行重采样[23]。其次将经过头动校正的平均大脑进行标准化,将平均大脑与结构像配准,实现功能图像与结构图像的空间对齐,得到校正后的完整大脑成像;考虑到人体大脑的个体差异性,将所有被试的结构大脑均先分割成灰质、白质和脑脊液,生成灰质和白质图像,过程中将被试的大脑结构空间与标准大脑空间进行对应,得到两个大脑结构空间相互转换的矩阵,利用转化矩阵,将被试的功能图像再配准到标准空间,转换成标准大脑,实现功能图像的标准化以便进行成像比对,为保证结果的准确性,在分割和标准化过程中使用基于李代数的微分解剖配准方法[24]。最后利用高斯核滤波器对三维的功能图像卷积,进行空间平滑[25],抑制生理噪声,完成脑成像的预处理。在统计分析阶段,根据实验设计的刺激方案建立一般线性模型,如式1所示,对模型中的参数即回归系数β用约束最大似然法进行估计,如果回归系数β显著不为零,则表明此体素的时间序列与该刺激任务显著相关,将以激活的形式显示到最终的大脑反应结果图中。即,用被试在刺激任务阶段与静息阶段的BOLD 信号相减,得到被试受刺激时大脑反应的变化情况,给其设定合适的激活阈值,得到最终的大脑激活结果。后推断与刺激相关的大脑结构和功能区信息。再对多个被试的数据结果进行多种方法的统计分析处理,如单样本t检验、双样本t检验、方差分析、多元回归分析、全因子分析等,后进行多重比较校验,其中最常用的就是FDR(False discovery rate)和FWE(Family wise error)[23]。
式中:Y指实验中利用 fMRI 同步扫描被试的大脑,检测到的被试真实的BOLD信号;X指根据BOLD原理假设的由于被试在实验中受到刺激而产生的理想的BOLD信号,是实验中操作的时间点序列与被试大脑的血液动力学响应函数进行卷积之后产生的矩阵; ε指误差。
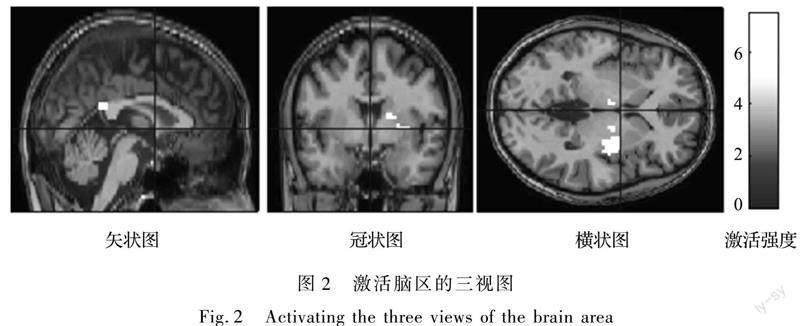
最后,在数据可视化阶段,可使用直接基于 Windows 操作系统的单独的软件程序如:DPABI(Data processing & analysis of brain imaging)、Xjview、Marsbar以及Anatomy 等软件对大脑成像进行可视化操作,通过探索刺激信号与脑激活信号之间的变化规律完成织物舒适度的大脑原位感知表征。数据表达形式多样化,不仅可通过脑成像激活切片图、激活切片三视图(如图2所示)、三维立体脑图(如图1右下角所示)直观多面显示观测,而且还可精确计算大脑响应位置、激活强度、激活占比以及脑区间功能连接协作情况[26]等一系列变化。正激活是指执行刺激任务时该脑区内相对于静息状态的局域脑血流升高,局域脑受到神经促进,表达为感觉舒适。刺激作用下的BOLD信号小于静息状态下的BOLD信号,供氧与耗氧不平衡,表现为负激活,代表感觉不舒适。激活强度越大,代表激活脑区的舒适或不舒适感觉越强;激活占比越大,代表激活脑区的范围越大,功能连接越强;功能连接越强,代表激活脑区之间的联系越密切[6]。SPM激活报表法确定激活点峰值强度以及激活体素个数,自动解剖标记图谱法(Automated anatomical labeling,AAL)[27-28]显示各激活点解剖学结构定位,利用DPABI将峰值点坐标输入,即可显示该点所在的确切功能脑区定位,Xjview可以进一步明确该激活团块所在蒙版脑区定位,这些软件均可以具体了解、检查和修正激活点功能区位置信息,Marsbar和Anatomy可以进行感兴趣区分析来表征大脑注意力和織物特定刺激之间的相关关系。图1中右下角图是以人体腋中线为中线将脑区分为前侧的腹侧面和后侧的背侧面。由上至下由左至右依次分别为左脑外侧、脑背侧、右脑外侧、左脑内侧、脑腹侧、右脑内侧、脑前侧、脑后侧,实现多维显示脑区激活情况。图2以大脑中前与中正交叉点为坐标原点,从大脑左侧向右侧延伸为X方向,由大脑后侧向前侧延伸为Y方向,由大脑下侧至上侧为Z方向建立三维坐标轴,将人体大脑定位[29]。图2从左至右依次为从左向右侧投射的大脑矢状图、从后向前投射的冠状图、从上向下投射的横状图,同样以不同色度代表激活强度,可以从3个方向直观定位同一脑区的激活情况[6]。
2 基于fMRI的织物单模式刺激脑感知研究
目前,基于fMRI技术的材料或织物刺激主要涉及触觉刺激、视觉刺激和视觉-触觉跨模式刺激3个方面。
2.1 基于fMRI的织物触觉刺激脑感知研究
根据织物接触人体皮肤作用面积和作用力的不同,来自织物触觉的刺激可划分为轻微触感刺激和接触压力刺激两类。织物轻微触感舒适度是指皮肤受到外加织物的微力和点作用时,织物表面性能对人体心理生理产生的触感[30]。织物接触压力舒适度是指织物包覆人体局部或整体皮肤面积,在作用时间和压力后产生的舒适感觉范围,主要表达对皮肤所涉及的神经系统和血液循环系统挤、压、阻碍所产生的不适感[31]。另外基于fMRI的触觉短时记忆(Tactile short-term memory,TSTM)研究也是探索织物触觉刺激的一个重要角度。
2.1.1 基于fMRI的织物轻微触感刺激脑感知研究
