
浙江省稻瘟病研究进展
2023-05-29 05:52杨海龙王蕾李白蔡金洋
中国稻米 2023年3期
杨海龙 王蕾 李白 蔡金洋
(浙江省嘉兴市农业科学研究院,浙江 嘉兴 314016;第一作者:774795537@qq.com;*通讯作者:cjyjiaxing@163.com)
由子囊真菌侵染水稻引起的稻瘟病是水稻主要病害之一[1]。全球水稻产量每年因稻瘟病影响会减产10%~30%[2]。我国稻瘟病发病面积约500 万hm2,受其影响水稻产量减少10%~20%,严重地区可达到50%,甚至绝收[3-4]。浙江省作为长江中下游地区主要粮食生产地,稻瘟病常年发生,给当地粮食生产带来了巨大经济损失,如2014 年浙江省杭嘉湖、宁绍平原突发流行稻瘟病,发生面积达2.33 万hm2,损失超过350 kg/hm2,秀水321、嘉粳88 和秀水134 等主推品种均不同程度发病[5-6]。推广抗病品种是目前防治稻瘟病最有效、绿色和经济的方法,抗稻瘟病基因的挖掘、分析和利用是选育抗病品种的重要措施[7-8]。由于稻瘟病病菌小种的不断变异,致病性不断进化,现在有必要通过基因定位来评估抗性基因对稻瘟病抵抗效果的强度。文章综述了浙江省稻瘟病优势种群、主栽品种抗性情况和抗性基因利用情况,以期为浙江省稻瘟病抗性研究和有效防治提供建议和参考。
1 浙江省稻瘟病的优势种群
长江中下游地区种植的水稻品种丰富多样,籼稻、粳稻和籼粳杂交稻均有,稻瘟病菌的致病性也容易发生变异,不同生理小种的组成和结构也非常复杂[9]。我国基于本土的稻瘟病生理小种鉴别寄主(Tetep、珍龙13、四丰43、东农363、关东51、合江18 和丽江新团黑谷)将稻瘟病生理小种划分为ZA(籼型)、ZB(籼型)、ZC(籼型)、ZD(粳型)、ZE(粳型)、ZF(粳型)和ZG(粳型)[9]。浙江省稻瘟病菌优势小种随着时间和地域变化而变化[6,10-11]。柴荣耀等[10]于20 世纪90 年代对浙江省太湖流域488 个稻瘟病菌种群进行鉴定,结果发现,ZD、ZA和ZE 属于优势种群,优势生理小种为ZD3、ZE3、ZA49和ZD1。肖丹凤[11]在2013 年对浙江省89 个稻瘟病菌株进行鉴定,发现ZB 为浙江省的优势种群,出现频率63.04%,其中优势生理小种为ZB15、ZB9、ZA1和ZB13。王晔青等[6]在2014—2015 年对嘉兴市南湖区、秀洲区和桐乡市稻瘟病发生情况进行监测,发现生理小种ZD1、ZE1和ZD3对粳稻的致病性强。HAO 等[12]于2021 年收集了浙江省近4 年46 个稻瘟病菌分离物(籼型分离物23 个,粳型分离物23 个),对1 252 个水稻品种进行接种,结果发现23 个籼型分离物大部分属于籼型小种,23 个粳型分离物中半数为籼稻种、半数为粳稻种。大多数籼稻品种感染籼型分离物后,其发病程度比感染粳型分离物的严重,分离菌株的种族与采集的水稻品种种类关系不大,而与样品采集年份有关。
2 浙江省水稻品种的稻瘟病抗性状况
