
卵黄原蛋白在昆虫生理猖獗机制中的作用及应用前景
2023-05-27 14:12鲁玉杰苗世远王争艳杨斌斌
环境昆虫学报 2023年2期
鲁玉杰,鲁 婷,苗世远,王争艳,杨斌斌
(1.河南工业大学粮食和物资储备学院,郑州 450001;2. 江苏科技大学粮食学院,江苏镇江 212100)
昆虫个体较小、易躲藏、环境适应性强、食性广泛且具有耐饥能力强、抗逆性强、繁殖速度快等惊人的优势,使其成为世界上种类繁多、数量庞大、分布广泛的动物种类。在农业生产中,许多昆虫种群因气候变化、人工杀虫剂滥用等因素的影响,逐渐从原先的次要害虫变为重要害虫,容易在适生的环境条件下发生猖獗并且可能爆发成灾。近年,草地贪夜蛾Spodopterafrugiperda入侵我国云南随后迅速扩散至全国,因适应区域广、迁移性强、繁殖力高、抗药性强和暴食为害的特点对我国农业生产造成了巨大的损失(吴秋琳,2019)。有研究表明,在田间种群发生较重情况下,草地贪夜蛾在美洲和非洲造成玉米减产20%~72%,严重威胁玉米种植户经济收入和全球粮食安全(万敏等,2022)。作为粮库中常见的害虫,储粮书虱,如嗜虫书虱Liposcelisentomophila已经从20年前的次要害虫成为危及世界粮食和食品安全的重要储粮害虫之一。昆虫猖獗不仅会造成农业减产、威胁储粮安全、增大经济损失,同时还会导致各种皮肤病、呼吸系统疾病等爆发(Ahmedanietal.,2010)、病毒的传播,如褐飞虱Nilaparvatalugens猖獗传播的水稻黑条矮缩病毒病严重危害水稻生产等问题(Sunetal.,2017)。因此,研究昆虫猖獗机制,控制昆虫猖獗问题的发生成为植物保护工作的重点和难点。
一般认为引起昆虫猖獗现象的机制主要是由生态及生理调控因素协同引发的一个动态响应过程,即昆虫逐渐适应生态,引发基因阶段式分化和突变的现象。气候变暖、CO2浓度升高及臭氧层破坏等生态因素加剧了害虫的发生世代。多数昆虫在繁殖的适温范围内繁殖力随温度的升高而增强,如甘蓝蚜Brevicorynebrassicae在适生温度范围内,温度越高繁殖能力越强,产卵量也更高(Sohetal.,2018)。CO2的升高对褐飞虱的增殖具有显著的促进作用,褐飞虱种群数量增加(Guru Pirasanna Pandietal.,2018)。紫外线照射棉铃虫Helicoverpaarmigera(Hübner)1~4 h后,其卵黄原蛋白(Vitellogenin,Vg)的表达量和产卵量会显著增加(王智健等,2014)。昆虫的强适应性在长期进化中引发基因突变现象,如植食性昆虫为了不断适应植物的防御反应,逐渐在形态上发生变异,包括多型翅,分化出口器,如咀嚼式、刺吸式以及虹吸式口器等(Hermsmeieretal.,2001)。多数昆虫食性的转变及扩大更是加剧了其猖獗,如大多数白蚁的食物原为枯木或草,但Donovan等(2001)发现其食物已经扩大到包括栽培真菌、碎屑和土壤有机质。更有昆虫已经进化出了利用专性共生细菌念珠菌CandidatusNasuiadeltocephalinicola携带额外Vgs的策略,以保证昆虫的最佳繁殖(Maoetal.,2020)。