基于fMRI的轻微触感刺激脑感知研究主要涉及粗糙感、黏着感、柔软感和刺痒感,主要会引起躯体感觉皮层和运动皮层的激活以及脑区间功能连接的变化。
关于粗糙感,Wang等[27-28]运用SPM12和MRIcro软件进行数据预处理,利用AAL和布罗德曼功能图(Brodmann′s area maps,BA)对手指主动触摸丝绸和亚麻织物时的激活脑区进行研究。所谓布罗德曼功能图,是1909年德国神经学家Korbinian Brodmann使用尼氏细胞染色法得到的以神经元细胞结构组织为基础的大脑分区,不同的脑区对应不同的大脑机能,分别用数字表示,共划分为52个脑区[32]。研究结果表明不同粗糙度的织物接触刺激会激活不同的脑区,主要分布在躯体感觉皮层,躯体感觉皮层包括初级感觉皮层(Primary somatosensory cortex,SI)和次级感觉皮层(Secondary somatosensory cortex,SII)。其中,BA3区、BA1区、BA2区、AAL81区属于SI,BA40区、BA43区、BA48区、AAL92区属于SII。亚麻触觉刺激引起的AAL81、BA1、BA40区的最大激活强度均大于丝绸,而丝绸触觉刺激引起的BA2区和BA3区的激活强度和激活占比均大于亚麻,说明前者与接触粗糙感有关,而后者与接触光滑感有关。Yuan等[33]运用Marsbar 和Anatomy软件计算脑区的信号变化百分比 (Percent signal change, PSC)表征大脑注意力与织物表面粗糙度之间的相关关系,PSC越大,代表该脑区对该刺激的关注度越。随着无骨紧身胸衣的表面逐渐由光滑到中等粗糙再转变为粗糙,出现最大PSC的脑区逐渐从SI 中的BA3区过渡到BA1区,最终转移至SII中的深层皮质顶叶岛盖。此外,对天鹅绒的实际触感和手感错觉都会激活SI[34],且BA3区的激活强度与错觉强度呈正相关,验证了SI与光滑感知有关。而触摸舒适感差的平角纹理会同时引起SI、SII和运动皮层的负激活[11],说明接触粗糙感知与SI、SII和运动皮层有关。除了脑区的激活外,触觉刺激也会引起脑区之间功能连接的变化。Rajaei 等[35]运用SPM12和CONN软件通过心理生理交互作用分析方法发现触觉错觉强度会调节左侧SI中的BA3区与SII中顶叶盖、顶上小叶、中央前回、脑岛和小脑这类高阶体感皮质之间的功能连接。
关于黏着感,文献[27-28]研究发现黏着感织物触觉刺激时,运动皮层中的BA6区和AAL83区和感觉皮层中的BA1、BA43、BA48区会出现不同程度的正激活,而且织物黏着表面接触刺激会使中央后回、中央前回、顶内沟等躯体感觉皮层和辅助运动脑区产生一定程度的功能连接[36]。Yeon等[30]发现黏着感会激活同侧背外侧前额叶皮层和区分黏性细微知觉的大脑区域,主要包括皮质区域和皮质下区域,且与黏性强度呈正相关。即,SI中的 BA1区、SII 中的BA43区和BA48区、运动皮层中的BA6和AAL83以及辅助运动区与黏着感相关。
关于柔软感,Wang等[37-38]发现用无毛皮肤主动触摸柔软的丝绸时,SI、SII和運动皮层均出现大量激活,且最大激活脑区位于左脑SI的BA2区,用有毛皮肤主动触摸柔软的丝绸时,最大激活脑区位于后岛叶,这可能与岛叶的AAL60反映触摸皮肤引发的愉快情绪高度相关[27]。另外Wang等[39]还运用Marsbar 和Anatomy对比研究用手指主动接触两种丝绸和亚麻所产生的PSC,发现了在触摸丝绸时SI和顶叶岛盖的PSC明显高于触摸亚麻的PSC。2019年,Kitada等[40]也发现顶叶盖、岛叶的激活与材料柔软度呈正相关,且这些脑区与SI、顶后小叶和枕叶皮质构成了触觉柔软知觉大脑网络的重要节点。即,SI、SII产生更多的感官或辨别信息,岛叶则处理情感触摸信息。
关于刺痒感,Wang等[41]在运用单纤维刺激前臂掌侧产生刺痒感的研究中发现,SI、SII、运动皮层、岛叶、边缘叶、视觉皮层、听觉皮层均有明显的激活,其中SI、视听觉皮层中的梭状回和运动皮层中的小脑激活程度较高,这可能与单纤维刺痒刺激会使受试者回忆痛感并控制身体运动有关。
总结以上研究发现,SI参与触觉信息处理,虽与薄感、柔软感、光滑感、粗糙感、黏着感等都有关,但按特征脑区激活强度和激活占比的大小,SI更与光滑感、柔软感紧密相关,另外SI与顶叶盖、顶上小叶、中央前回、脑岛、丘脑、边缘上回这类高阶体感皮质相互作用实现有意识纹理触觉感知,是材料表面触感的下方大脑网络关键节点。SII参与触觉加工并产生感知或辨别性知觉,SII和运动皮层与粗糙感、黏着感、表面纹理精细感相关。而刺痒感激活躯体感觉皮层、运动皮层、情绪脑区、视觉听觉皮层,属于一种多维触感。
2.1.2 基于fMRI的织物接触压力刺激脑感知研究
基于fMRI的织物接触压力刺激脑感知研究主要是探索感知织物渐增压力的特征脑区方面。
