
水稻株型生理生态与遗传基础研究进展
2023-05-11 01:15铨徐正进陈温福
作物学报 2023年7期
许 娜 徐 铨徐正进 陈温福
沈阳农业大学水稻研究所, 辽宁沈阳 110866
1 水稻株型的概念
早在1923年, Engledow等[1]就提出聚合作物控制产量优异性状的设想, 后来Heath等[2]认为可以通过改变叶形和受光姿态等改变作物生产能力。在此基础上, 1953年门司正三等[3]引入群体结构和冠层消光系数概念, 为作物相关研究奠定了理论基础。1959年角田重三郎等[4]首先根据对水稻、大豆和甘薯的研究, 提出了耐肥性与株型的关系及不同肥水条件下茎叶同化系统的理想型。Donald[5](1968)指出作物应具有充分利用自身有限空间而不侵占相邻植株生长空间的特点, 进一步丰富发展为作物理想株型(ideotype)。20世纪70年代, 松岛省三[6]从高产栽培角度建立了理想稻群体与个体形态生理指标。杨守仁等认为株型可分为狭义株型(plant morphology)和广义株型(plant type), 狭义株型指植物体的形态特征及空间排列方式, 广义株型还包括与群体光能利用直接相关的生理、生态方面的机能性状, 而理想株型是指在特定生态条件下与丰产性有关的各种有利性状的最佳组配[7-8]。
2 水稻株型相关研究回顾
2.1 矮化育种与株型
20世纪50年代末60年代初的矮化育种是株型改良的初始阶段, 黄耀祥等育成以桂朝2号为代表的一批产量潜力超过7.5 t hm-2的矮秆品种[9], IRRI育成了产量潜力接近10 t hm-2的半矮秆高产品种IR8[10]。以利用矮秆基因semidwarf 1(sd1)为基础进行的矮化育种, 大幅度提高了水稻的单产潜力, 被称为“绿色革命”。矮化育种通过降低株高、提高抗倒伏性和经济系数, 使水稻产量比原有农家品种提高了20%~30%, 实现了水稻产量的第一次飞跃[11]。受角田、Donald和松岛等株型相关研究结果启发,水稻株型改良从初期侧重茎秆性状的矮化育种, 逐步进入茎秆、叶片、穗部性状乃至根系性状综合改良的理想株型阶段[7-8]。