昆虫适应能力的最典型特征之一是繁殖能力的增强,雌性昆虫不断进化,优化卵巢管结构和卵子发生过程等都使昆虫能够适应各种逆境而繁衍生息。在昆虫种群繁衍和扩增过程中,生殖系统对于维持其种群数量及后代延续至关重要,尤其是昆虫Vg基因已成为了昆虫繁殖的关键因子之一。Vg是由6~7 kb Vg mRNA合成的200 kDa左右的含有共价结合的碳水化合物、磷酸盐和非共价结合脂质的高分子量磷酸糖蛋白(Tufail and Takeda,2008);广泛存在于卵生雌性动物的血淋巴、脂肪体和卵巢中,是几乎所有卵生动物卵黄蛋白的前体(Yolk protein precursors,YPPs)(Sappingtonetal.,1998)。在大多数昆虫中,Vg在雌性昆虫脂肪体中经糖基化、脂化、磷酸化、蛋白水解裂解等过程合成后,释放到血淋巴,经结构修饰后成为由几个亚基组成的天然卵黄原蛋白,后被发育中的卵母细胞外表面的卵黄蛋白原受体(vitellogenin receptor,VgR)介导的内吞作用选择性吸收(Tufail and Takeda,2009;戈林泉和吴进才,2010)。Vg在昆虫卵黄发生(Vitellogenesis)过程及昆虫生殖过程起重要作用(Wuetal.,2018)。
目前,对于昆虫Vg的理化性质及分子调节通路等研究已有很多,但缺乏昆虫Vg与昆虫猖獗关系之间的系统总结与Vg的实践应用分析。因而,本文从Vg入手,阐述Vg对昆虫猖獗的影响与机制,探讨昆虫Vg作为害虫防治靶点的实际性操作技术的利弊,为研发害虫防治技术提供新靶标和新思路。
1 卵黄原蛋白在昆虫生理猖獗中的作用
害虫防治过度依赖或滥用化学农药易诱导害虫再猖獗现象的产生,再猖獗机制一般可分为生态再猖獗和生理再猖獗,其中,生理再猖獗主要是药剂引起害虫的毒物兴奋效应及补偿作用(刺激产卵),是药剂亚致死剂量刺激害虫生殖的现象。亚致死剂量是指由于昆虫个体对杀虫剂的不同接触程度以及所施用杀虫剂的降解程度,在害虫暴露于亚致死浓度或剂量的毒物时,对该虫生理或行为特征的影响,包括使害虫神经系统、发育速度、产卵能力、寿命、免疫力、性比以及摄食行为等发生改变(Lüetal.,2021)。所以药剂刺激昆虫Vg表达量上调成为害虫再猖獗发生的一条重要途径。
1.1 卵黄原蛋白对昆虫生殖的影响
迄今为止,已报道了200多种昆虫Vg基因,部分见表1。昆虫Vgs属于大脂质转移蛋白(LLTP)超家族,由3个高度保守的结构域组成,分别是脂质结合结构域(LPD_N)、未知功能结构域(DUF1943)、以及von Willebrand因子型D域(VWD)(Tufail and Takeda,2008)。大多数昆虫Vgs中发现了一些特定的保守单元,如切割位点RXXR、功能位点GL/ICG和DGXR以及C端附近的保守半胱氨酸残基,其中多丝氨酸束被认为是昆虫Vgs最显著的特征(Miaoetal.,2021)。
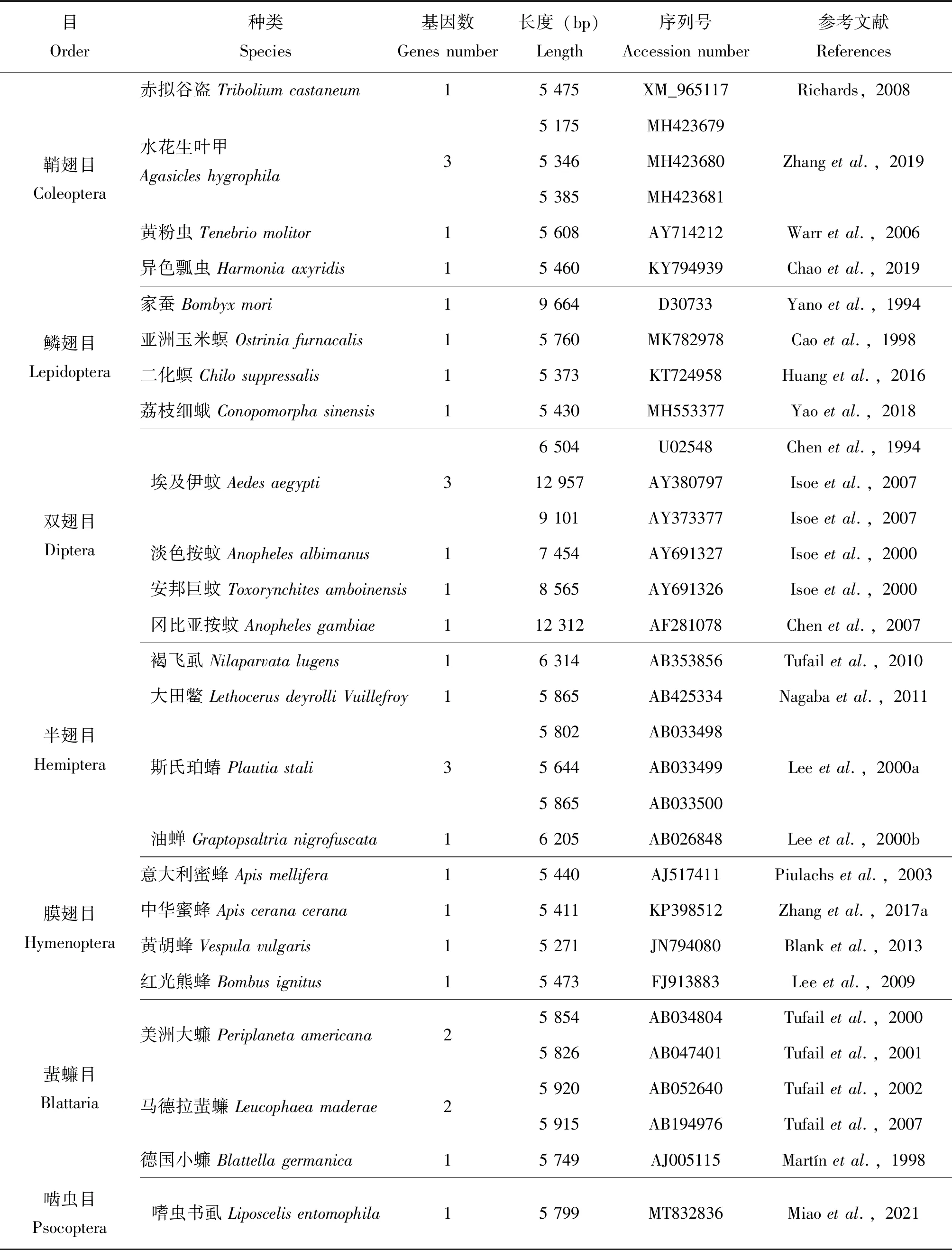
表1 部分昆虫Vg全长序列信息
