
转录组与代谢组联合分析揭示遮阴胁迫下甘薯的代谢响应途径
2023-05-11 01:15王雁楠陈金金卞倩倩胡琳琳张莉尹雨萌乔守晨曹郭郑康志河赵国瑞杨国红杨育峰
作物学报 2023年7期
王雁楠陈金金卞倩倩胡琳琳张 莉尹雨萌乔守晨曹郭郑康志河赵国瑞杨国红杨育峰,*
1河南省农业科学院粮食作物研究所, 河南郑州 450002;2郑州大学农学院, 河南郑州 450001;3河南省农业科学院农业经济与信息研究所, 河南郑州 450002
甘薯(Ipomoea batatas(L.) Lam.)是重要的粮食、饲料、淀粉(及其制品)生产原料以及替代性生物能源作物, 我国甘薯种植面积和产量均居世界首位[1]。近年来, 甘薯逐渐餐桌化, 成为人们调节饮食结构的重要食物来源, 其种植效益也逐渐提升, 成为诸多地方乡村振兴的主推作物。在种植模式上, 甘薯经常与玉米、烟叶、芝麻、果树等进行套种以充分利用空间和地力, 获得更高的产量和经济效益。然而作为喜光作物, 甘薯在套种栽培模式中经常处于低位被遮阴, 严重影响了其产量的形成。同时, 甘薯生长中后期经常面临阴雨寡照天气而影响其干物质的积累。因此, 解析甘薯在遮阴胁迫下的代谢响应途径可为其耐荫性品种改良提供理论依据。
一般认为植物响应遮阴的策略有避荫性(shade avoidance)和耐荫性(shade tolerance)[2]。典型的避荫性反应有叶片上仰、茎伸长、减少分枝及叶片发育异常、早花等[3-4]。在遮阴条件下, 红光/远红光(R/FR)比值降低, 光敏色素变为非激活态(Pr), 导致PIFs(phytochrome interacting factors, PIF4、PIF5、PIF7)的积累, 进而激活生长素、赤霉素、油菜素内酯、细胞分裂素以及乙烯合成(或运输)途径中的E-box和G-box目标基因, 从而促进细胞伸长[5-6]。避荫性反应对于农作物的生长是不利的, 阻碍了植株生物量的扩大和籽粒产量的增加[7]。耐荫性则主要表现为比叶面积(specific leaf area, SLA)增加、叶绿素a/b比值降低以及增加光合系统II和光合系统I的比值,也就是增加碳源的获取[8-9]。当环境中的R/FR比值降低时, 耐荫性植物会通过不同植物激素的合成以及响应来抑制自身的避荫性反应, 同时还会触发一些避荫性反应的拮抗调节因子。例如拟南芥属于避荫性植物, 但遮阴条件同样会诱导其合成非典型性(非DNA结合性)的bHLH蛋白(如HFR1、PAR1、PAR2)来抵抗自身的避荫性反应[10-11]。遮阴条件在影响植物光合效率的同时, 也会导致叶片的衰老, Brouwer等[12]的研究表明, 只有强遮阴条件会诱发叶片的衰老, 弱遮阴条件下拟南芥则会进行光合作用的自适应调节以及碳源的优化分配。遮阴对农作物的品质也会产生显著影响, 番茄叶片或果实遮阴显著降低了果实中抗坏血酸的含量及其合成相关基因的表达, 进而影响了番茄果实品质[13]。黑暗遮阴后, “龙井43”茶叶中生物碱、儿茶素类及儿茶素二聚体类等物质含量显著上升, 氨基酸含量显著下降, 酚氨比显著上升[14]。