2.2 籼粳稻杂交育种与株型
亚洲栽培稻分为籼和粳两个亚种, 二者形态生理生态特性有明显差异, 农艺性状各具优缺点而且有很强的互补性, 基因组也发生了深刻分化,中国是世界上唯一同时大面积种植籼稻和粳稻的国家[12]。因此, 在中国籼粳稻杂交育种的必要性和可行性无需论证。麦类穗本身光合作用不容忽视,而水稻穗通常被认为是非光合器官[13]。典型粳稻抽穗后不久穗开始逐渐弯曲, 剑叶虽直立但是较短,穗颈一般细而长并随穗弯曲, 穗位于群体冠层最上部, 因此一是穗遮光恶化冠层生态环境影响群体光能利用, 二是重心高不利于抗倒伏, 三是灌浆进程中穗和穗颈弯曲可能诱发倒伏(图1-A和图2)[14]。典型籼稻穗长而下垂, 穗颈短粗, 剑叶长而直立,实现了“高冠层、矮穗层”, 既有利于改善冠层生态环境又促进群体光能利用, 还降低重心增强了抗倒伏性(图1-B)[15], 而直立穗型粳稻则克服了典型粳稻株型弊端(图1-C), 实现了既有利于群体光能利用又抗倒伏, 与典型籼稻一样具有较为合理的冠层结构和受光态势[13-14]。
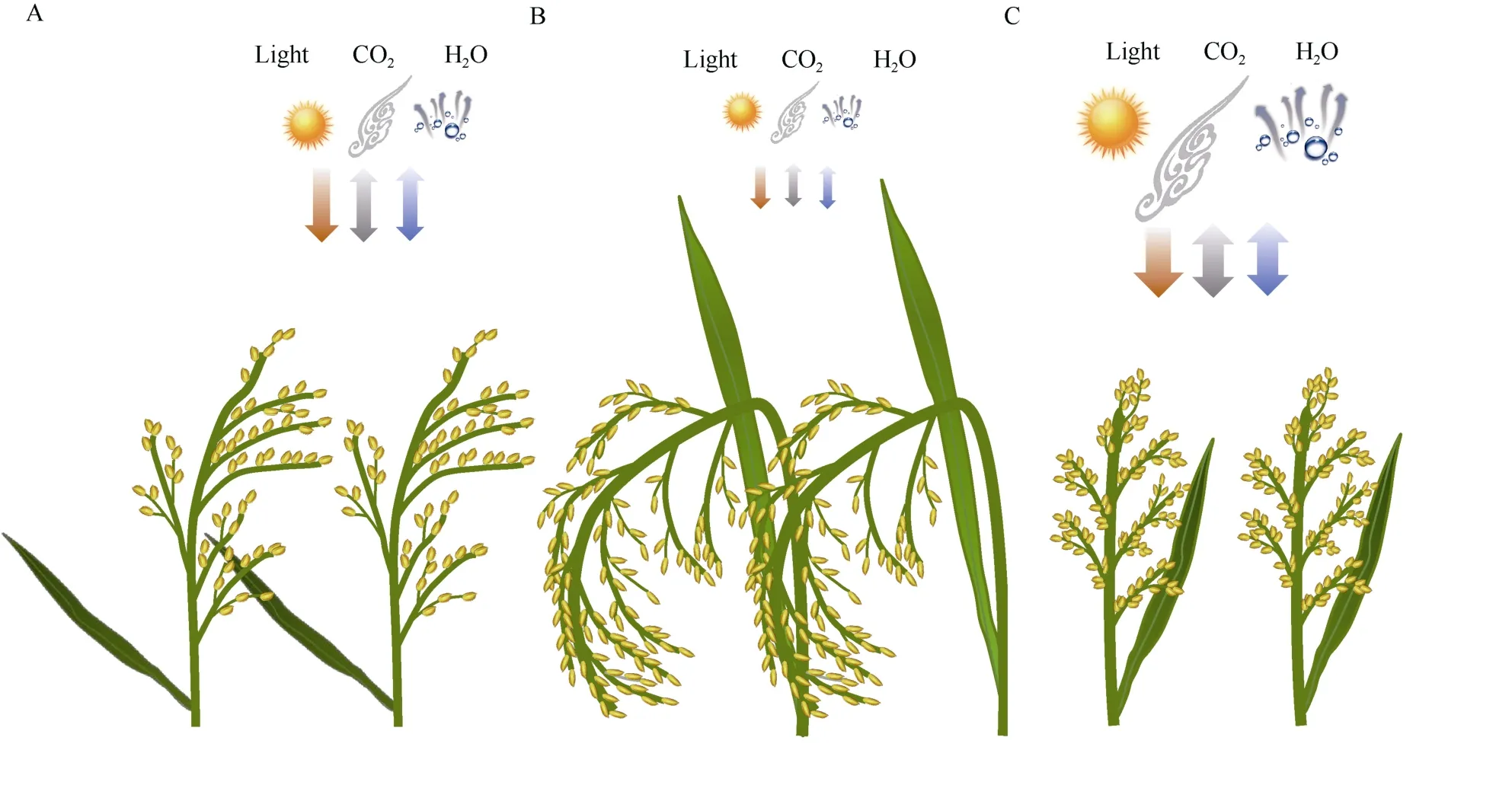
图1 典型粳稻(A)、典型籼稻(B)和直立穗型粳稻(C)群体生理生态特性比较Fig. 1 Comparison of physiological and ecological characteristics among the typical GJ rice variety (A), the typical XI rice variety(B), and the erect panicle architecture GJ rice variety (C)图标和箭头大小表示太阳辐射、H2O和CO2扩散强度高低The size of icons and arrows indicates the diffusion intensity of solar radiation, H2O and CO2. GJ:geng/japonica; XI:xian/indica.