脂肪体中Vg的合成是雌性昆虫卵黄发生的基础,决定着卵子成熟与胚胎发育的成功。对于大多数昆虫的卵黄发生,涉及脂肪体产生和分泌Vgs或其他YPPs,并通过膜结合受体VgR介导的胞吞作用分泌到血淋巴中,再转运到正在生长的卵母细胞中(Tufailetal.,2014)。进入卵母细胞后,Vgs被水解、糖基化、脂化和磷酸化修饰加工后以卵黄蛋白(Vitellins,Vns)的形式储存,为昆虫的卵子发育提供能量储备(Valerioetal.,2008)。在昆虫营养积累过程中,脂肪体内Vg的表达水平增加,伴随卵母细胞的发育,卵巢成熟,最后至性成熟而交配产卵(Tufail and Takeda,2008)。在昆虫胚胎发育过程中,Vg是蛋白质合成的氨基酸储备(Royetal.,2018)。有研究发现,沉默昆虫Vg表达后会影响昆虫体内营养物质的转运,缩短生殖期,导致胚胎发育受到抑制,影响昆虫产卵,甚至导致产生无效卵(Chaoetal.,2019)。
1.2 农药刺激昆虫生殖的机制
1.2.1刺激雌虫生殖的机制
药剂刺激害虫生殖的分子机制主要是影响与生殖相关的蛋白酶基因表达。它的调控机制为药剂抑制保幼激素脂酶(juvenile hormone esterase,JHE)基因的表达,但保幼激素(juvenile hormone,JHs)基因的表达量显著上调,致使成虫体内JH水平显著提高,Vns表达量显著上调。如吡虫啉处理的三化螟Scirpophagaincertulas、二化螟Chilosuppressalis成虫体内JH和蜕皮激素(20-hydroxyecdysone, 20E)水平变化显著,促进卵黄发生的JH滴度显著上升,使得二化螟、三化螟雌成虫产卵量也显著增加(Wangetal.,2005;Yuetal.,2007)。三唑磷与溴氰菊酯两种杀虫剂均能显著诱导褐飞虱雌虫交配后JHEmRNA的表达,JH III滴度随杀虫剂浓度的增加而增加(Geetal.,2010)。亚致死浓度的氯虫苯甲酰胺处理二化螟,所有发育阶段处理组中的甲基转移酶基因(Juvenile hormone acid O-methyltransferase,JHAMT)和JH水平均显著提高,表明氯虫苯甲酰胺亚致死浓度可上调二化螟JH生物合成基因的表达,进而导致JH水平升高,促进卵黄发生增加产卵量(Xuetal.,2017)。
药剂刺激害虫生殖的调控是通过激活脂肪体合成Vg的RNA转录水平开始,亚致死剂量杀虫剂会引起害虫兴奋效应,刺激昆虫Vg表达。如长期使用亚致死剂量甲维盐处理草地贪夜蛾会使其产生抗性并促进繁殖,双三氟虫脲处理甜菜夜蛾Spodopteraexigua3龄幼虫,发现羽化后2 d和3 d的雌成虫体内SeVgmRNA表达水平显著降低(杨范琴,2021;Hafeezetal.,2021)。最近,Gomaa等(2021)用病原菌芽孢杆菌Paenibacillus幼虫饲喂蜂王后发现处理后蜂王的卵巢蛋白提取量显著增加,Vg表达量更是比未处理组高20倍。亚致死剂量的三唑磷处理的褐飞虱雌虫通过实时荧光定量PCR证实了Vg在褐飞虱雌成虫中高表达。这表明杀虫剂可以通过改变Vg的表达来调控飞虱的繁殖,从而影响种群密度(Geetal.,2011;Zhouetal.,2020)。
1.2.2刺激雄虫生殖及交配传导效应