在感知织物压力舒适度的特征脑区方面,Yuan等[42]运用SPM12、DPABI、Xjview软件分析数据,对人体腰腹部施加渐增织物压力刺激时发现,织物舒适接触压力刺激时,右侧SII脑血流升高,局域脑受到神经促进,感觉舒适,出现正激活,且脑岛与楔前叶的功能连接增强,舒适感知增加;而低于和高于织物舒适压时,SI和杏仁核脑区都会产生负激活,不同的是前者耗氧量过大,对SI的神经抑制作用明显,后者造成供氧量不足,对杏仁核的神经抑制作用更为显著,即随压力增加,负激活强度最大的脑区从SI转移到杏仁核,且负激活范围增加,负激活强度降低。而在脑区所含神经方面,右侧SII抑制疼痛,并且有毛皮肤的舒适感知由A-β传入神经兴奋性在躯体感觉皮层处理;SI反映疼痛,过于轻微的织物触压也会在大脑中产生不适感,这种不适感很可能是由织物表面的宏观几何特性引起的;杏仁核含有疼痛情绪神经元,参与疼痛感知,SI和SII伤害性信息都在杏仁核处理结束。
总结发现,右侧SII是织物舒适压力的感知脑区,SI对面料宏观表面触觉感知具有显著向下调节作用,杏仁核是与织物压迫知觉相关的特征脑区。总之,躯体感觉皮层、运动皮层和情绪功能区相互作用形成触觉刺激信息。
2.1.3 基于fMRI的织物触觉短时记忆脑感知研究
TSTM是将感知到的有限数量的触觉刺激在短时间内保留下来的一种能力,决定人对一定数量触觉信息的准确感知[43]。基于fMRI的织物触觉短时记忆脑感知研究较少,研究主要集中在探索振动TSTM特性以及短时记忆对触觉感知的影响。
童新宇等[43]发现手指振动触觉记忆容量略高于手腕部振动触觉记忆容量,但低于视觉记忆容量,振动强度离散间隔越大,则振动触觉记忆容量越大,振动持续时间过长或过短都会导致振动记忆容量减少,男性最佳持续时间为400 ms,女性为 300 ms,振动刺激序列按强度递增或递减的顺序排列比混乱顺序时更容易记忆,振动刺激系列越长,再认正确率越低,反应时间越长。Zhang等[44]采用主观评价法对11种面料的厚度进行触觉感官评价序列实验,发现在感官评价中存在STM效应,导致幻觉记忆差异,即前一种织物的触觉感觉会直接影响对下一种织物的感觉,连续实验的时间间隔越短,影响越显著。因此在感官评价中,适当的时间间隔可以抵消STM效应。在感知脑区对服装压力的适应性方面,Chung等[45]在观测服装压力刺激持续时间与感知脑区反应之间的关系中发现,对侧SI和SII激活程度随时间呈指数下降,脑半球间SI和SII、同侧SII、对侧SI之间的功能连接线性降低,而对侧SI中BA3和躯体感觉联合皮质中的BA5区之间的功能连接线性增加。即,在处理有效触觉信息时,皮层的激活和脑区间的功能连接会发生动态适应性变化。
总结以上研究发现,刺激部位、性别、振动强度、振动持续时间、振动刺激序列都对振动触觉记忆容量有影响,而且SI和SII之间的功能连接程度变化与持续时间的压力刺激有关,因此,在刺激过程中,我们掌握并有效控制这些影响因素可实现控制STM效应。
2.2 基于fMRI的织物视觉刺激脑感知研究
基于fMRI的织物视觉刺激脑感研究主要集中于感知材料属性和判断材质类别以及有关视觉短时记忆(Visual short-term memory,VSTM)研究。
关于感知材料属性,视觉材料属性如光泽、纹理、重量、形状和硬度等。在光泽方面,与哑光相比,初级视觉皮层、次级视觉皮层和腹侧视觉皮层的梭状回和背侧视觉皮层对有光泽的物体反应更敏感。在纹理和形状方面,腹侧视觉皮层内侧和外侧区域并行处理材料的自然纹理和形状信息[46],进行织物纹理视觉感知时,沿内侧腹侧视觉皮层的侧沟、海马旁回、海马旁区域、舌回,枕内侧皮层,顶上叶,内侧颞叶,外侧前额叶皮层,楔前叶都对纹理信息感知敏感[47],感知形状差异的脑区主要位于外侧腹侧视觉皮层如枕外侧复合区(Lateral occipital complex,LOC)[48]。其中,LOC的梭状回可以综合表示自然纹理、颜色和形状的信息,其中每一个信息都是围绕其单独分析。在硬度方面,感知材料硬度的脑区主要位于侧沟、舌回、双侧枕颞皮层。
关于判断材质类别,感知不同的视觉材料属性倾向于每个材料类别聚集。对判断材料类别已经从早期视觉区域的图像表征转移到梭状回及其周围区域这类高级视觉区域的知觉表征。Suzuki等[49]发现一名左脑腹内侧枕颞叶皮层受损的患者在匹配和命名材料纹理信息遇到困难,这可能与腹内侧枕颞叶皮层中的梭状回及其周围区域,比如舌回、侧沟、海马旁回等,对材料类别的感知较为敏感有关[50]。
关于视觉短时记忆,一些研究也阐明VSTM与材料类别相关的大脑机制。VSTM大约有4个物体的固定容量,Alvarez 等[51]发现VSTM容量极限是由材料数量和视觉信息负荷共同决定,顶内沟和枕内沟是VSTM存储容量的关键区域[52],两个区域中与容量有关的激活主要与VSTM中编码对象的空间位置密切相关。其中,下顶内沟占72%~87%左右,且随视觉记忆负荷的增加,右脑下顶内沟反映VSTM负荷,与视野位置无关[53]。Xu等[54]发现在编码和记忆维持中材料颜色的VSTM与外侧枕叶皮层和上顶内沟有关。Otsuka等[55]利用感兴趣区方法评估材料类别短时记忆的神经反应,并计算上顶内沟和对材料类别感知敏感的梭状回的平均信号变化,使用基于图像和基于触摸的两种变化检测任务来测量材料类别的VSTM,使用相关分析和来自主扫描的数据进行全脑分析来检验两个区域的脑激活以及与其他大脑区域之间的关联,研究发现梭状回的信号变化百分比在视觉刺激和触觉刺激任务中存在差异,说明梭状回可能参与不同材质在视觉刺激和触觉刺激之间差异的VSTM。而最近VSTM在预测理论的背景下被提及,Li等[56]在使用重复抑制范式测量视觉联合皮层中梭状回的BOLD信号时发现,当VSTM过程中,梭状回的重复抑制效应显著减弱,说明VSTM与梭状回脑区也存在一定关联。