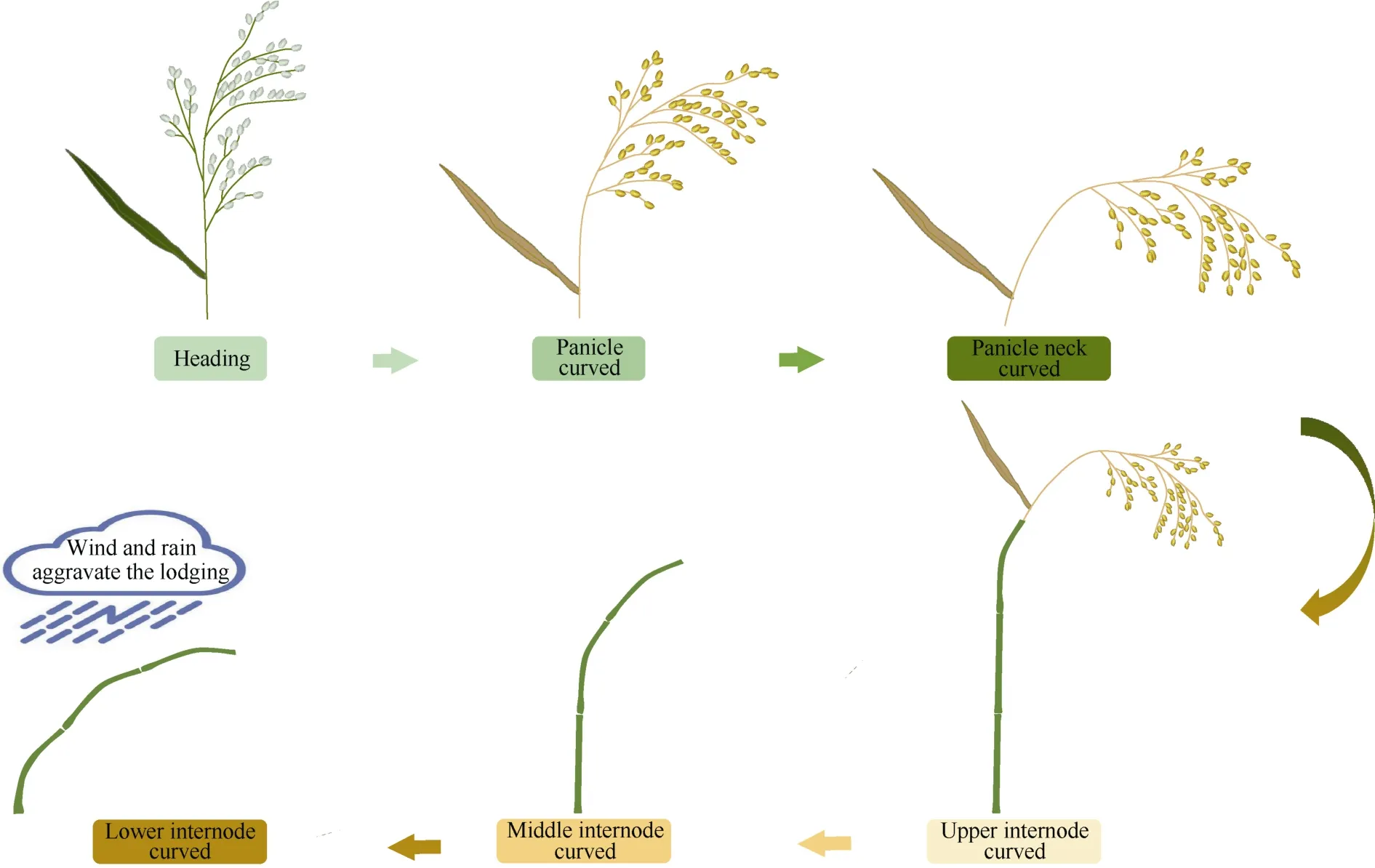
图2 典型粳稻穗弯曲诱发的倒伏Fig. 2 Lodging process induces by panicle curvature in typical GJ rice variety
2.3 超级稻与株型
日本于20世纪80年代初率先开展水稻超高产育种研究[16]。1989年IRRI也开始了水稻新株型超高产育种(new plant type, NPT), 后来被媒体广泛宣传为超级稻(super-rice)[17]。国内分别于1996年和1998年启动了“中国超级稻研究”和“超级杂交稻育种”计划[18]。纵观国内外超级稻研究, 基本都是以改良株型为主体技术路线, 日本是利用籼稻株型和粳稻光合特性, IRRI一开始就叫“新株型”, 国内常规稻是“理想株型与优势利用相结合”, 杂交稻是“理想株型与杂种优势利用相结合”, 并提出了北方粳稻直立大穗型模式、杂交籼稻超高产模式、华南半矮生早长模式、西南亚种间重穗型模式等[9,14,18]。
3 水稻株型生理生态研究
3.1 根系性状
水稻根系是水分和养分吸收的主要器官, 也是多种氨基酸、激素及有机酸合成的重要场所, 同时对地上部分起到支撑固定作用, 并且与抗逆性有密切关系, 是直接影响产量、品质和适应性的重要的株型性状[19-20]。作为多基因控制的数量性状, 根长、根粗、根数、根体积、根表面积、根干重等指标相互联系影响, 遗传机制复杂且易受环境条件影响,由于研究方法限制难以准确快速鉴定表型, 因此水稻根系生理生态研究相对滞后[19-20]。大量研究表明,水稻根系多项指标大多与产量及其构成因素正相关, 但是不同层次根系与地上部的关系及其功能却有所不同, 根系分布多纵向且较深时, 叶片趋向于直立[21]。穗数型和矮秆多穗型品种根较细而短, 多横向生长处于土壤浅层, 而穗重型和高秆大穗型品种根较粗而长, 多纵向生长处于土壤深层[22]。生育前期形成的下层根对足穗、大穗有积极作用, 与穗分化同步形成的上层根与结实率和粒重关系更密切。总体上说, 深层根比例大更有利于品质改善和产量提高, 大穗型品种这种趋势尤其明显[23]。上述研究都是以地上部分和根系表型有显著差异的品种为试材, 遗传背景及其他性状等影响错综复杂, Uga等[24]用近等基因系研究水稻根系功能证明, 深层根有利于提高氮素利用率、延缓地上部分衰老, 通过结实率和千粒重显著增加产量。