药剂除了可刺激雌虫生殖外,还可刺激雄虫生殖并通过交配传导信号给雌虫进而刺激雌虫生殖。使用三唑磷和溴氰菊酯处理褐飞虱后可导致雄性附腺蛋白(Male accessory gland proteins,MAGs)水平升高,交配时更多的MAGs传导给雌虫,使得雌成虫的脂肪体和卵巢中的卵黄素含量显著升高,且药剂处理后的Vg水平较对照组更高(Wangetal.,2010)。且与对照相比,雄虫的JH III滴度在交配前升高,交配后降低,说明JH III通过交配转移到雌虫体内(Geetal.,2010)。药剂刺激雄虫生殖的主要相关基因是精子发生相关蛋白(NISPATA5)。通过RNAi将褐飞虱雄虫NISPATA5 抑制会导致其交配对象的脂肪体和卵巢蛋白含量降低,Vg表达降低,影响卵巢发育(Geetal.,2016)。
2 内分泌激素对卵黄原蛋白影响进而对昆虫猖獗的调控作用
昆虫对生殖的调控主要是激素对卵黄发生的调控作用,其中Vg的合成和成熟的卵母细胞对Vg的吸收等过程均由激素及激素间的级联作用严密调控。过去10年的研究证明,雌性昆虫生殖主要由保幼激素、蜕皮激素、神经肽、胰岛素(insulin)以及microRNAs调控(Khalidetal.,2021)。本文主要综述激素对昆虫生殖的调控。
2.1 保幼激素对卵黄原蛋白调控的分子机制
JH诱导脂肪体中的Vg合成,促进卵泡上皮细胞间隙的开放,促进通过受体介导的内吞作用从血淋巴选择性摄取Vg,从而来调节雌性昆虫的生殖能力(Santosetal.,2019)。JH通过JH受体基因(Methoprene-tolerant,Met)作用控制卵黄发生和卵母细胞成熟。在直翅目东亚飞蝗Locustamigratoria中,RNAi介导的Met缺失可阻止JH诱导的Vg表达,降低将Vg运输到发育中的卵母细胞所需的卵泡细胞间的开放空间并阻止卵巢发育(Songetal.,2014)。同样地,在蜚蠊目太平洋硕蠊Diplopterapunctata中,RNAi沉默Met会导致初级卵母细胞发育受阻,Vg不再转录,也不再被卵巢吸收。Met还可参与调节性信息素的释放,促进性腺的发育,从而提高Vg的合成及卵巢对Vg的摄取(Marchaletal.,2017)。沉默赤拟谷盗Triboliumcastaneum中Met会导致1龄和2龄幼虫的存活率显著降低,甚至影响胚胎发育(Naruseetal.,2020)。此外,Krüppel homolo1(Kr-h1)是JH信号通路中下游关键应答基因,Kr-h1缺失会阻碍卵巢的发育,同时也显著降低产卵量(Yueetal.,2018;Heetal.,2020)。Taiman(Tai)也是bHLH-PAS转录因子家族中的一员,Met/Tai异源二聚体介导JH信号,触发Kr-h1,是诱导卵黄形成和产卵所必需的(Liuetal.,2018),沉默东亚飞蝗中的Tai-A会导致脂肪体中VgmRNA水平急剧下降到正常水平的1%,VgA和VgB转录本分别下降了91%和89%,并伴随卵巢发育和卵母细胞成熟的停滞(Wangetal.,2017),JH诱导Met与Tai结合为功能复合体,调控Kr-h1的转录。将JHAMTdsRNA注射到赤拟谷盗雌性成虫中成功地沉默JHAMT的表达后,雌虫Vg表达量显著下降,而又局部应用JH刺激后,诱导了JHAMTRNAi雌虫类胰岛素肽(Insulin-Like Peptides,ILPs)编码基因的表达,并诱导Vg基因的表达。表明JH通过胰岛素信号通路间接刺激Vg的产生(Shengetal.,2011)。
2.2 蜕皮激素对卵黄原蛋白调控的分子机制