目前, 有关甘薯响应遮阴胁迫的报道较少, 王庆美等[15-16]研究表明, 遮阴胁迫使紫心甘薯块根总淀粉含量、淀粉积累速率、花青素含量显著降低, 并从酶活角度探讨了块根淀粉品质发生改变的主要原因。赵习武等[17]研究了不同遮阴处理对4种观赏甘薯光合特性的影响, 认为“黑美人”耐荫性最强。周雅倩等[18]又利用主成分分析法对6种观赏甘薯的耐弱光性进行了评价。蒋亚[19]对29个不同甘薯品种(系)进行耐荫性评价, 测定分析了其中5个不同耐荫性甘薯品种的农艺性状、光合特性、逆境生理等生理生化及产量品质指标, 深入研究了甘薯对弱光的生理响应。可以看出, 甘薯耐荫性相关研究主要集中在遮阴胁迫后甘薯的农艺性状及生理指标变化方面,缺少对遮阴胁迫下整体代谢通路变化的研究。本研究以甘薯品种郑红23号为材料, 研究了遮阴处理对其生理指标的影响, 并通过转录组与代谢组联合分析揭示了遮阴条件下甘薯的主要代谢响应途径, 为甘薯耐荫性遗传改良提供理论参考。
1 材料与方法
1.1 供试材料与遮阴处理方法
试验于2021年夏季在河南省农业科学院进行,试验材料为甘薯品种郑红23号。剪取状态一致的郑红23号大田茎尖苗8株(约20 cm, 3~4茎节), 用基质土种于营养钵内并置于室外自然光下返苗, 待植株充分返苗且生长状态旺盛时(17 d后)再进行遮阴处理。返苗后取4株(重复)采用聚乙烯遮阴网进行遮阴处理(透光率约为50%), 其余4株置于自然光下生长。遮阴15 d后进行各项生理指标测定并取叶片样品用液氮冷冻后保存于-80℃冰箱, 生理指标的测定均为4次生物学重复取平均值。
1.2 测定项目及方法
1.2.1 叶绿素含量 取主茎同叶位功能叶0.2 g,切碎成细丝后装入10 mL离心管, 加入10 mL 95%乙醇, 黑暗浸泡约48 h直至完全褪绿, 期间可摇晃试管, 此即为叶绿素浸提液。测定浸提液在665 nm(叶绿素a)及649 nm (叶绿素b)的吸光值, 空白溶液为95%乙醇。叶绿素a浓度Ca=13.95×A665-6.88×A649,叶绿素b浓度Cb=24.96×A649-7.32×A665, 色素含量(mg g-1FW)=色素的浓度(C)×提取液体积×稀释倍数/样品鲜重。
1.2.2 叶片光合及叶绿素荧光参数 利用便携式光合仪CIRAS-3测定光照和遮阴条件下郑红23号的光合相关参数。使用Pocket PEA植物效率分析仪(Hansatech)测定叶绿素荧光参数, 叶片进行暗适应20 min后, 避开叶脉使叶片在饱和脉冲光3500 μmol m-2s-1中暴露1 s后, 测定叶绿素光系统PSII最大光化学效率(Fv/Fm)、PSII潜在活性(Fv/Fo)和光合性能综合指数(PIABS)。
1.2.3 超氧化物歧化酶(superoxide dismutase, SOD)及过氧化物酶(peroxidase, POD)活性 取同叶位成熟叶样品, 使用索莱宝试剂盒(BC0175、BC0095)测定SOD和POD活性。
1.2.4 蔓长、茎节数、比叶面积、根鲜重 用尺子测量光照和遮阴条件下郑红23号植株主茎从茎基部到顶芽的长度即为蔓长, 同时测定主茎从茎基部到顶芽的茎节数。取各植株主茎同叶位功能叶,用打孔器(打孔半径0.25 cm)在其上取40个圆片,80℃烘干16 h至恒重, 比叶面积(cm2g-1)=叶片面积/叶片干重。将植株从营养钵内小心挖出, 将根系洗净晾干后称量鲜重。
1.3 转录组、代谢组联合分析