水稻是浙江第一大粮食作物,据统计,全省常年播种面积在80.0 万hm2左右[13]。2021 年浙江省农业农村厅发布的水稻主导品种有22 个,其中早籼稻有甬籼15、中早39 和中嘉早17 等,粳稻有浙粳99、秀水14 和秀水121 等,籼粳杂交稻有甬优1540、甬优15 和甬优538 等。在稻瘟病高发地区,上述有些材料会不同程度发病[6,14]。鄂志国等[15]分析了浙江省近20 年来水稻品种的抗性变化,发现浙江最近数年审定品种的稻瘟病综合指数从2012 年的平均“中抗”水平下降至2018 年“中抗-中感”水平,但2003—2012 年稻瘟病抗性平均水平一直高于“中抗”。研究还发现尽管近年来浙江省稻瘟病抗性水平有所下降,但还是高于同稻区全国其他品种的平均水平。施俊生等[13]分析了2007—2018 年浙江省审定品种的抗性表现,结果发现审定的262 个品种中有201 个的稻瘟病抗性水平达到中抗及以上。笔者基于中国水稻品种及其系谱数据库(http://www.ricedata.cn/),筛选出2012—2021 年浙江省审定的203个品种,分析其稻瘟病抗性综合指数,结果表明,2012—2021 年浙江省稻瘟病抗性水平大体呈下降趋势,从2019 年开始又有所提高(图1)。
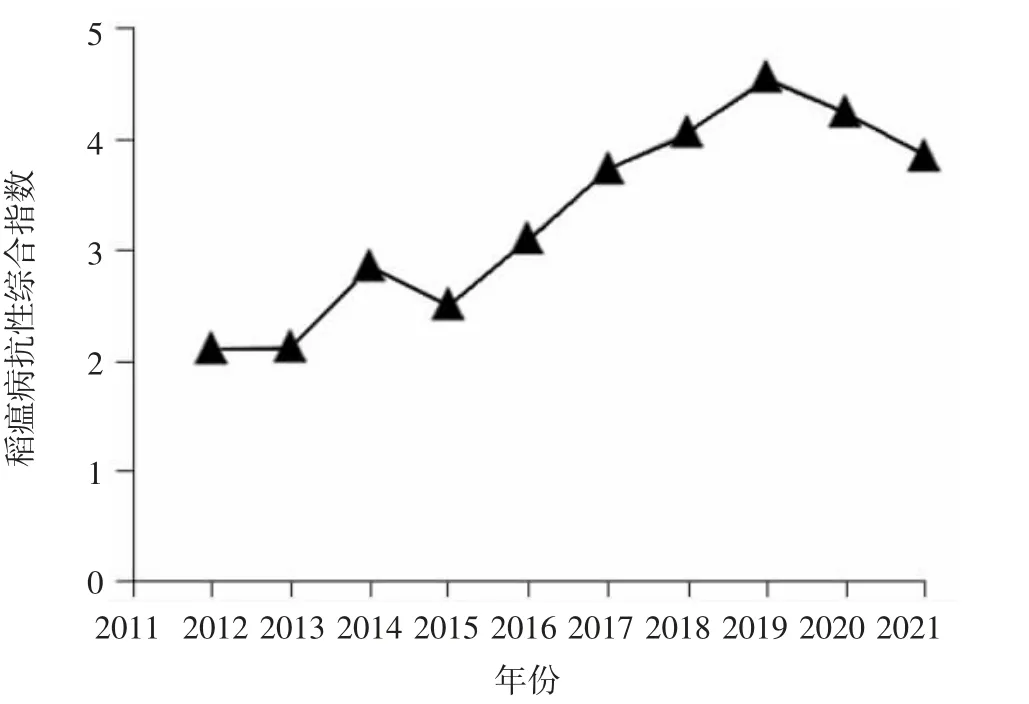
图1 2012—2021 年浙江省审定品种的稻瘟病抗性情况
3 浙江省稻瘟病抗性基因利用情况
前人于2003 年提出基因功能标记的概念,即1 个分子标记位点代替1 个特定的等位基因,可以基于分子标记的筛选来对特定性状进行筛选[16]。目前已经通过分子标记技术发现至少69 个抗病位点100 多个稻瘟病抗性主效基因,已经有36 个抗病基因被克隆,分别 是Pi1[17]、Pi2[18]、Pi5[19]、Pi9[20]、Pi25[21]、Pi33[22]、Pi35[18]、Pi36[23]、Pi37[24]、Pi50[20]、Pi56[25]、Pi63[26]、Piz-t[20]、Pit[27]、Pit-a[18]、Pia[28]、Pib[29]、Pish[30]、Pib1[18]、Pik[31]、Pik-m[18]、Pik-p[18]、Pid3[21]、Pi-d2[32]、pi21[33]、Ptr[18]、Pi-CO39[19]、Pigm[20]、Pii[20]、Pi54[33]、Pi42[18]、Pid3-A4[18]、Pi54rh[31]、Pi54of[34]、Pike[35]和Pikh[36]。绝大多数抗性基因定位于第6 和第11 染色体上,少部分定位于第1、第9 和第12 染色体上,在第2、第4 和第8 染色体上也发现各有1 个抗性基因[18]。不同类型的基因在抵抗稻瘟病的效果上有着不同的作用。例如6 号染色体克隆的稻瘟病抗性基因Pi2、Pi9、Pi25、Pi50、Piz-t、Pid3、Pid2、Pigm 和Pid3-A4 中,Pi2 对我国13 个稻区的792 个菌株表现为抗[18];Pi25 对稻瘟菌菌系92-183(小种ZC15)有较强的叶瘟和穗瘟抗性。