3.2 茎秆性状
株高是最重要的茎秆性状, 也是影响抗倒伏性的关键因素。矮化育种的核心是降低株高, 提高抗倒伏能力和经济系数进而产量潜力跃升, 但是矮秆高产品种氮肥响应减弱、利用效率低的问题逐渐显现[25]。矮秆高产品种经济系数已经达到0.5~0.6, 在此基础上进一步追求高产, 可能依赖于生物产量的提升或与经济系数的优化组配[7]。针对这一问题研究发现, 矮秆高产品种适当增加株高, 可以降低叶面积密度(单位冠层高度的叶面积), 有利于群体CO2扩散, 进而提高了最适叶面积指数和群体光合速率。这可能是株高一般与生物产量正相关的生理生态机制之一, 寻求利用株高以外性状提升抗倒伏性, 就成为矮化育种后持续增产面临的新问题[18]。从另一个角度分析, 增加株高改善群体结构的关键是叶片空间分布, 主要由剑叶长和中下部节间长决定, 与倒一节间特别是穗颈长无关, 粳稻穗颈像籼稻那样缩短长度增加粗度, 既不影响受光态势又降低穗位还减少穗对茎秆弯曲力矩, 有效协调了株高、抗倒伏性和生物产量的矛盾(图1)[18]。影响水稻倒伏的因素很多而且有复杂的相互作用, 其中与茎秆性状关系最密切, 相关研究进展近期已有全面综述[26-27]。分蘖角度与群体结构和受光态势有直接关系, 前期分蘖角度较大有利于减少漏光损失同时抑制杂草生长, 中后期分蘖角度缩小促进群体通风透光并且增强抗倒伏性[28]。
3.3 叶片性状
水稻产量的90%来自叶片光合作用, 叶片光合作用取决于单叶光合特性在群体条件下的综合作用。在单叶或个体层次, 光合速率与叶片长、宽、角度等形态性状关系并不密切, 通常与气孔密度和比叶重(叶片单位面积干物重、是氮和叶绿素含量、RuBP羧化酶含量及活性的间接指标)显著正相关,将籼稻的高气孔密度与粳稻的高比叶重相结合, 是提高水稻叶片光合速率的可行途径(图3)[29]。