遮阴结束后取光照和遮阴条件下的叶片样品液氮冻存, 每处理包括4棵植株, 每植株取3片叶混合。由北京诺禾致源公司进行转录组测序及类靶向代谢组的分析。
采用Illumina Novaseq平台对转录组进行测序,使用HISAT2将clean reads与甘薯近缘野生种Ipomoea triloba(I. triloba)的参考基因组(http://sweetpotato.uga.edu/index.shtml)进行比对。featureCounts(1.5.0-p3)用于计算映射到每个基因的读数, 然后根据基因的长度计算每个基因的FPKM, 用DEseq2检测差异表达基因(DEGs), 用clusterProfiler (3.8.1)软件进行差异表达基因的GO富集和KEGG富集。
采用LC-MS方法对代谢组进行检测, 代谢物的鉴定基于诺禾致源自建数据库novoDB, 采用多反应监测模式(MRM)对试验样本进行检测, 采用SCIEX OSV1.4软件对色谱峰进行分析并得到代谢物定性和定量结果, 使用KEGG数据库、HMDB数据库和LIPIDMaps数据库对鉴定到的代谢物进行注释。使用R语言进行火山图绘制、聚类热图绘制、差异代谢物相关性分析、气泡图绘制。之后对差异代谢物与差异表达基因进行关联分析。
1.4 数据处理与分析
采用Microsoft Excel 2019对数据进行分析并作图, 采用SPSS19.0进行方差分析和差异显著性检验。
2 结果与分析
2.1 遮阴胁迫对郑红23号形态及生理指标的影响
从图1及表1可以看出, 郑红23号在50%透光率的遮阴胁迫下仍然能正常生长, 但是其蔓长相较于自然光照下显著增加, 茎节数则变化不显著, 说明茎节显著伸长; 遮阴胁迫下叶片变大, 比叶面积显著增加; 此外, 自然光照下的郑红23号已出现膨大块根, 而遮阴胁迫下植株根系仍以须根为主, 根鲜重显著降低。从表1同时看出, 遮阴胁迫显著提高了郑红23号叶片SOD酶和POD酶的活性。

表1 遮阴胁迫对郑红23号植株形态指标及叶片酶活的影响Table 1 Effects of shade stress on plant morphological indexes and leaf enzymatic activity of Zhenghong 23

图1 郑红23号自然光照与遮阴胁迫下的植株形态对比Fig. 1 Comparison of plant morphology of Zhenghong 23 under natural light and shade stress
从表2可知, 遮阴胁迫对郑红23号叶绿素a的含量没有显著影响, 叶绿素b以及总叶绿素含量在遮阴后均显著提高, 叶绿素a/b则显著下降。叶绿素光系统PSII最大光化学效率(Fv/Fm)、PSII潜在活性(Fv/Fo)和光合性能综合指数(PIABS)在遮阴胁迫下均显著下降。光合参数方面(表3), 遮阴胁迫显著降低了郑红23号的净光合速率和水分利用效率, 胞间CO2浓度显著提高, 蒸腾速率、气孔导度以及水汽压亏缺在遮阴胁迫下则未发生显著变化。说明, 遮阴胁迫对郑红23号的植株形态及生理指标均发生了显著影响。
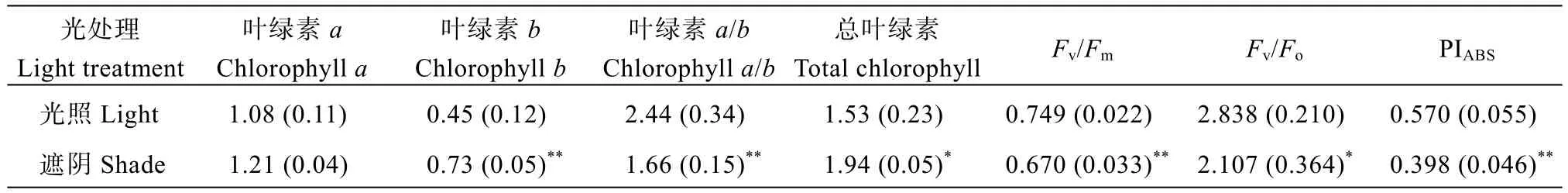