总结以上研究发现,关于材料的视觉信息主要通过腹侧视觉皮层的层次结构来处理和表征。其中,区分材料图像信息在初级视觉区域中表示,而高级视觉区域反映感知材料属性和材质类别差异。对于视觉短时记忆,顶内沟和枕内沟是VSTM存储容量的关键区域,梭状回参与材料在视觉和触觉刺激之间差异的VSTM。
2.3 基于fMRI的织物视-触觉跨模式刺激脑感知研究
视-触觉跨模式脑感知,是指通过视觉或触觉两种感官形式获得的信息在大脑知觉中紧密联系比如感知材料属性或类别脑区的多感官表征,视-触觉信息交互影响等[57]。关于视-触觉织物的跨模式脑感知研究主要在于感知材料属性脑区多感官表征,视-触觉信息的相互影响和视-触觉短时记忆3个方面。
在感知材料属性脑区的多感官表征方面,如纹理、形状、硬度等。在纹理、重量和形状方面,在触觉感知织物粗糙度时,沿内侧腹侧视觉皮层的侧沟、海马旁回、海马旁区域、舌回,枕内侧皮层,顶上叶,内侧颞叶,外侧前额叶皮层,楔前叶同样激活,这表明纹理特征识别表征有跨皮质区域网络分布,其中外侧前额叶皮层和内侧颞叶之间有自上而下的功能连接[58],这表现出顶上叶,内侧颞叶,前额叶皮层,楔前叶和腹侧视觉皮层大脑网络具备处理材料特性的多感官表征;而且通过触摸材料纹理来视觉判断材料重量属性时,腹侧视觉皮层的侧沟激活[59],这表明腹侧视觉皮层通过多感官交叉刺激已经开始代表反映重量这类非视觉属性。Newman等[60]研究发现盲人触摸材料形状会激活LOC,而且不管盲人曾触摸还是明视人曾看过材料形状,左侧LOC都激活,这表明该脑区不仅是形状选择区,同时还与材料属性的回忆有关。其中,LOC的梭状回可以综合表示自然纹理、颜色和形状的信息。在硬度方面,感知材料硬度的脑区还包括左侧顶内沟,其中观看左手触摸材料硬度的镜面反射激活的双侧枕颞皮质,可能与右手触觉感知材料硬度時左前顶内沟的输入信息有关[61]。
在视觉与触觉信息的相互影响方面,首先视觉刺激对触觉反应的影响,在观看不同面料照片时,刺激中存在的表面属性如颜色梯度、柔软度、纹理会影响触觉刺激的大脑匹配方式[62]。剥夺视力2 h后进行视觉感知材料形状时会显著改变形状触觉知觉的神经加工,纹外视皮层和顶内沟的激活和两个脑区间功能连接都降低[63]。其次触觉刺激对视觉反应的影响,O'callashan等[64]发现跨模式训练即先对受试者进行织物纹理的触觉感知训练再进行视觉感知纹理测试,左内侧枕叶皮质的激活程度比模式内高。Otsuka等[55]在研究材料类别的VSTM中发现梭状回的激活与中央前回和中央后回的激活有关,说明梭状回可能参与的不同材质在视觉刺激和触觉刺激之间差异的VSTM通过触觉来调节。触觉刺激对双眼竞争的影响是由视觉和触觉中常见的原始特征决定的,如:方向和空间频率[65]。当视-触觉刺激空间频率相同时,视-触觉交互作用最大,但当触觉刺激物距离视觉刺激物大于30 cm时,触觉刺激将对双眼竞争无影响[66]。Ono等[65]考虑原始特征特征条件下,触摸粗糙织物和光滑大理石并进行双眼竞争测试时,发现与视觉图像一致的触觉刺激延长该视觉刺激的主导时间,粗糙织物比光滑大理石所占有的主导时间长,表明触觉刺激对双眼竞争下的交替视觉感知还可以通过粗糙度来调节。
在视-触觉短时记忆方面,织物纹理的STM可被保留并在视觉和触觉感知之间传递,无论是触觉或视觉感官编码,以及回忆织物的顺序如何,都可以实现织物纹理的跨模式识别[67],虽然短时记忆中的顺序保持与呈现的材料性质无关,但抑制左侧缘上回激活会扰乱视觉、触觉刺激任务中的顺序保持[68]。触觉刺激是评估记忆空间-触觉联系能力的有效刺激,但记忆空间-视觉联系的能力占主导地位[69]。Dehghan等[70]在研究向手指施加压力刺激对视觉WM的跨模式交互作用时发现,视觉WM表现不会受到触觉压力刺激的影响。
总结以上研究发现,感知材料属性的激活脑区的识别表征通常在多感觉区域交叉模式整合,腹侧视觉皮层大脑网络具备处理材料特性的多感官表征。视觉信息和触觉信息在脑区激活中是相互反应影响的,而且织物纹理的STM也可跨模式识别,视觉刺激占主导。
3 结论与展望
综上所述,fMRI技术在织物刺激脑感知的研究领域主要涉及轻微触觉刺激、接触压力刺激、视觉刺激、视-触觉跨模式刺激,目前尚处于探索响应织物刺激的激活脑区阶段,研究脑区主要聚焦于躯体感觉皮层、运动皮层、腹侧视觉皮层,表征指标主要为激活强度和激活占比。但基于fMRI织物刺激脑感知研究仍存在一些问题,主要有如下:
a)虽然fMRI在触觉刺激和视觉刺激均有了较大突破,但在织物热湿刺激领域尚待进一步探索和挖掘。