11 号染色体克隆的稻瘟病抗性基因Pi1、Pi35、Pia、Pik、Pi-CO3 和Pi54 中,来自西非的Pi1 对我国8 个稻区的稻瘟病菌株表现为抗[18];Pi35 和Pia 有较好的抗叶瘟能力;Pik 对我国许多稻瘟病小种有稳定的抗性[19]。研究还发现,定位于第4 染色体上的pi21 是第1 个被定位的隐性抗瘟性基因位点,研究发现,携带隐性pi21 等位基因的近等基因系AA-pi21 对10 种稻瘟病病菌具有抗性,但抗性没有专一抗病性的R 基因强[33]。
浙江省的育种工作者将分子标记辅助技术结合到传统育种中,培育出一批对稻瘟病抗性强和抗谱广的水稻品种。何海燕等[36-37]在2019 年分析了浙江省40 个水稻栽培品种10 个稻瘟病抗性基因的分布情况,结果发现,Pib、Pia 和Pit-a 抗性基因的分布频率最高,分别为63.04%、58.70%和47.83%,但浙江省主要栽培品种中没有发现Pi1 的分布。沈浙南等[38]利用Pi1、Pi5、Pi2、Pi9、Pia、Piz-t、Pigm、Pib、Pik 和Pikh 等10 个抗稻瘟病基因的分子标记对63 个浙江省主栽品种(24 个常规籼稻、30 个常规粳稻和9 个籼粳杂交稻)的抗病基因进行鉴定,发现Pikh、Pia、Piz-t、Pib、Pi1 和Pik 等抗病基因利用频率较高,而Pigm 和Pi9 的利用频率较低。翟荣荣等[34]对浙江省12 个常规晚粳稻品种10 个稻瘟病抗性基因进行了分子检测,结果发现,12 个供试水稻品种中都携带Pib、Pi25、Pi40 和Pi41 抗性基因,Pi2和Pikm 的分布频率为58.33%,Pit-a 的分布频率为50.00%,Pi5 的分布频率为33.33%,Pi9 的分布频率为25.00%,没有发现Pigm 的分布。稻瘟病菌的快速进化和优势生理小种的频繁更替,仅含单个抗病基因的抗病品种在连续种植几年后很容易丧失抗性,因此,抗病育种中需聚合多个抗稻瘟病基因。但在聚合多个抗病基因时也需要考虑所聚合基因的主效性、正向性和负向性[39]。例如Pigm 与Pib、Pi2 与Pi1、Pib、Pi1 与Pik、Pib以及Pikh 与Pib 之间是正向显著相关,而Pizt 与Pi2、Pi1 是负向极显著相关[37]。
4 浙江省稻瘟病无毒基因研究进展
水稻与稻瘟病菌之间的特异互作关系符合基因对基因学说[40]。无毒基因(AVR)是一类能够诱发水稻产生抗病性的病原物遗传因子,在水稻稻瘟病菌与水稻互作中,无毒基因的作用是转录翻译成无毒蛋白质,然后被水稻细胞内的抗性基因识别,从而对稻瘟病产生抗性[41]。目前已经鉴定到与稻瘟病抗性基因对应的无毒基因有12 个被克隆(AVR-Pita、ACE1、AVR-Pizt、AVR-Pii、AVR-Pi9、AVR-Pib、AVR-Pia、AVR-Pi54、AVR-Pik/km/kp 和AVR1-CO39 等)[41-42]。