表2 遮阴胁迫对郑红23号叶绿素含量(mg g-1FW)及荧光特性的影响Table 2 Effects of shade stress on chlorophyll content (mg g-1FW) and fluorescence characteristics of Zhenghong 23
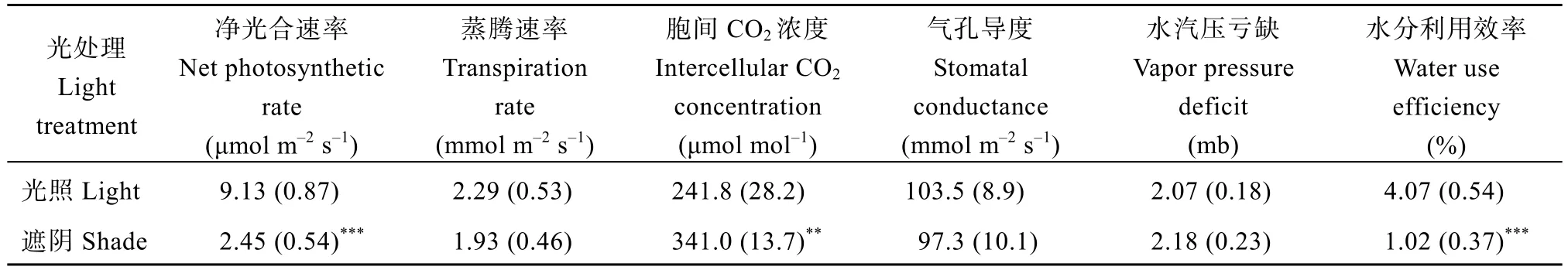
表3 遮阴胁迫对郑红23号光合参数的影响Table 3 Effects of shade stress on photosynthetic parameters of Zhenghong 23
2.2 转录组数据组装质量分析
为研究郑红23号响应遮阴胁迫的主要代谢途径, 对遮阴胁迫(shade, S)和自然光照(light, L)下的叶片样品进行了RNA-seq。结果表明, 相同生物学重复间相关性较好,R2范围在0.852~1.000; 遮阴和光照样品间相关系数R2范围在0.729~0.839 (图2-a)。8个样品RNA-seq文库获得的Clean reads同甘薯近源野生二倍体I. triloba基因组的比对率在69.64%~72.72%, 比对到单一位置的比例在67.16%~70.06%。各样品Q30碱基百分比在92.72%~93.30%。说明试验取样合理, RNA-seq数据质量可靠(表4)。
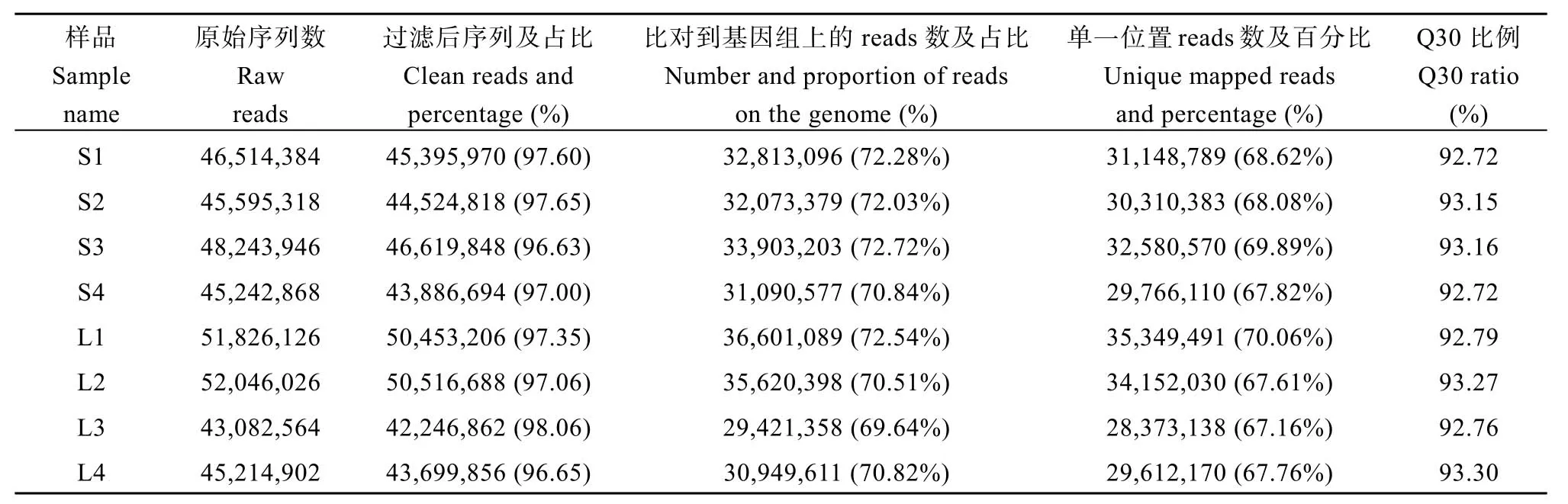
表4 样品RNA-seq质量Table 4 RNA-seq quality of samples
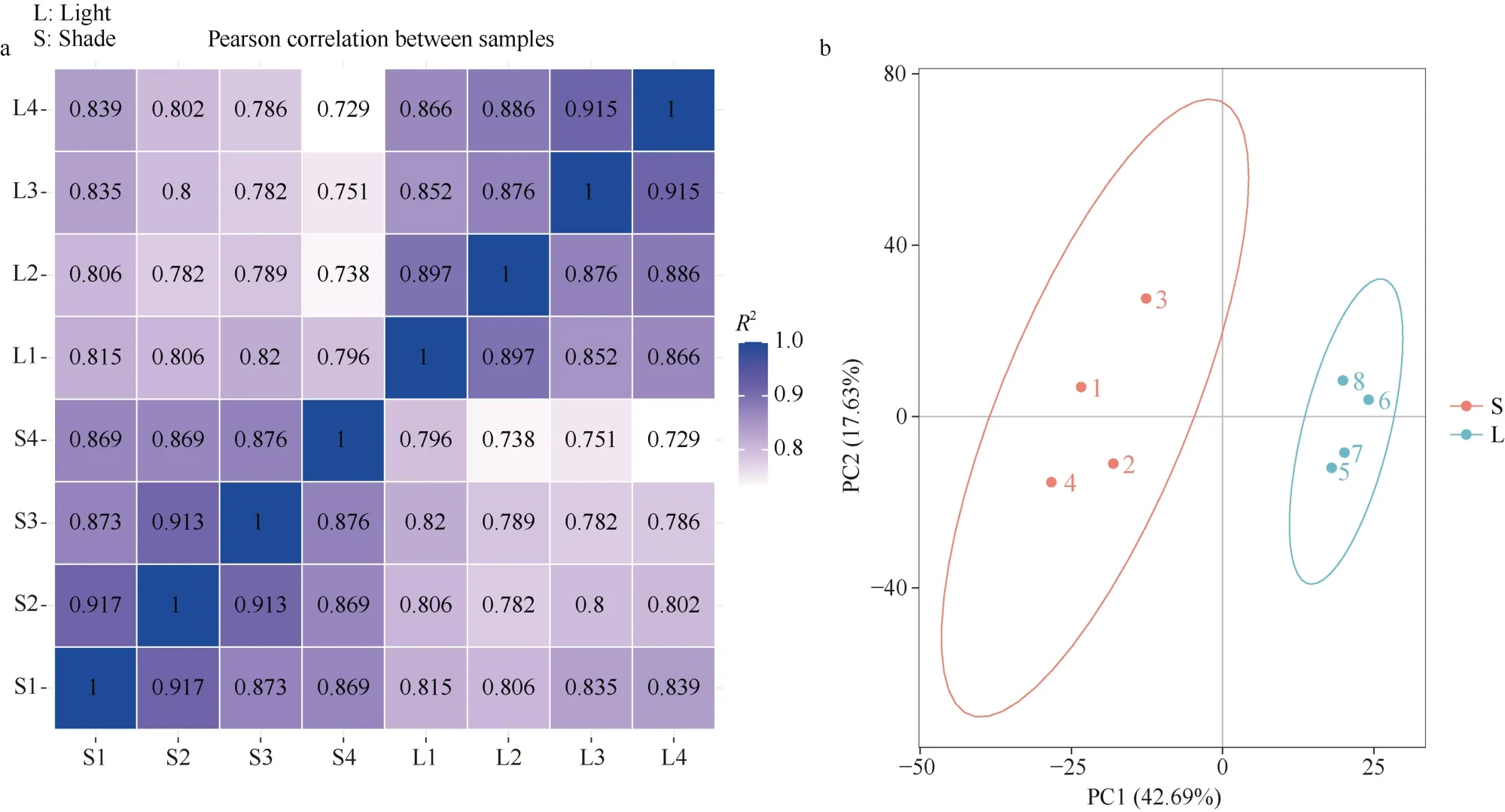
图2 RNA-seq样品相关性热图(a)及代谢组各样品间的主成分分析(b)Fig. 2 Heatmap of the Pearson correlation between RNA-seq samples (a) and principal component analysis plots of samples in metabolome (b)S: 遮阴; L: 光照。S: shade treatment; L: light treatment.
2.3 差异表达基因分析
RNA-seq结果表明, 遮阴胁迫下(S)鉴定出19,888个表达基因, 自然光照下(L)鉴定出20,050个表达基因, 其中二者共表达基因数为18,889个(图3-a)。以|log2(Fold Change)|≥1且Padj≤0.05为标准, S vs. L共筛选出2632个差异表达基因(DEGs),包含715个上调表达基因和1917个下调表达基因(图3-b)。
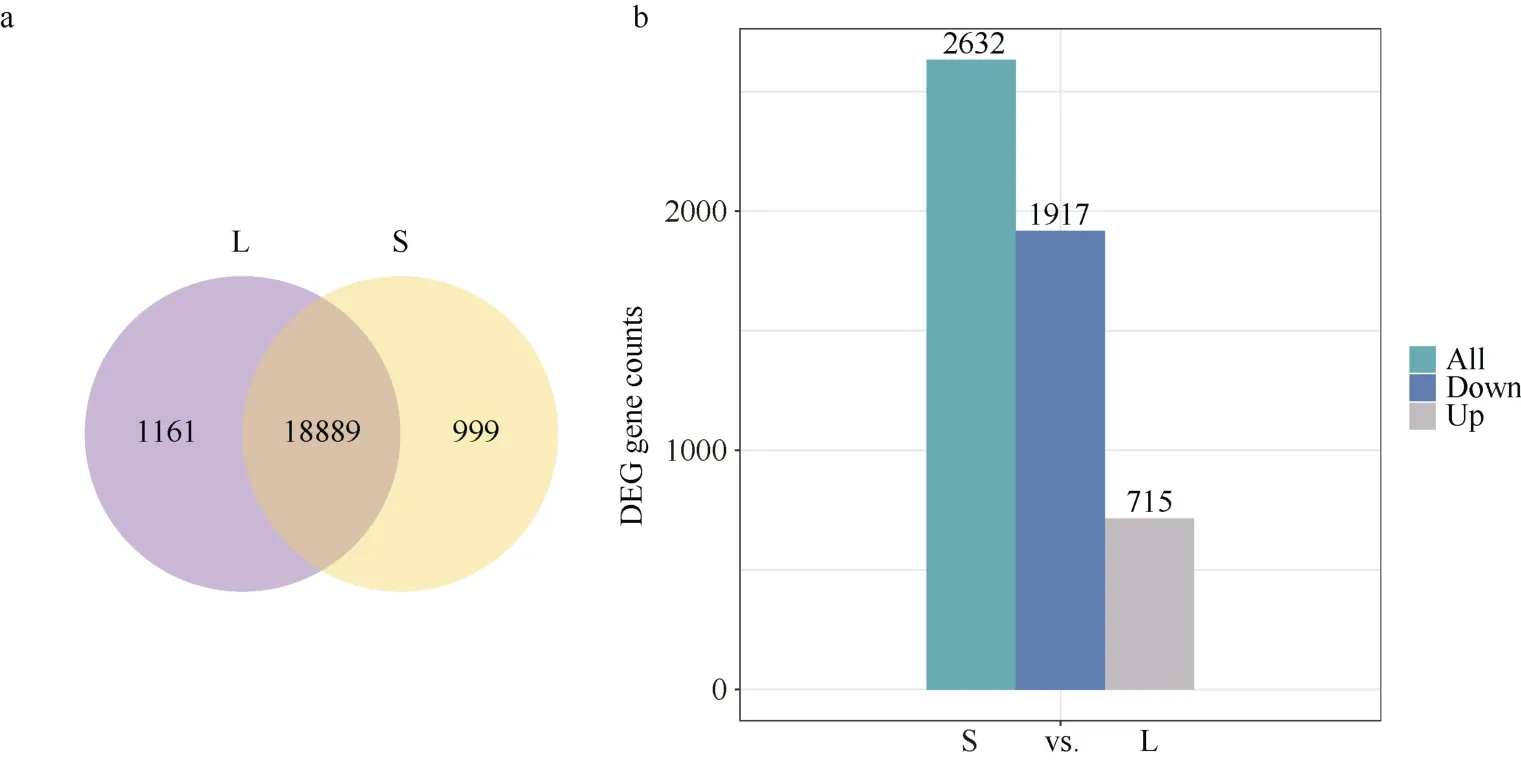
图3 遮阴胁迫和自然光照下鉴定出的表达基因数(a)和差异表达基因数(b)Fig. 3 Number of total genes (a) and DEGs (b) identified under shade stress and natural light conditionsS: 遮阴; L: 光照。S: shade treatment; L: light treatment.
