
小麦矮腥黑粉菌效应蛋白g10613的基因克隆与生物信息学分析
2023-05-12 06:22郭志浩高利刘琦
新疆农垦科技 2023年2期
郭志浩,高利,刘琦
(1.中国农业科学院植物保护研究所/植物病虫害生物学国家重点实验室,北京 100193;2.新疆农业大学农学院/农林有害生物监测与安全防控重点实验室)
小麦矮 腥黑粉菌(Tilletia controversa Kühn,TCK)所引发的小麦矮腥黑穗病是一个主要的国际检疫性病害,其对农业生产构成严重威胁[1]。这种真菌可以通过种子、土壤及空气等传播并且可以在土壤中生活很多年,一旦被引入就很难被根除[2]。
本研究利用小麦矮腥黑粉菌基因组数据克隆出小麦矮腥黑粉菌的一个效应蛋白基因g10613,对该蛋白基因进行克隆以及生物信息学进行分析,为小麦矮腥黑粉菌的后续相关研究奠定基础[3]。
1 材料与方法
1.1 试验材料
供试菌株:小麦矮腥黑粉菌菌株,由中国农业科学院植物保护研究所麦类真菌病害研究组储藏供应。
供试试剂:2×Taq 酶、2×Phanta酶购自南京诺唯赞生物科技股份有限公司。卡那霉素、Kanamycin sulfate(Biosharp)、质粒提取检测试剂盒买于北京天根生物技术有限公司,凝胶材料纯化回收检测试剂盒买于北京天根生物技术有限公司。其他试剂盒为实验室常用试剂盒。试验于2021年在中国农业科学院植物保护研究所麦类真菌病害研究组实验室进行。
主要仪器设备:FastPrep -96高通量快速样品制备仪(美国MP Biomedicals公司)、PCR仪(美国赛默飞世尔技术有限公司(Thermo Fisher Scientific))、离心机(德国eppendrof生命科学公司)、高压灭菌锅(美国zealway公司)、GHB-202恒温金属浴(浙江博日技术公司)。
1.2 试验方法
1.2.1 小麦矮腥黑粉菌效应蛋白基因的克隆
取培养基中的各阶段TCK,按试剂盒说明书方法提取RNA。再利用试剂盒将RNA合成cDNA。根据TCK基因组设计的引物数据设计全长序列引物g10613-F,g10613-R;以TCK的cDNA为模板进行了PCR扩增,反应体系包括:cDNA模板1 μL,2 × 酶12.5 μL,引物g10613-F 1 μL,引物g10613-R 1 μL,ddH2O 9.5 μL等,共25 μL。