浙江省关于水稻稻瘟病菌无毒基因的研究较少。穆慧敏[43]利用无毒基因研究了浙江省稻瘟病菌群体的遗传多样性,结果发现,浙江省丽水市稻瘟病菌的遗传多样性最高、舟山市最低,浙江各地的无毒基因的变异类型主要为缺失和插入,大多数菌种含有基因ACE1 和AVR-Pizt,少部分含有AVR-Pia、AVR-Pii 和PWL4,未含有AVR1-CO39 和PWL1。生产上水稻品种的抗病性往往因为稻瘟病小种的变异而过早的丧失抗性,对于水稻育种家来说,了解无毒基因的变异机制,能够更加深刻认识到无毒基因的位点变异、理解病原物丧失无毒功能的机制和小种产生的规律,从病原物着手,揭示其遗传变异的本质,为稻瘟病防治提供理论基础。

表1 已克隆水稻抗稻瘟病基因相关信息
5 展望
浙江省是我国长江中下游主要的粮食生产基地,长期以来稻瘟病严重影响着水稻生产[5-6]。稻瘟病菌耐药性的增强、病原菌生理小种的进化、致病性的变异、水稻品种抗性的失效和大规模单一化种植是导致近年水稻稻瘟病日益加重的主要原因[10,44]。因此,挖掘和利用新的稻瘟病抗性基因对防治稻瘟病具有重要意义。2021 年“中央一号文件”指出要推进绿色农业发展、持续推进化肥农药减量增效、推广农作物病虫害绿色防控产品和技术,因此在水稻稻瘟病的治理上更应以环保为主。为进一步控制好稻瘟病对浙江水稻生产的影响,更加绿色、健康和高效防治稻瘟病,需开展以下三方面研究:第一,更新水稻栽培管理模式。根据浙江省不同地域土壤的养分情况和基肥施用情况,决定氮肥的施用量,切忌氮肥用量过大,造成水稻贪青晚熟,同时增加钾肥的使用,以提高水稻的抗病性。水分管理上采用浅湿灌溉,该浅则浅、该晒则晒、该深则深,可适当晾田壮根,提高水稻的抗病能力。第二,利用分子标记聚合多个抗稻瘟病基因。但抗病基因聚合不是简单的基因叠加效应,抗性基因的不同组合可能直接导致不同的抗性水平,育种家在育种过程中,可以考虑靶基因的优良等位基因[45]。第三,了解稻瘟病基因与其对应的无毒基因在田间水稻稻瘟病菌的毒力组成是认识ETI免疫机制的关键,目前育种家已对AVR-Pita、ACE1、AVR-Pizt、AVR-Pii、AVR-Pi9 等12 个无毒基因进行分析、检测、标记及克隆,分析浙江省地区无毒基因的组成及分布特点,可为下一步预测稻瘟病菌的群体变化趋势,为能准确、有效和快速的预测田间潜在流行致病菌系奠定基础[43]。
猜你喜欢
今日农业(2022年4期)2022-06-01
作物学报(2022年6期)2022-04-08
南方农业学报(2022年11期)2022-03-18
植物保护(2017年4期)2017-08-09
浙江农业学报(2017年1期)2017-05-17
中国马铃薯(2017年1期)2017-03-02
河南农业(2016年6期)2016-11-26
作物学报(2016年10期)2016-10-19
新农业(2016年20期)2016-08-16
现代农业(2016年5期)2016-02-28