b) 触觉短时记忆主要研究集中在探索振动触觉刺激和触觉空间刺激的特性,并且织物属性的短时记忆研究较少,材料类别的视觉短时记忆大脑机制尚不清楚,应进一步探索织物感知、认知和记忆的潜在机制以及短时记忆在预测重复抑制中的作用。
c)织物刺激信息在特征激活脑区的流动过程和有效连接以及长时间刺激作用下,脑区的动态适应调节机制尚未明确认知。
d)脑区的干扰因素众多,如何排除各类生理干扰、心理干扰、外部环境干扰、实验因素对脑区舒适度表征的影响,或者发现各类干扰因素对特征感知脑区的影响规律并有效消除,对进一步实现织物舒适度的大脑原位表征具有重大意义。
参考文献:
[1]MALEK A S, ELNAHRAWY A, ANWAR H, et al. From fabric to smart T-shirt: Fine tuning an improved robust system to detect arrhythmia [J].Textile Research Journal, 2022, 92(17-18): 3204-3220.
[2]AWAIS M, KRZYWINSKI S, WENDT E. A novel modeling and simulation approach for the prediction of human thermophysiological comfort[J]. Textile Research Journal, 2021, 91(5-6): 691-705.
[3]K1RC1 F, KARAMANLARGIL E, DURU S C, et al. Comfort Properties of medical compression stockings from biodesigned and cotton fibers[J]. Fibers and Polymers, 2021, 22(10): 2929-2936.
[4]CAMILLIERI B, BUENO M-A, FABRE M, et al. From finger friction and induced vibrations to brain activation: Tactile comparison between real and virtual textile fabrics[J]. Tribology International, 2018, 126: 283-296.
[5]TANG W, ZHANG M, CHEN G, et al. Investigation of tactile perception evoked by ridged texture using ERP and non-linear methods[J]. 2021, 15: 756.
[6]苑洁.基于fMRI的织物接触压力舒适性脑感知表征[D].上海:东华大学,2019:78-79.
YUAN Jie. Brain Perception Representation of Fabric Contact Pressure Comfort Based on fMRI[D]. Shanghai: Donghua University, 2019: 107-116. 2019: 78-79.
[7]LIU Y J, CHEN D S. An analysis on EEG power spectrum under pressure of girdle[J]. International Journal of Clothing Science and Technology, 2015, 27(4): 495-505.
[8]尹玲.基于心率變异和脑波分析的塑身腹带着装压力舒适性研究[D].上海:东华大学,2012:107-116.
YIN Ling. Study on Pressure Comfort in Wearing a Waist-Nipper on the Basis of Human's Heart Rate Variability and Electronencephalogram[D].Shanghai: Donghua University, 2012: 107-116.
[9]夏羽.基于神经电生理学的丝织物触感评价和认知研究[D].苏州:苏州大学,2017:28-32.
XIA Yu. Research on Silk Tactile Perception Cognitive Evaluation Based on the Electric Neurophysiology[D]. Suzhou: Soochow University, 2017: 28-32.
[10]TANG W, LIU R, SHI Y, et al. From finger friction to brain activation: Tactile perception of the roughness of gratings [J]. Journal of Advanced Research, 2020, 21: 129-139.