研究表明脱皮激素(20E)可以直接或间接参与膜翅目、鳞翅目和所有双翅目昆虫Vg的产生,20E调节卵母细胞的成熟,从而间接影响脂肪体内Vg的合成。如蝶蛹金小蜂Pteromaluspuparum的卵黄发生主要受蜕皮激素调控,而JH只是起到辅助作用(Dongetal.,2007)。熊蜂BombusIantschouensis体内的卵黄原蛋白在转录水平的基因调控并且证明了在熊蜂中蜕皮激素能够诱导Vg的转录(甄丛爱等,2019)。在褐飞虱Vg的潜在分子调控机制中,蜕皮激素应答基因E74和1个重要的生殖力相关基因血管紧张素转换酶(ACE)与Vg基因表达调控有相关性。不同发育阶段的褐飞虱,其高繁殖力群体中ACE的mRNA表达水平显著高于低繁殖力群体,此外通过RNAi沉默ACE的表达,会导致Vg表达水平降低,褐飞虱的繁殖力降低。后注射E74dsRNA后,发现ACE和Vg表达水平显著降低,这表明蜕皮激素对Vg转录的调控可能是由ACE通过ACE启动子区域的E74结合位点介导的(Sunetal.,2017,2018)。20E调控级联对双翅目昆虫的卵黄发生和卵子生成至关重要。蜕皮触发激素(Ecdysis trigger hormone,ETH)是一种小型的c-末端酰胺化肽激素,由Inka细胞释放,ETH信号作为变异体激素在维持雌性昆虫正常JH水平、卵黄发生和生殖所需的JHAMT表达方面发挥着重要的功能(Ladislavetal.,2010)。人为使用20E可明显增加昆虫产卵量,在注射ETHdsRNA导致ETH、Vg和JHAMT表达量明显降低的桔小实蝇Bactroceradorsalis中,体外局部注射20E,可提高ETH和ETHR的转录,也可以增加卵子的生产(Shietal.,2019)。20E缺失不仅导致ETH水平下降,还导致JH产量减少,而ETH的沉默则导致Vg和JH生物合成基因mRNA水平下降(Khalidetal.,2021)。
2.3 保幼激素和蜕皮激素共同调控卵黄原蛋白分子合成的分子机制
果蝇的卵子发生依赖于JH和20E的平衡调控(Solleretal.,1999)。在正常情况下,JH刺激脂肪体内的卵黄蛋白合成。在卵巢中,JH与其他因素联合促进卵黄蛋白内吞,使卵母细胞发育。合成和摄取的组合作用促使成熟卵母细胞足够的卵黄沉积,维持了正常的卵黄发生进程(Yamamotoetal.,2013;Seidelmannetal.,2016)。在埃及伊蚊Aedesaegypti中,发现ETH可激活JHAMT,进而使Vg的表达显著上升(Areizaetal.,2014)。调控ETH基因表达的一个关键上游信号是20E。在桔小实蝇的RNAi实验中,ETH信号的缺失会导致JH水平急剧下降、降低Vg的合成,进而导致卵巢体积缩小、卵子产量和成熟卵母细胞卵黄沉积的减少,进而抑制雌虫的产卵量,注射JH类似物烯虫酯可使Vg表达量上升,增加产卵量。在上游注射20E,可使ETH转录,促使雌虫产卵(Shietal.,2019)。可知,20E、ETH和JH构成了部分昆虫正常生殖生理所必需的激素网络,共同调控Vg的合成。
2.4 神经肽对卵黄原蛋白的调控