扩增程序为:98 ℃30 s,(98 ℃10 s,55 ℃5 s,72 ℃10 s)×35个循环,72 ℃1 min。采用琼脂糖凝胶电泳验证PCR产物,跑出符合预计长度的目的片段。将目的片段用切胶处理,送到北京擎科生物技术有限公司进行检测。
1.2.2 生物信息学分析软件
运用ProtParam软件(https://web.expasy.org/protparam/)分析蛋白质理化性质,TMHMMServerv.2.0软件(http://www.cbs.dtu.dk/services/TMHMM/)预测其跨膜区,运用NCBI 中的Conserved Domains 软件(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)分析其结构域,使用SOPMA软件(https://npsaprabi.ibcp.fr/ cgi-bin/ npsa_automat.pl? page=%20npsa_sopma.html)进行蛋白二级结构分析,用Swiss-Model软 件(https://swissmodel.expasy.org/)进行蛋白三级结构建模[4]。
2 结果与分析
2.1 小麦矮腥黑粉菌基因g10613的克隆
用引物g10613-F和g10613-R(表1)扩增出约330 kb的目的基因片段。将PCR 凝胶产物回收,测序后获得330 bp长度的序列,如图1所示。
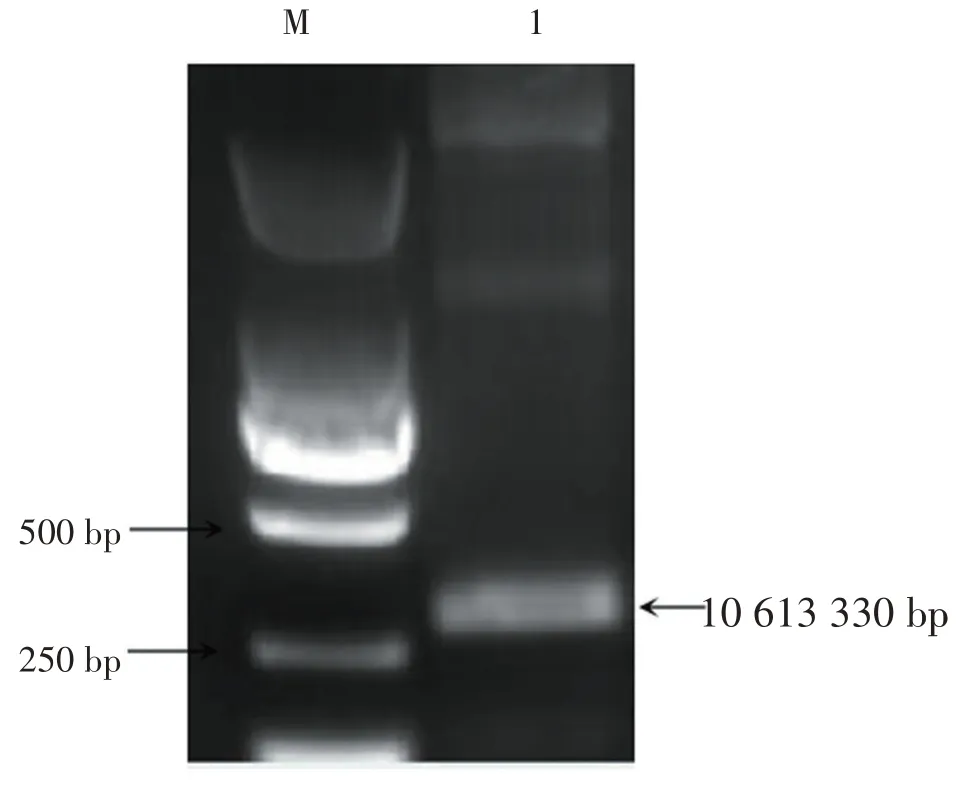
图1 基因g10613的PCR扩增产物
2.2 小麦矮腥黑粉菌效应蛋白生物信息学分析
2.2.1 效应蛋白g10613的保守结构域分析
对g10613基因进行保守结构域分析,g10613基因长度为330 bp,共编码了109个氨基酸,它没有明显的家族结构特征。
2.2.2 效应蛋白g10613理化性质分析
效应蛋白g10613的相对分子质量为11 723.80,相对理论等电点PI为9.06,说明了蛋白质的偏碱。蛋白质的总分子式是C504H826N142O150S14,分子数量是1 636。