[11]唐玮,张梅梅,刘瑞,等.不同尖锐度纹理形状的摩擦触觉感知与表征研究[J].摩擦学学报2021,41(3):373-381.
TANG Wei, ZHANG Meimei, LIU Rui, et al. Tactile perception of texture shape with different sharpness from finger friction to brain activation[J]. Tribology, 2021, 41(3): 373-381.
[12]TANG W, LU X, CHEN S, et al. Tactile perception of skin: research on late positive component of event-related potentials evoked by friction[J]. The Journal of the Textile Institute, 2020, 111(5): 623-629.
[13]刘陶峰,李一员,李炜,等.确定性纹理表面特征高度对皮肤摩擦感知的影响[J].西南交通大学学报,2020,55(2):372-378.
LIU Taofeng, LI Yiyuan, LI Wei, et al. Influence of surface feature height of deterministic texture on tactile perception of fingertip[J]. Journal of Southwest Jiaotong University, 2020, 55 (2): 372-378.
[14]LIU Y J, CHEN D S. The influence of clothing pressure exerted by girdle on inhibition ability of young females[J]. International Journal of Clothing Science and Technology, 2016, 28(5): 712-722.
[15]陳雁李栋高.服装颜色的感觉生理研究[J].纺织学报,2004,25(3):68-69.
CHEN Yan, LI Donggao. Study on the color perception of the garment[J]. Journal of Textile Research, 2004, 25(3): 68-69.
[16]STYLIOS G K, CHEN M. The concept of psychotextiles; interactions between changing patterns and the human visual brain, by a novel composite smart fabric[J]. Materials, 2020, 13(3): 725.
[17]莫换平.纺织品冷暖色搭配视觉认知研究[D].苏州,苏州大学,2020:28-46.
MO Huanping. Research on the Cognitive Cognition of Textile Cold and Warm Colors[D]. Suzhou: Soochow University, 2020: 28-46.
[18]DING M, SONG M, PEI H, et al. The emotional design of product color: An eye movement and event-related potentials study[J]. Color Research & Application, 2021, 46(4): 871-889.
[19]OGAWA S, LEE T M, KAY A R, et al. Brain magnetic resonance imaging with contrast dependent on blood oxygenation[J]. Proceedings of the National Academy of Sciences, 1990, 87(24): 9868-9872.
[20]王其才.基于FMRI的织物触觉刺激大脑感知表征[D].上海:东华大学,2015.
WANG Qicai. Brain Cognitive Characterization of Tactile Stimulation from Fabric Touch Based on FMRI[D]. Shanghai:Donghua University, 2015.
[21]PAULING L, CORYELL C D. The magnetic properties and structure of hemoglobin, oxyhemoglobin and carbon-monoxyhemoglobin[J]. Proceedings of the National Academy of Sciences, 1936, 22(4): 210-216.
[22]BUXTON R B. Introduction to Functional Magnetic Resonance Imaging: Principles and Techniques[M].
Cambridge: Cambridge university press, 2009.
[23]FRISTON K J, ASHBURNER J, KIEBEL S J, et al.Statistical Parametric Mapping: The Analysis of Functional Brain Images[M]. Amsterdam: Elsevier, 2011.
[24]ASHBURNER J. A fast diffeomorphic image registration algorithm[J]. NeuroImage, 2007, 38(1): 95-113.
[25]Poldrack R A, Mumford J A, Nichols T E. Handbook of functional MRI data analysis[M]. Cambridge University Press, 2011, 15-17.
[26]YUAN J, YU W, WANG Q, et al. A potential brain zone perceiving a comfortable fabric pressure touch[J]. Textile Research Journal, 2018, 89(17): 3499-3505.
[27]WANG Q, TAO Y, ZHANG Z, et al. Representations of fabric hand attributes in the cerebral cortices based on the Automated Anatomical Labeling atlas[J]. 2019, 89(18): 3768-3778.
[28]WANG Q, TAO Y, YUAN J, et al. Application of Brodmann's area maps for cortical localization of tactile perception evoked by fabric touch[J]. Fibers and Polymers, 2019, 20(4): 876-885.
[29]苑洁,娄琳,王其才.织物触觉舒适度大脑感知技术研究进展[J].纺织学报,2022,43(09):211-217.
YUAN Jie, LOU Lin, WANG Qicai. Research progress of brain perception technology for tactile comfort of fabric [J]. Journal of Textile Research, 2022, 43 (09): 211-217.
[30]YEON J, KIM J, RYU J, et al. Human brain activity related to the tactile perception of stickiness[J]. Frontiers in Human Neuroscience, 2017, 11: 8.
[31]苑潔,于伟东,陈克敏,等.基于功能磁共振的织物触压舒适度脑感知研究进展[J].纺织学报,2017,38(10):146-152.
YUAN Jie, YU Weidong, CHEN Kemin, et al. Research progress of brain perception of fabric touch comfort based on functional magnetic resonance[J]. Journal of Textile Research, 2017, 38(10): 146-152.
[32]GUILLERY R W. Brodmann's 'Localisation in the Cerebral Cortex'[J]. The Journal of Anatomy, 2000, 196(3): 493-496.
[33]YUAN J, XU C, WANG Q, et al. Brain signal changes of sensory cortex according to surface roughness of boneless corsets[J]. Textile Research Journal, 2020, 90(1): 76-90.