对差异表达基因进行GO功能富集, 共1666个基因被显著富集于70个功能类中, 富集显著性最高的3个功能类依次为cell periphery、calcium ion binding和protein ubiquitination, 而富集差异基因数最多的功能类则为 DNA-binding transcription factor activity (81个) (图4-a)。KEGG通路富集分析显示, 富集显著性排名前3的通路依次为plantpathogen interaction、phenylpropanoid biosynthesis和pentose and glucuronate interconversions, 分别富集到57个、42个以及19个DEGs (图4-b)。富集结果说明遮阴胁迫对甘薯造成了复杂的分子生物学影响。
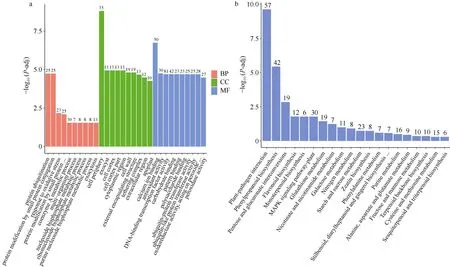
图4 差异表达基因GO富集中显著性排名前30的功能类(a)和KEGG富集中显著性排名前20的通路(b)Fig. 4 Top 30 function terms significantly enriched by GO (a) and the top 20 pathways significantly enriched by KEGG (b)BP: 生物过程; CC: 细胞组成; MF: 分子功能。柱上数字代表富集到的差异基因个数。纵坐标为显著性水平, 数值越高越显著。BP: biological process; CC: cellular component; MF: molecular function. Numbers of DEGs enriched are marked above the bars. The higher the ordinate value is, the more significant the enrichment is.
2.4 差异代谢物分析
代谢组数据表明, 在S和L组样品中共检测出1185种代谢物, 主成分分析(PCA)结果表明, 第一主成分能解释总方差的42.69%, 第二主成分能解释总方差的17.63%, 两者可将不同处理明显区分开来(图2-b)。以VIP>1.0、差异倍数FC>1.5或FC<0.667且P-value<0.05为条件筛选出差异代谢物(differential metabolite, DM) 420种, S vs. L显著上调的有113种, 显著下调的有307种。对差异显著性排名前20的DMs进行相关性分析, 结果显示不同DMs之间正(负)相关系数的绝对值在0.673~0.999,相关性较高, 说明差异显著的DMs间趋同(异)变化明显(图5-a)。KEGG通路富集显示, 差异代谢物主要富集于galactose metabolism、flavone and flavonol biosynthesis和alpha-linolenic acid metabolism, 分别富集到14个、8个以及3个DMs (图5-b)。
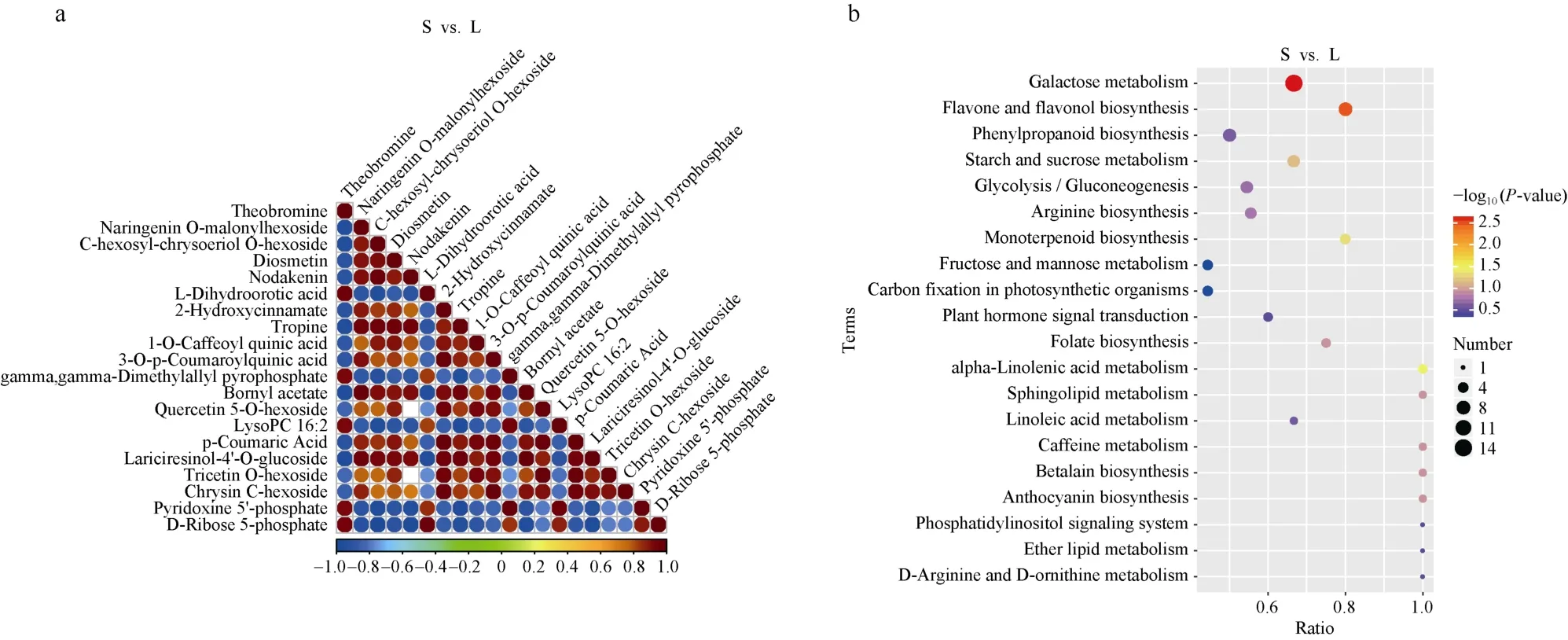
图5 Top20差异代谢物相关性图(a)及差异代谢物KEGG富集气泡图(b)Fig. 5 Correlation chart of the Top20 DMs (a) and the bubble chart of the KEGG enrichment of DMs (b)(a) 红色代表正相关, 蓝色代表负相关, 没有颜色的点表示无显著相关性(P> 0.05)。(b) 气泡颜色与大小代表富集可信度及富集到的差异代谢物数目。-log10(P-value)越大, 富集可信度越高。S: 遮阴; L: 光照。(a): the red shape indicates the positive correlation and blue indicates the negative correlation. The dot with no color means the correlation is not significant atP> 0.05. (b): the color and size of the bubble represent the enrichment reliability and the number of DMs enriched, respectively. The bigger the -log10(P-value) is, the more reliable the enrichment is. S: shade treatment; L: light treatment.