而氨基酸总数则是109个,其中其中Ala(A)8 个(7.3%)、Arg(R)3 个(2.8%)、Asn(N)6 个(5.5%)、Asp(D)6个(5.5%)、Cys(C)10个(9.2%)、Gln(Q)2个(1.8%)、Glu(E)3个(2.8%)、Gly(G)11个(10.1%)、His(H)0个(0%)、Ile(I)5个(4.6%)、Leu(L)6个(5.5%)、Lys(K)14个(12.8%)、Met(M)4个(3.7%)、Phe(F)2个(1.8%)、Pro(P)6个(5.5%)、Ser(S)4 个(3.7%)、Thr(T)8 个(7.3%)、Trp(W)2 个(1.8%)、Tyr(Y)2个(1.8%)、Val(V)7个(6.4%)。Asp和Glu的总数即负电性残体总数为9个,而Arg和Lys的总量即正电荷残体总数为17个。g10613蛋白的不稳定指数是29.78,属于稳定蛋白。脂肪指数为65.32,而总平均亲水性则为-0.252。预测该蛋白质的半衰期将是:在哺乳动物的网织红血球中超过30 h,在酵母中超过20 h,以及在大肠杆菌感染中超过10 h。
分析该蛋白的亲疏水性,最疏水性数值是3.433,而最亲水数值则为-2.844。如图2所显示,在g10613效应蛋白中亲水的氨基酸数量更多,是偏亲水蛋白质。
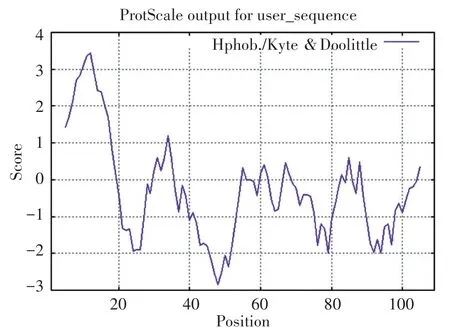
图2 效应蛋白g10613的亲疏水性图
2.2.3 效应蛋白g10613信号肽分析
效应蛋白g10613信号肽分析,切割位点C的最高值为0.851,位于第22位氨基酸;信号肽分数S的最高值为0.939,位于第2位氨基酸;合并后切割位点的Y最高值约为0.867,位于22位氨基酸。S平均值为0.811,所预测的切割位点约在1~21位氨基酸之间。如图3所示。
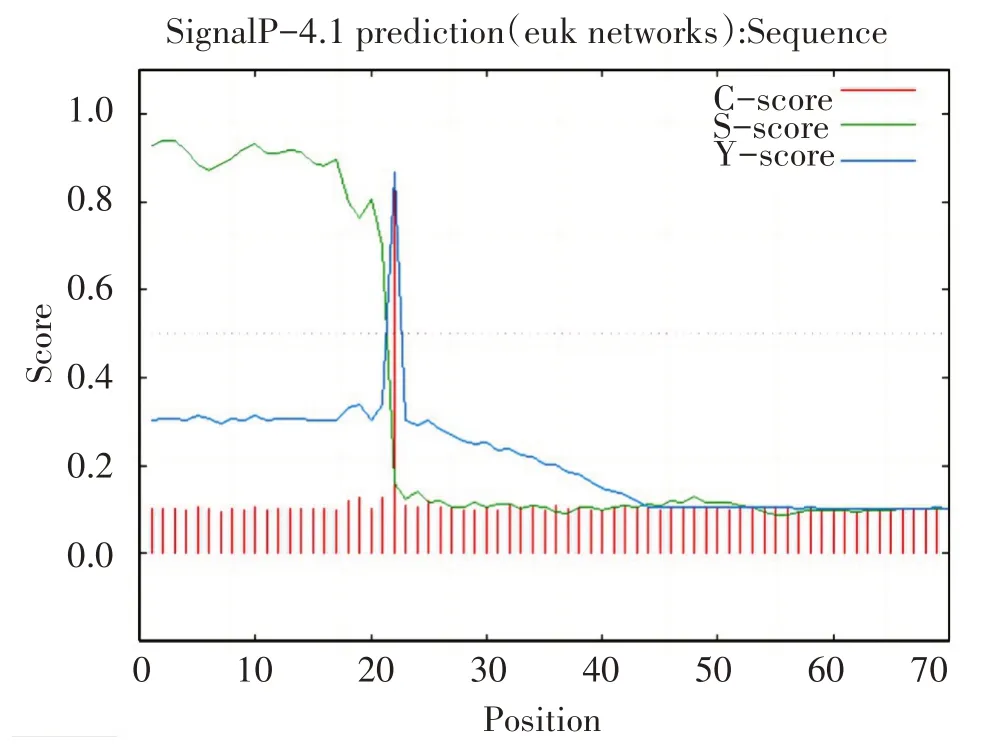
图3 效应蛋白g10613信号肽预测
2.2.4 效应蛋白g10613跨膜区分析
效应蛋白g10613跨膜区分析,通过对跨膜区的预测发现,在效应蛋白g10613的全部109个氨基酸中,存在一个跨膜区,处于0~23氨基酸的位置,如图4所显示。

图4 效应蛋白g10613的跨膜区预测
2.2.5 效应蛋白g10613二级结构预测分析
效应蛋白g10613二次结构预测分析,二次结构分析结果表明,该氨基酸具有α-螺旋、延伸链、β-转角和不规则卷曲。其中,有44个氨基酸形成α-螺旋占40.37%,7个氨基酸形成延伸链,占6.42%;5个氨基酸形成β-转角,占4.59%;53个氨基酸形成不规则卷曲,占百分之46.82%,如图5所显示。