[34]GURTUBAY-ANTOLIN A, LEON-CABRERA P, RODRI-GUEZFORNELLS A. Neural evidence of hierarchical cognitive control during haptic processing: An fMRI study[J]. Eneuro, 2018, 5(6): 295-318.
[35]RAJAEI N, AOKI N, TAKAHASHI H K, et al. Brain networks underlying conscious tactile perception of textures as revealed using the velvet hand illusion [J]. Human Brain Mapping, 2018, 39(12): 4787-4801.
[36]SO Y, KIM S P, KIM J. Perception of surface stickiness in different sensory modalities: An functional MRI study[J]. 2020, 31(5): 411-415.
[37]WANG Q, YU W, HE N, et al. Investigation of the cortical activation by touching fabric actively using fingers[J]. Skin Research and Technology, 2015, 21(4): 444-448.
[38]WANG Q, YU W, CHEN K, et al. Brain cognitive comparison of fabric touch on human glabrous and hairy skin[J]. Textile Research Journal, 2016, 86(3): 318-324.
[39]WANG Q, YU W, CHEN K, et al. Brain discriminative cognition on the perception of touching different fabric using fingers actively[J]. Skin Research and Technology, 2016, 22(1): 63-68.
[40]KITADA R, DOIZAKI R, KWON J, et al. Brain networks underlying tactile softness perception: A functional magnetic resonance imaging study[J]. Neuroimage, 2019, 197: 156-166.
[41]WANG Q, TAO Y, SUN T, et al. Analysis of brain functional response to cutaneous prickling stimulation by single fiber[J]. Skin Research and Technology, 2021, 27(4): 494-500.
[42]YUAN J, YU W, CHEN K, et al. A potential new fabric evaluation approach by capturing brain perception under fabric contact pressure[J]. Textile Research Journal, 2018, 89(16): 3312-3325.
[43]童新宇,吳新丽,李思儒,等.人手指振动触觉感知的短时记忆特性[J].生理学报,2020,72(5):643-650.
TONG Xinyu, WU Xinli, Li Siru, et al. Short-term memory characteristics of vibro-tactile perception in human fingertips[J]. Acta Physiologica Sinica, 2020 and 72 (5): 643-650.
[44]ZHANG J, TAO H, JIANG X. Cognitive Behavior Difference Based on Sensory Analysis in Tactile Evaluation of Fabrics[C]//International Conference on Applied Human Factors and Ergonomics. Springer, Cham, 2019: 430-437.
[45]CHUNG Y G, HAN S W, KIM H S, et al. Adaptation of cortical activity to sustained pressure stimulation on the fingertip[J]. BMC Neuroscience, 2015, 16(1): 1-11.
[46]OISHI Y, IMAMURA T, SHIMOMURA T, et al. Visual texture agnosia in dementia with Lewy bodies and Alzheimer's disease[J]. Cortex, 2018, 103: 277-290.
[47]CANT J S, XU Y. The contribution of object shape and surface properties to object ensemble representation in anterior-medial ventral visual cortex[J]. Journal of Cognitive Neuroscience, 2017, 29(2): 398-412.
[48]LACEY S, SATHIAN K. Visuo-haptic multisensory object recognition, categorization, and representation[J]. Frontiers in Psychology, 2014, 5: 730.
[49]SUZUKI K. Visual texture agnosia in humans[J]. Shinkei Kenkyu no Shinpo, 2015, 67(6): 701-709.
[50]JACOBS R H A H, BAUMGARTNER E, GEGENFURTNER K R. The representation of material categories in the brain[J]. Frontiers in Psychology, 2014, 5: 146.
[51]ALVAREZ G A, CAVANAGH P J P S. The capacity of visual short-term memory is set both by visual information load and by number of objects [J]. Psychological Science, 2004, 15(2): 106-111.
[52]HARRISON A, JOLICOEUR P, MAROIS R J C C. "What" and "where" in the intraparietal sulcus: An fMRI study of object identity and location in visual short-term memory[J]. Cerebral Cortex, 2010, 20(10): 2478-2485.
[53]SHEREMATA S L, BETTENCOURT K C, SOMERS D C J J O N. Hemispheric asymmetry in visuotopic posterior parietal cortex emerges with visual short-term memory load [J]. Journal of Neuroscience, 2010, 30(38): 12581-12588.
[54]XU Y, CHUN M M J N. Dissociable neural mechanisms supporting visual short-term memory for objects[J]. Nature, 2006, 440(7080): 91-95.
[55]OTSUKA S, SAIKI J J H. Neural correlates of visual short-term memory for objects with material categories[J]. Heliyon, 2019, 5(12): e03032.
[56]LI C, KOVCS G, TRAPP S J B P. Visual short-term memory load modulates repetition related fMRI signal adaptation[J]. Biological Psychology, 2021, 166: 108199.
[57]KOMATSU H, GODA N. Neural mechanisms of material perception: Quest on Shitsukan[J]. Neuroscience, 2018, 392: 329-347.
[58]ECK J, KAAS A L, MULDERS J L, et al. The effect of task instruction on haptic texture processing: The neural underpinning of roughness and spatial density perception[J]. Cerebral Cortex, 2016, 26(1): 384-401.
[59]GALLIVAN J P, CANT J S, GOODALE M A, et al. Representation of object weight in human ventral visual cortex[J]. Current Biology, 2014, 24(16): 1866-1873.
[60]NEWMAN SD, KLATZKY RL, LEDERMAN SJ, et al. Imagining material versus geometric properties of objects: An fMRI study[J]. CognitiBrain Res, 2005, 23: 235-246.