2.5 转录组与代谢组关联分析
为确定DMs与DEGs的相关性, 对二者基于皮尔森相关系数进行相关性分析, 相关系数小于0时,为负相关; 大于0时, 为正相关。图6-a展示了Top50的DMs与Top100的DEGs之间的相关性分析, 可以看出, DMs与DEGs之间更多地表现为正相关, 且同一DM与不同DEGs的相关性方向(正/负)保持一致。对DMs和DEGs分别进行KEGG富集后获得二者的共有富集通路(图6-b), 可信度排名前十的代谢通路为半乳糖、α-亚麻酸、单萜、淀粉和蔗糖、鞘脂、叶酸、精氨酸、糖酵解及糖异生、苯丙素、亚油酸。其中可能与植物逆境胁迫响应相关的途径主要有半乳糖、淀粉和蔗糖、鞘脂、精氨酸、糖酵解及糖异生、苯丙素。

(图6)
苯丙素合成途径富集到的42个DEGs共编码5种酶: beta-Glucosidase、SAM-dependent methyltransferase、peroxidase、phenylalanine ammonia-lyase、UDP-glucosyl transferase。该途径共检测到7个DMs, 除spermidine外, 其余代谢物均在遮阴胁迫下含量降低(图7-a), DEGs与DMs间的相关性均达到了显著水平(图7-b)。半乳糖代谢、淀粉和蔗糖代谢、糖酵解及糖异生均与糖代谢相关, 合并去重后3个途径共筛选到38个DEGs和19个DMs,38个DEGs共编码20种酶。19个差异代谢物中仅UDP-galactose和phosphoenolpyruvate在遮阴胁迫下含量提高, 其他均含量降低(图8-a), DEGs与DMs间的相关性也均达到了显著水平(图8-b)。鞘脂类物质是生物膜的主要结构成分, 同时参与多种信号转导途径, 在植物生长、发育和胁迫应答过程中发挥重要作用[20-21], 本研究中遮阴胁迫下鞘脂类物质磷酸乙醇胺(O-phosphorylethanolamine)和4-羟基鞘氨醇(4-D-hydroxysphinganine)含量分别是自然光照下的5.19倍和1.68倍。精氨酸是植物多胺、一氧化氮、脯氨酸、谷氨酸、氨基丁酸等的生物合成前体, 而多胺、一氧化氮则作为信号分子广泛参与植物的生长发育和逆境响应[22]。遮阴胁迫下精氨酸合成途径中的鸟氨酸、瓜氨酸和精氨酸分别是自然光照下的8.63倍、4.31倍和1.43倍, 而谷氨酰胺(L-glutamine)、精氨基琥珀酸(L-argininosuccinate)以及酮戊二酸(ketoglutaric acid)的含量则有所下降。
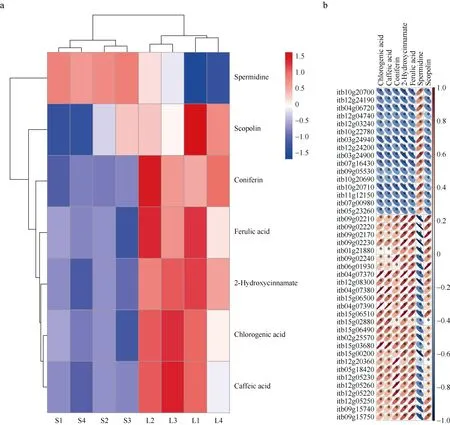
图7 苯丙素合成途径DMs的含量热图(a)以及DMs与DEGs之间的相关性热图(b)Fig. 7 Content heatmap of DMs enriched in the phenylpropanoid biosynthesis pathway (a) and correlation heatmap of the DMs and DEGs enriched in this pathway (b)
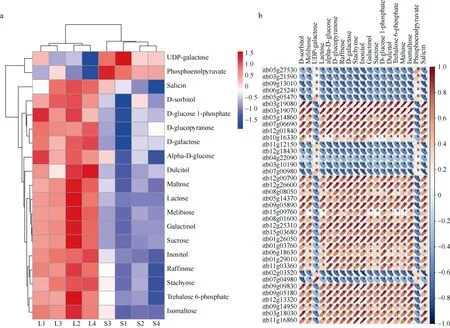
图8 糖代谢相关途径DMs的含量热图(a)以及DMs与DEGs之间的相关性热图(b)Fig. 8 Content heatmap of DMs enriched in the sugar metabolism related pathways (a) and correlation heatmap of the DMs and DEGs enriched in these pathways (b)
3 讨论
3.1 遮阴对甘薯形态及生理指标的影响