激脂激素(Adipodynamic hormone,AKH)是昆虫心肌细胞合成的神经激素,能动员脂肪和碳水化合物进行包括生殖在内的能量消耗活动,AKH激脂激素受体AKHR通路参与促进卵黄发生过程(Attardoetal.,2012;Zandawalaetal.,2015)。近年,越来越多的研究表明,AKH/AKHR信号系统对雌性生殖力具有正向刺激作用(郑洪远和范书凡,2021)。如利用RNA干扰褐飞虱雌虫AKH基因,可观察到Vg在血淋巴中的积累增加,而在卵巢中的沉积减少,表明雌性生殖过程中AKH/AKHR信号系统在卵黄发生过程中起作用(Luetal.,2019)。飞蝗Locustamigratoria中AKH III主要在卵黄发生和卵成熟过程中起营养调控作用,成年雌虫每日注射AKH Ⅲ可刺激卵黄发生和卵子发育。相反,RNA干扰介导的AKHR沉默导致脂肪体中Vg表达显著降低,卵巢中Vg沉积减少。目前推测AKH/AKHR信号可能是通过三酰基甘油动员和海藻糖稳态来调节卵巢对Vg的摄取(Zhengetal.,2020)。
昆虫鞣化激素(Bursicon)是由神经系统分泌的一种异源二聚体神经肽,除对昆虫表皮鞣化、展翅等功能具有调控作用外,还参与调节Vg的表达。在家蚕BombyxmoriRNA干扰实验中,干扰Bursicon基因后,家蚕Bursicon基因表达量显著下调,其相关基因BmVg的表达水平均显著低于对照组。表明Bursicon可以参与调节Vg等生殖相关基因,从而影响家蚕的繁殖能力(杨佳鹏等,2021)。
2.5 胰岛素对卵黄原蛋白调控的分子机制
胰岛素(ILP)是一类在生物体内具有调控生长、发育、生殖等功能的肽类激素,昆虫中以类胰岛素类型存在(Nässel and Broeck,2016)。越来越多的研究证明,在昆虫卵黄发生阶段,ILP也参与昆虫Vg表达过程。埃及伊蚊ILPs可促进其卵巢产生蜕皮激素和卵母细胞吸收卵黄蛋白(Valzaniaetal.,2019)。将牛胰岛素注射到昆虫Vg合成前、饥饿或保幼激素缺失的成虫雌性体内,可以提高Vg转录和蛋白水平,表明ILPs参与了Vg合成的调控,并且其卵巢发育及繁殖能力均有显著提高(Zhangetal.,2017a)。RNAi豆荚螟虫MarucavitrataILP,幼虫表现出明显的生长迟缓,成年雌性卵巢发育被抑制(Al Bakietal.,2019)。在大草蛉Chrysopaseptempunctata中干扰PI3K通路中的关键基因Akt,中断了胰岛素信号,限制了卵巢的发育(Zhangetal.,2017b)。在东亚飞蝗中通过干扰关键信号转导因子ERK,抑制了飞蝗脂肪体中Vg的mRNA和蛋白表达,表明ERK在飞蝗卵黄发生过程中起着重要作用(李东,2018)。在赤拟谷盗雌性成虫中,JH-Met和饲喂营养物都可以诱导脑和脂肪体产生特异的ILPs。ILPs通过Akt使叉头状转录因子FOXO磷酸化,导致FOXO出核,从而引发Vg合成(Shengetal.,2011)。表明类胰岛素及其通路其不仅能直接调控昆虫生殖活动,还能与蜕皮激素和JH互作起着协同调控作用。
3 卵黄原蛋白作为杀虫剂新靶标在昆虫猖獗治理中的应用
3.1 昆虫卵黄原蛋白RNA干扰技术的研究现状
Vg是调控昆虫生长发育、繁殖和其他行为的关键基因,其合成具有性别、组织、发育期的特异性,并受激素调节,是一种研究基因表达与调控的良好模式系统。RNA干扰造成Vg合成受阻后,引起昆虫雌虫死亡率的显著升高,抑制昆虫卵母细胞成熟和卵细胞的产生,卵巢形态发育异常,昆虫的产卵量降低,进而影响种群数量的增长。表明Vg可以通过RNAi靶向控制害虫,成为害虫防治研究的重要切入点之一,亦可作为昆虫控制的精确靶点(Mitchelletal.,2019;Husainetal.,2020)。以此靶标开发新型杀虫技术,可为害虫防治提供一个新的方向。