[61]KIM Y, USUI N, MIYAZAKI A, et al. Cortical regions encoding hardness perception modulated by visual information identified by functional magnetic resonance imaging with multivoxel pattern analysis[J]. Front Syst Neurosci, 2019, 13: 52.
[62]XIAO B, BI W, JIA X, et al. Can you see what you feel? Color and folding properties affect visual-tactile material discrimination of fabrics[J]. Journal of Vision, 2016, 16(3): 1-15,34.
[63]WEISSER V, STILLA R, PELTIER S, et al. Short-term visual deprivation alters neural processing of tactile form[J]. Experimental Brain Research, 2005, 166(3): 572-582.
[64]O'CALLAGHAN G, O'DOWD A, SIMES-FRANKLIN C, et al. Tactile-to-visual cross-modal transfer of texture categorisation following training: An fMRI study [J]. Frontiers in Integrative Neuroscience, 2018, 12: 24.
[65]ONO M, HIROSE N, MORI S J, et al. Tactile information affects alternating visual percepts during binocular rivalry using naturalistic objects[J]. Cognitive Research: Principles and Implications, 2022, 7(1): 1-12.
[66]LUNGHI C, MORRONE M C. Early interaction between vision and touch during binocular rivalry[J]. Multisensory Research, 2013, 26(3): 291-306.
[67]CHIARI I, HIROKI Y, SACHIKO S J J O T E. Visio-haptic cross-modal recognition for fabrics [J]. Journal of Textile Engineering, 2020, 66(1): 1-6.
[68]GUIDALI G, PISONI A, BOLOGNINI N, et al. Keeping order in the brain: The supramarginal gyrus and serial order in short-term memory[J]. Cortex, 2019, 119: 89-99.
[69]MUNOZ-MONTOYA F, JUAN M-C, MENDEZ-LOPEZ M, et al. Augmented reality based on SLAM to assess spatial short-term memory[J]. IEEE Access, 2018, 7: 2453-2466.
[70]DEHGHAN NAYYERI M, BURGMER M, PFLEIDERER B J P O. Impact of pressure as a tactile stimulus on working memory in healthy participants[J]. Plos One, 2019, 14(3): e0213070.
Abstract: At present, the brain perception mechanism in the field of fabric comfort is not clear, and the existing characterization technology of fabric comfort has not been quantified. Functional magnetic resonance imaging technology has shown good technical advantages in the field of fabric stimulation brain perception representation with its high spatial resolution, which is of great significance for exploring the brain perception mechanism of fabric comfort.
By exploring the changes between stimulation signals and brain activation signals, fMRI technology can effectively and accurately identify the relevant brain regions under fabric stimulation, and complete the brain in situ perception representation of fabric comfort. This paper reviews the research status of brain perception of fabric stimulation based on fMRI technology from the three fields of tactile stimulation, visual stimulation, and visual-tactile cross-stimulation. Among them, the somatosensory cortex, motor cortex and ventral visual cortex are the relevant response brain regions for tactile stimulation, visual stimulation and visual-tactile cross-stimulation. The characteristic indicators are mainly activation intensity and activation proportion.
At present, the study of brain perception of fabric stimulation based on fMRI technology has become a popular research topic. It has been confirmed that the activation point coordinates, activation intensity, activation proportion, functional connectivity and other information of relevant brain regions under fabric stimulation can effectively represent the brain perception of fabric stimulation, which provides great potential for in situ representation of fabric stimulation.
Based on the principle of brain perception representation of fMRI technology, this paper summarizes the research status of brain perception of fMRI technology in fabric slight touch stimulation, contact pressure stimulation, visual stimulation, and visual-tactile cross-stimulation. It is concluded that SI, SII and motor cortex in the somatosensory cortex are related to slight touch stimulation and contact pressure stimulation of fabrics. Among them, SI is related to smoothness and softness, and SI, SII and motor cortex are related to roughness, adhesion and itching. On the right side, SII is the fabric comfortable pressure perception brain area, SI and amygdala are the fabric uncomfortable pressure perception brain area, while amygdala is the fabric compression pressure perception brain area. The changes of functional connectivity between SI and SII are related to the duration of stress stimulation. The ventral visual cortex brain network is not only a relevant brain area for the perception of fabric visual stimuli, but also a multi-sensory representation brain area for material properties. The fusiform gyrus is not only sensitive to material category perception, but also related to visual short-term memory. In the future fabric stimulation process based on fMRI technology, it is important to master and control or eliminate interference factors for fabric comfort representation research. However, in the field of thermal and wet stimulation of fabrics, the brain mechanism of short-term memory of fabric properties, the flow process and effective connection of characteristic activated brain regions, and the dynamic adaptation and regulation mechanism of brain regions under long-term stimulation need to be further explored.
Keywords: comfort; brain perception; functional magnetic resonance imaging; tactile perception; vision perception
猜你喜欢
纺织科学研究(2021年9期)2021-10-14
海外星云(2021年6期)2021-10-14
铁道通信信号(2020年1期)2020-09-21
特别健康(2018年3期)2018-07-04
浙江工业大学学报(2017年5期)2018-01-22
演艺科技(2016年10期)2016-11-26
戏剧之家(2016年19期)2016-10-31
启迪与智慧·教育版(2016年8期)2016-10-20
工业设计(2016年4期)2016-05-04
工业设计(2016年10期)2016-04-16