通过对甘薯品种郑红23号进行遮阴胁迫处理发现, 其蔓长、比叶面积、根鲜重、SOD和POD活性、叶绿素含量、叶绿素荧光参数、光合参数等形态和生理指标与自然光照相比均发生了显著变化。叶绿素a/b显著降低、比叶面积显著增加以及总叶绿素含量的显著增加都是植株为了获取更多的碳源而出现的耐荫性响应, 而蔓长、茎节数的增加则是避荫性反应[2,9]。一般认为, 叶绿素a主要吸收长波光, 而叶绿素b主要吸收短波光, 较大比例的叶绿素b更有利于吸收弱光中占优势的蓝紫光。蒋亚[19]研究表明, 遮阴处理显著提高了5个甘薯品种的叶绿素b及总叶绿素含量, 其中4个品种的叶绿素a含量显著提高, 叶绿素a/b则均显著降低, 这与本研究结果一致。李伟等[23]对黄瓜幼苗进行弱光胁迫后, 叶片叶绿素a、叶绿素b、总叶绿素均极显著提高; 净光合效率、气孔导度、蒸腾速率和水分利用效率显著降低, 胞间CO2浓度则显著提高; 光系统PSII中Fv/Fm呈增大趋势。而付景[24]研究表明, 玉米在遮阴胁迫下吐丝期叶片的叶绿素a、叶绿素b、总叶绿素的含量以及叶绿素a/b较自然光照均呈现无规律变化。可见, 光合系统在应对弱光胁迫时因物种及其发育阶段而异。在正常光照或强光照下, 植物叶片通常发育较厚, 比叶面积值(SLA)较低, 而低SLA值也意味着在弱光条件下单位质量的叶片具有较低的光源捕获面积。因此, 植物在弱光下会提高SLA值从而获取更多光能[25]。本研究中, 郑红23号的SLA值在遮阴胁迫下是正常光照下的2.16倍, 达到极显著差异, 说明遮阴触发了其强烈的耐荫性响应以通过光合作用获取更多碳源。然而弱光下的碳源获取仍不能支撑其根系的膨大, 遮阴胁迫下郑红23号的平均根鲜重仅为1.2 g, 而光照条件下为17.9 g,根系已明显开始膨大(图1), 说明遮阴对郑红23号的产量影响较大。
3.2 苯丙素合成途径对甘薯遮阴胁迫的响应
苯丙素合成途径以苯丙氨酸(莽草酸合成途径终产物)为起始, 涉及到的芳香类代谢物有8000余种, 光照、葡萄糖、温度、盐、干旱、病菌侵染、昆虫取食等都能调控苯丙素合成途径, 而其最主要的分支途径为木质素合成途径和类黄酮合成途径[26-27]。植物在受到盐、干旱等非生物胁迫时, 体内会积累活性氧(ROS), 而ROS会对植物造成二次伤害。转录组联合代谢组分析表明, 苦豆子在受到盐胁迫后会触发苯丙素合成途径中木质素类和类黄酮类物质的合成, 二者会参与到根部ROS的清除[28]。超氧化物歧化酶(SOD)和过氧化物酶(POD)是植物体内重要的抗氧化酶, 在植物受非生物胁迫时其活性会提高从而起到清除体内活性氧、维持氧化还原平衡的作用[29-30], 而POD也是木质素合成的关键调控酶。本研究转录代谢联合分析表明, 苯丙素合成途径中富集到的上调差异表达基因多数为POD酶家族基因, 同时, 郑红23号在遮阴胁迫下的SOD活性和POD活性均显著高于自然光照条件(表3), 说明遮阴胁迫使郑红23号体内积累了ROS并触发了活性氧清除系统。亚精胺(spermidine)是植物体内一种常见的多胺物质, 具有重要的生理活性, 广泛参与植物逆境胁迫响应(盐、干旱、机械损伤、低温、水淹、重金属), 其可通过激活抗氧化酶活性(SOD、POD、CAT)来参与调节胁迫条件下植物体内的活性氧稳态[31-32]。同时, 外施多胺类物质也可提高植物对非生物逆境的抗性[33-34]。本研究中, 遮阴胁迫下亚精胺的平均含量是自然光照下的2.22倍, 说明亚精胺可能通过提高植株的抗氧化能力来参与甘薯的遮阴胁迫响应。
3.3 糖代谢相关途径对甘薯遮阴胁迫的响应
糖代谢不仅是植物的能量来源也为其提供了重要的结构物质组成元件, 许多种类的糖还能与蛋白质等结合成复杂的化合物(如糖蛋白)而参与到细胞识别和细胞间物质运输等生命活动, 调节植物生长发育和应对不良环境[35-36]。转录组DEGs和代谢组DMs的KEGG共有富集途径中排名靠前的有3个和糖代谢相关, 分别为半乳糖代谢途径、淀粉和蔗糖代谢途径、糖酵解和糖异生途径, 3个途径富集到的38个DEGs中有26个在遮阴胁迫后下调表达, 19个DMs中有17个在遮阴胁迫后含量降低, 说明遮阴胁迫降低了甘薯植株的糖代谢水平。其中11个下调DEGs编码了与淀粉代谢直接相关的8种酶, 与淀粉合成相关的为 PGM (EC 5.4.2.2)、ADPG (EC 2.7.7.27)、SSS 1/3/4 (EC 2.4.1.21)以及GBSS (EC 2.4.1.242), 与淀粉降解相关的为α/β-Amylase (EC 3.2.1.1/EC 3.2.1.2), 说明遮阴胁迫对甘薯的淀粉合成与降解均有抑制作用。此外, 蔗糖、葡萄糖、麦芽糖、乳糖、棉籽糖等重要的可溶性糖在遮阴胁迫下的含量均不同程度下降。