饲喂体外合成的dsVg可显著抑制嗜虫书虱Vg的转录水平,导致卵巢发育不充分,降低产卵能力和卵孵化率(Miaoetal.,2021)。将dsVg注射到异色瓢虫Harmoniaaxyridis体内,结果显示Vg基因被特异性沉默,其相对表达量明显降低,产卵总数与孵化率低于对照组(Chaoetal.,2019)。使用显微注射法将dsRNA注射到白背飞虱Sogatellafurcifera雌虫体内,注射后的白背飞虱雌虫卵巢发育被抑制,不能产下卵粒。解剖发现,卵巢发育被抑制(邓瑶,2018)。家蚕Vg在雌虫中特别高表达,通过RNAi下调Vg基因,会影响昆虫卵的数量和颜色,干扰卵的形成和胚胎的发育,后用性类固醇17-β-雌二醇处理后,Vg在体内和体外均上调(Yangetal.,2014)。这都证明了Vg在昆虫卵的形成和胚胎发育中举足轻重的作用。Vg可作为未来进行害虫控制与防治靶标候选基因的潜力之一。干扰目标害虫的Vg,不仅可以抑制其猖獗危害问题,通过减少繁殖来控制害虫数量(Alietal.,2017)。
3.2 昆虫卵黄原蛋白沉默技术的实践意义
RNAi昆虫Vg可以通过沉默Vg基因、阻止卵巢发育、降低害虫产卵量的生殖隔断技术来防止害虫猖獗现象的发生,是一种开发周期短、操作简单、特异性高、对环境友好等优势的靶向型害虫防治技术。作为新型防治技术应用于害虫防治具有巨大潜力。但是目前对于RNAi技术的应用存在多处问题:昆虫间RNAi效率不稳定及易降解、缺乏可靠的dsRNA传递方法、脱靶和非靶标效应以及昆虫种群中潜在的抗性发展等。
针对于RNAi的沉默效率及易降解问题,相较于其他昆虫,鳞翅目昆虫RNAi效率因大部分dsRNA进入虫体会被中肠或血淋巴中的相关核酸酶降解,使其不能进入RNAi途径发挥沉默靶标基因的功能,且dsRNA被细胞吸收的效率较低,RNAi关键蛋白表达量低。目前已有用壳聚糖、脂质体、阳离子树状大分子等纳米颗粒递送dsRNA/siRNA,此技术可免受环境中核酸酶的降解,并促进其跨细胞膜转运,发挥高效的基因干扰功能,增强其细胞摄取,进而提高干扰效率。Zhang等(2010)首次用壳聚糖介导的dsRNA饲喂显著提高了冈比亚按蚊Anophelesgambiae的RNAi效率。使用涂有受体特异性配体的脂质体或无机纳米颗粒等有机纳米载体,可以保证VgdsRNA在不同环境条件下的稳定性,提高dsRNA的递送效率(王晓迪等,2020)。纳米技术与基因沉默已在其他昆虫及基因中得到研究,壳聚糖与三聚磷酸钠交联生成与dsRNA的纳米级聚电解质复合物,增大了埃及伊蚊幼虫死亡率和基因沉默效率(Dhandapanietal.,2019),以纳米粒子为载体,可有效防止dsRNA在昆虫体内被核酸酶降解,保护dsRNA安全到达靶标位点后有效释放并发挥RNAi作用。提高dsRNA的递送效率,可利用多肽转导结构域-dsRNA结合结构域(PTD-DRBD)融合蛋白,高效地将siRNA传递到细胞中(Eguchietal.,2009)。此外,在递送中,可用凝集素替代PTD,生成凝集素-DRBD融合蛋白是可行的,携带DRBD结合的dsRNA的凝集素融合可能会增强dsRNA在肠道的摄取,对棉铃虫有很好的致死效应(Ohizumietal.,2009)。