相对玉米等C4作物而言, C3作物水稻具有更强的弱光适应性, 在群体条件下可以通过叶片直立牺牲其受光强度, 降低消光系数(K)提高最适LAI获得更高的群体光合速率[18]。角田认为耐肥高产品种理想型表现为叶片厚、小、直立而深绿色[4], Donald理想株型的核心是叶片小而直立[5], 松岛从栽培角度提出高产品种应该具有短、厚、直立的叶片[6], 袁隆平超级杂交籼稻的主要特点是叶片窄、凹、直、长、厚[15], Khush[30]认为叶片浓绿、厚且直立的叶形更有利于高产, 共同目标都是追求叶片直立降低消光系数。比较分析可以发现, 超级杂交籼稻叶片长度取向与其他理论迥异。究其原因: 一是五六十年代之交正是高秆农家品种向矮化育种转换之际, 矮秆高产品种耐肥抗倒; 二是日本粳稻叶片一般较短, 栽培不当剑叶过于伸长往往减产; 三是相对常规品种特别是北方粳稻常规品种, 超级杂交籼稻穗数较少而叶片和穗较长, 穗弯垂、叶片长而直立有利于保证足够的叶面积指数, 充分发挥上部叶片特别是剑叶对产量的贡献。
3.4 穗部性状
穗部性状是与产量和品质直接相关的重要复杂农艺性状, 包括穗长、一次枝梗数和粒数、二次枝梗数和粒数及其分布、每穗粒数、着粒密度、粒形、粒重等, 下面分为穗型和粒型分别评述。
迄今提出了多种穗型分类方法, 以形态特征为指标的主要有: 依据穗直立程度的直立、半直立和弯曲穗型; 依据着粒密度的密(紧)、半密(紧或散)和稀(散)穗型; 依据二次枝梗籽粒着生方式的二次枝梗上位、中位和下位优势型; 依据穗颈与倒二节间大维管束关系的穗数、偏穗数(重)和穗重型等[31]。有关直立穗型的生理生态特性已在本文2.2中评述。一般认为着粒密度大是一种高产性状, 但是通常对结实性和品质性状表现负效应[32]。北方粳稻穗较短,着粒密度已经达到较高水平, 尽管着粒密度仍然与产量显著正相关, 但是为兼顾加工和外观品质, 不宜再增加着粒密度[33]; 籼稻穗较长, 着粒密度较低,进一步增加着粒密度可能是高产优质的可行选项(图4)[34]。受一次枝梗数和每个一次枝梗着生粒数的限制, 每穗粒数主要由二次枝梗粒数决定, 二次枝梗特别是穗轴中下部二次枝梗籽粒(弱势粒)结实性较差, 因此每穗粒数通常与结实率负相关。