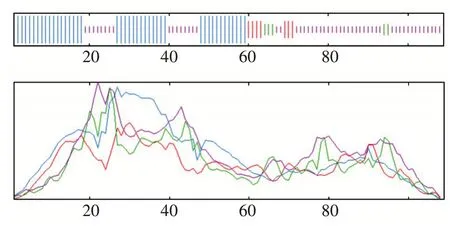
图5 效应蛋白g10613的二级结构预测
2.2.6 效应蛋白g10613三级结构预测分析
利用SwissModel 软件,对效应蛋白g10613三级结构预测,如图6。

图6 效应蛋白g10613的三级结构预测
3 讨论
小麦矮腥黑粉菌所导致的小麦矮腥黑穗病,在其通常流行的年份的发病率与发病率下降的速度大致相同。这种真菌所引起的小麦矮腥黑穗病是一种重要小麦类病害,在国际上这种真菌病害也具有相当的影响力。当环境条件适宜,有大量的感病植株时,可造成50%的产量损失,有的严重病害地块产量损失高达80%[5],甚至灭绝。但即便如此,我们对小麦矮腥黑粉菌的发病机制仍然缺乏系统深入地了解。
病原菌在侵染植物时,在与寄主细胞的相互作用的同时,会分泌许多蛋白,这些分泌蛋白被称为“效应蛋白”。研究表明,许多已经被确认的病原菌分泌蛋白质所存在的典型特点是:具有远端信号肽序列,该序列可以引导蛋白质完成跨膜运动,从而促进蛋白质被转移到细胞膜外。
由于效应蛋白是病原菌侵染植物时的重要武器,在病原菌感染植物的整个过程中起着重要的作用,因此对小麦矮腥黑粉菌效应蛋白的研究有助于阐明小麦矮腥黑穗病与小麦矮腥黑粉菌之间的互作机理。
本文成功克隆出小麦矮腥黑粉菌编码效应蛋白的g10613基因,并对该效应蛋白基因进行了克隆和生物信息学分析,但该效应蛋白的作用机制尚未清楚。本研究为下一步研究小麦矮腥黑粉菌效应蛋白的功能,深入分析基因g10613及编码的效应蛋白在小麦矮腥黑粉菌中的具体功能,以及其影响小麦植物体的方式的研究奠定了良好的基础。

猜你喜欢
环球时报(2022-09-20)2022-09-20
今日农业(2020年24期)2020-12-15
中国洗涤用品工业(2019年4期)2019-05-11
中成药(2018年1期)2018-02-02
兽医导刊(2016年12期)2016-05-17
法医学杂志(2015年4期)2016-01-06
法医学杂志(2015年4期)2016-01-06
动物医学进展(2015年10期)2015-12-07
现代检验医学杂志(2015年4期)2015-02-06
食品工业科技(2014年7期)2014-03-11