微注射可精确对靶点进行干扰但无法大规模应用于害虫防治。对于饲喂传递,使用表达dsRNA的工程菌也是一个常见的施用dsRNA的方法,可以将dsRNA构建体克隆到细菌酵母质粒中来大量生产。或用转基因植物来沉默特定害虫的Vg,增加死亡率,来阻止猖獗现象的发生(Ibrahimetal.,2017),基于工程菌高效合成靶向昆虫dsRNA的技术为田间喷洒、根灌推广提供了可行性及技术支撑。也可通过土壤灌溉和树干注射来传递dsRNA溶液(Ghoshetal.,2018)。dsRNA可被制备成用于喷雾或灌溉施用的制剂,应用在大面积的农作物上等,因此可以选择性地针对害虫(Wangetal.,2011)。但以上技术目前存在的最大障碍是安全性问题,而这需要更多的深入研究来确定其对于人类、环境和其他非靶标生物的潜在威胁。
脱靶效应会导致应用RNAi进行昆虫功能基因组学研究产生错误结论,如果利用RNAi进行害虫防治出现脱靶效应,就会因为靶标的非特异性而出现严重的环境安全性问题,甚至影响人类健康,目前还没有更好的办法去避免这个问题。害虫可以通过多种机制产生对RNA农药的抗性,包括靶基因或核心RNAi机制基因的突变、序列多态性、增强的dsRNA降解和减少的dsRNA摄取(Zhu and Palli,2020)。
4 小结和展望
近些年来,科学家们就昆虫的猖獗危害、卵黄原蛋白与昆虫的生理猖獗机制、RNA干扰技术在昆虫猖獗治理中的应用等进行了大量的研究工作,但如何将昆虫Vg作为害虫防治靶点的实际性操作技术仍有待深入探讨:1)如何解决鳞翅目昆虫对RNAi的低敏感性;2)如何使dsRNA能在不影响非靶物种的情况下,以高保真度靶向合适的转录本;3)如何解决Dicer对不同种昆虫中对dsRNA进行剪切的不同形式;4)如何做到在低成本的条件下广泛开发使用相关药剂,如昆虫生长调节剂;5)VgdsRNA技术是否可以广泛使用,会不会对生态多样性产生不可逆转的危害?
雌性昆虫生殖过程中的Vg基因作为达到有效利用RNAi技术防控害虫的目的的有效靶基因,在了解害虫猖獗机制和作为害虫防治靶点的巨大潜力受到广泛的研究,为从新型生物技术方面提供防治止昆虫猖獗及再猖獗的生物农药的新靶标奠定理论依据。通过基因编辑技术沉默害虫Vg基因,使得昆虫变态失败致死或生殖力低下,大幅度降低昆虫产卵量,有效地减少害虫数量,同时这项技术还可避免因过量使用化学杀虫剂带来的生物多样性锐减、生态平衡破坏等问题,从而减少农林业的经济损失。
猜你喜欢
热带生物学报(2022年6期)2022-11-29
中国生殖健康(2020年8期)2021-01-18
中国生殖健康(2020年2期)2021-01-18
环境昆虫学报(2020年5期)2020-11-11
中国生殖健康(2018年1期)2018-11-06
中国生殖健康(2018年2期)2018-11-06
河南畜牧兽医(2017年8期)2017-11-24
上海农业学报(2017年3期)2017-04-10
百科知识(2016年13期)2016-08-05
江苏农业科学(2014年3期)2014-07-16