遮阴胁迫下叶片糖代谢水平的下降也直接导致了向根部运输的碳同化物的减少以及块根膨大受阻(图1)。而对玉米的研究也表明, 弱光胁迫下, 蔗糖-淀粉转化相关酶活性降低,淀粉合成不足, 籽粒充实度较差, 导致产量显著降低[37]。甘薯是喜光作物, 遮阴对其糖代谢产生了负面影响, 而对于某些喜阴植物如胡椒, 当遮阴度为30%左右时, 可以促进糖积累, 为花芽发育提供充足的物质基础[38]。同为地下根茎类植物, 中药材白及在强光环境下会受到胁迫, 影响产量和品质, 而50%遮阴处理则会提高白及叶片光合能力和糖物质积累, 块茎产量显著提高, 蛋白组学显示遮阴和光照条件下的差异蛋白主要富集于光合作用、碳代谢、糖酵解/糖异生、磷酸戊糖途径等[39]。这说明遮阴胁迫对喜光植物和喜阴植物的糖代谢途径造成了相反的结果, 而这其中的关键调控因素则有待进一步研究。
3.4 鞘脂及精氨酸代谢途径对甘薯遮阴胁迫的响应
鞘脂构成了植物细胞膜至少40%的脂质成分,而植物内膜系统(如内质网、液泡)同样富含鞘脂[20]。植物鞘脂主要包括鞘氨醇即长链基团(LCBs)、神经酰胺、葡萄糖神经酰胺以及糖基肌醇磷酸神经酰胺。鞘脂代谢广泛参与到植物对生物或非生物胁迫的响应。鞘氨醇含量的增加能够增强拟南芥的耐盐性,而鞘氨醇-1-磷酸(LCB-1P)能参与调节植物气孔的开闭, 并参与到脱水、低温胁迫响应[40-43]。4-羟基鞘氨醇也即植物鞘氨醇(phytosphingosine)是植物中主要的鞘氨醇长链基团, 干旱胁迫下, 植物鞘氨醇含量在耐旱型樱桃砧木中显著提高, 可作为樱桃砧木的抗旱标志代谢物[44]。本研究中4-羟基鞘氨醇在遮阴胁迫下的含量是正常光照下的1.68倍, 说明遮阴胁迫诱导了甘薯叶片中膜脂成分的变化, 从而使植株更加适应遮阴环境。同时, 遮阴胁迫下磷酸乙醇胺的含量是自然光照的5.19倍, 而磷酸乙醇胺是植物生物膜组分磷脂酰胆碱的合成前体, 后者对维持生物膜的功能有重要作用[45]。以上结果说明甘薯植株能通过改变生物膜脂质成分来响应遮阴胁迫, 从而提高其在遮阴胁迫下的生存能力。
精氨酸及其代谢产物鸟氨酸是一些逆境相关信号分子如多胺、脯氨酸、谷氨酸等的合成前体物质,因此, 精氨酸代谢通路在植物逆境信号转导中发挥重要作用[46]。精氨酸可在精氨酸酶的催化下单向水解生成鸟氨酸和尿素, 鸟氨酸进而可生成瓜氨酸,而瓜氨酸经过两步催化又可生成精氨酸。同时, 精氨酸酶是植物精氨酸代谢的关键酶之一, 其基因主要通过JA和ABA途径参与响应盐、干旱、病害等(非)生物胁迫[47-49]。本研究中, 遮阴胁迫下鸟氨酸、瓜氨酸和精氨酸分别是光照条件下的8.63倍、4.31倍和1.43倍, 多氨类物质精氨是光照条件下的1.51倍,而精氨酸酶基因的表达量则是光照条件下的1.91倍,说明遮阴胁迫诱导了精氨酸酶基因的表达并将精氨酸更多地催化生成鸟氨酸, 而鸟氨酸可能会进一步参与到多胺类抗逆因子的合成中从而使植株增强对遮阴胁迫的适应能力。
4 结论
本研究以郑红23号为材料通过转录组和代谢组联合分析首次解析了遮阴胁迫下甘薯的代谢响应途径。遮阴胁迫下郑红23号的植株形态、叶片光合能力、叶绿素含量、抗氧化酶活性以及块根膨大均受到了显著影响。转录组和代谢组联合分析表明,苯丙素合成途径、糖代谢相关途径、鞘脂和精氨酸代谢途径是甘薯响应遮阴胁迫的主要代谢途径。苯丙素途径中的POD酶家族基因被广泛激活, 提高了植株遮阴胁迫下的抗氧化能力。同时, 遮阴胁迫降低了叶片糖代谢水平, 淀粉的合成和降解均受到抑制, 可溶性糖含量下降, 导致了向根部运输的碳同化物的减少以及块根膨大受阻。而鞘脂及精氨酸代谢途径则可能通过提高生物膜的稳定性以及增加多胺类抗逆因子的合成底物来使植株更好地适应遮阴胁迫。以上结果为理解遮阴胁迫下甘薯的代谢响应途径提供了新的理论依据。
猜你喜欢
中国机械工程(2022年8期)2022-05-09
中国机械工程(2021年8期)2021-05-07
音乐教育与创作(2019年8期)2019-05-16
今日农业(2019年16期)2019-01-03
西南农业学报(2016年5期)2016-05-17
中外医疗(2015年11期)2016-01-04
中国当代医药(2015年30期)2015-03-01
河南科技(2014年4期)2014-02-27
当代畜禽养殖业(2014年7期)2014-02-27
河南医学研究(2012年3期)2012-08-15