穗型指数(panicle type index, PTI)是二次枝梗粒数最多的一次枝梗所在穗轴节位与一次枝梗数与之比, 反映二次枝梗籽粒在穗轴上分布趋势, PTI较高即二次枝梗籽粒偏向穗轴上部分布的二次枝梗上位优势型, 有利于缓解结实率随每穗粒数增加而降低的矛盾, 也是降低直立密穗型粳稻强势粒和弱势粒外观、加工及食味品质性状差异的可行途径[35]。籼粳稻茎秆大维管束数差异不大, 但是籼稻大维管束效率(穗颈与倒2节间大维管束数之比)高于粳稻, 而一次枝梗效率(一次枝梗数与穗颈大维管束数之比)小于粳稻,粳稻每个一次枝梗效只有1个大维管束, 而籼稻基部一次枝梗效有2个大维管束(图3)[18], 因此籼稻增加一次枝梗数的潜力大于粳稻。
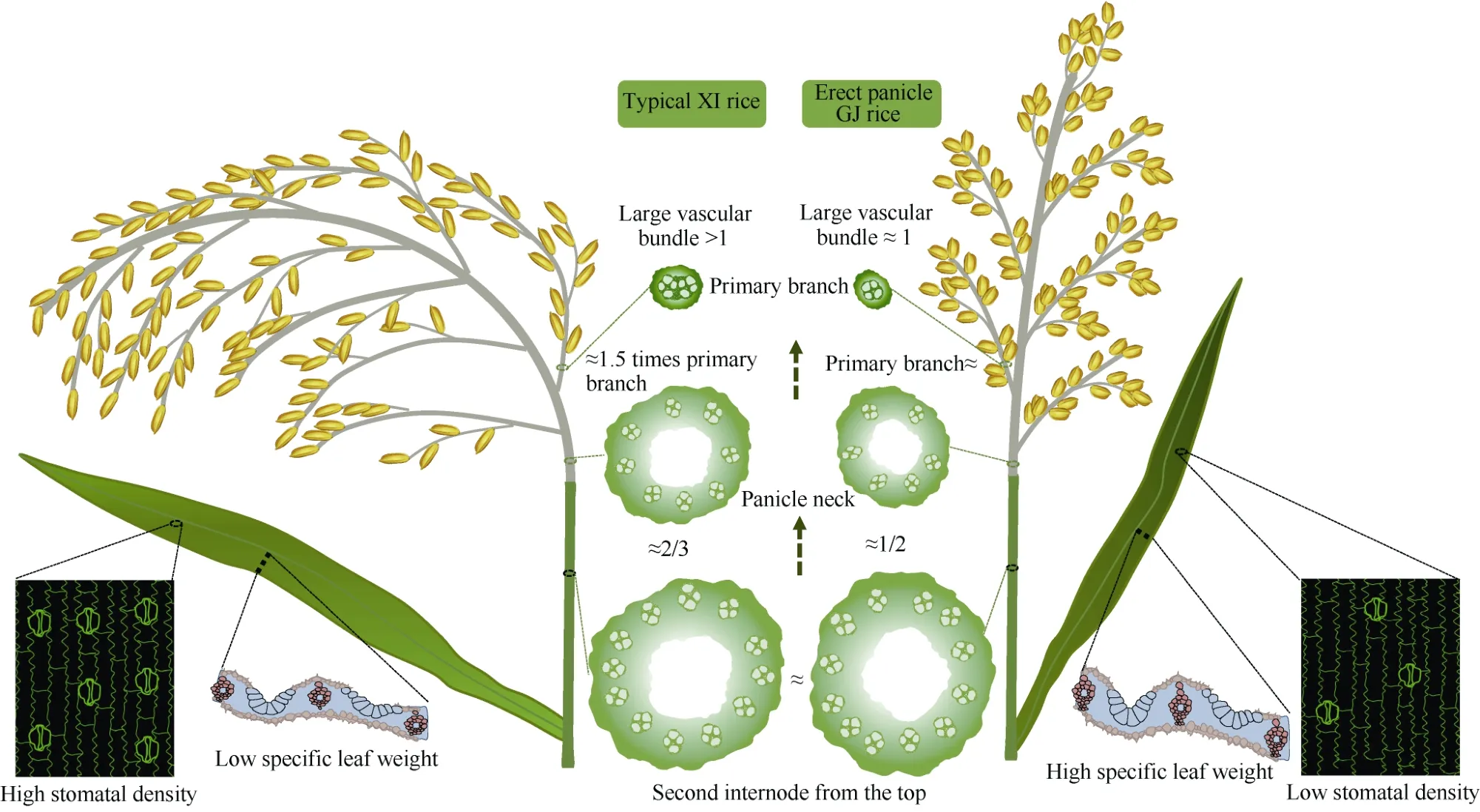
图3 典型籼稻(左)和直立穗型粳稻(右)形态解剖特性Fig. 3 Morphological and anatomical characteristics of typical XI rice variety (left) and erect panicle architecture GJ rice variety(right)
粒型包括粒长、粒宽、粒厚、长宽比(粒形)、长厚比及粒重。粒形即籽粒长宽比是最重要的粒型性状,是粒型分类的指标, 也是籼粳亚种重要差异性状; 总的趋势是在一定范围内籽粒长宽比大有利于提高外观品质, 籽粒长宽比小有利于改善加工品质[36]。近年来相关研究表明, 籼稻长宽比大有利于高产, 同时显著改善垩白性状, 对加工品质影响较小; 而粳稻长宽比小有利于提高糙米率, 对外观品质影响不大; 南方粳稻增加长宽比将显著降低糙米率和整精米率(图4)[34]。从图4还可以看出, 籼稻增加千粒重将明显降低整精米率, 提高垩白粒率和垩白度, 粳稻提高千粒重对产量和糙米率有正向作用, 对整精米率和垩白性状影响较小。粳稻粒厚和粒宽一样与千粒重正相关, 但是对外观品质影响较小, 还有利于改善加工和食味品质[34]。
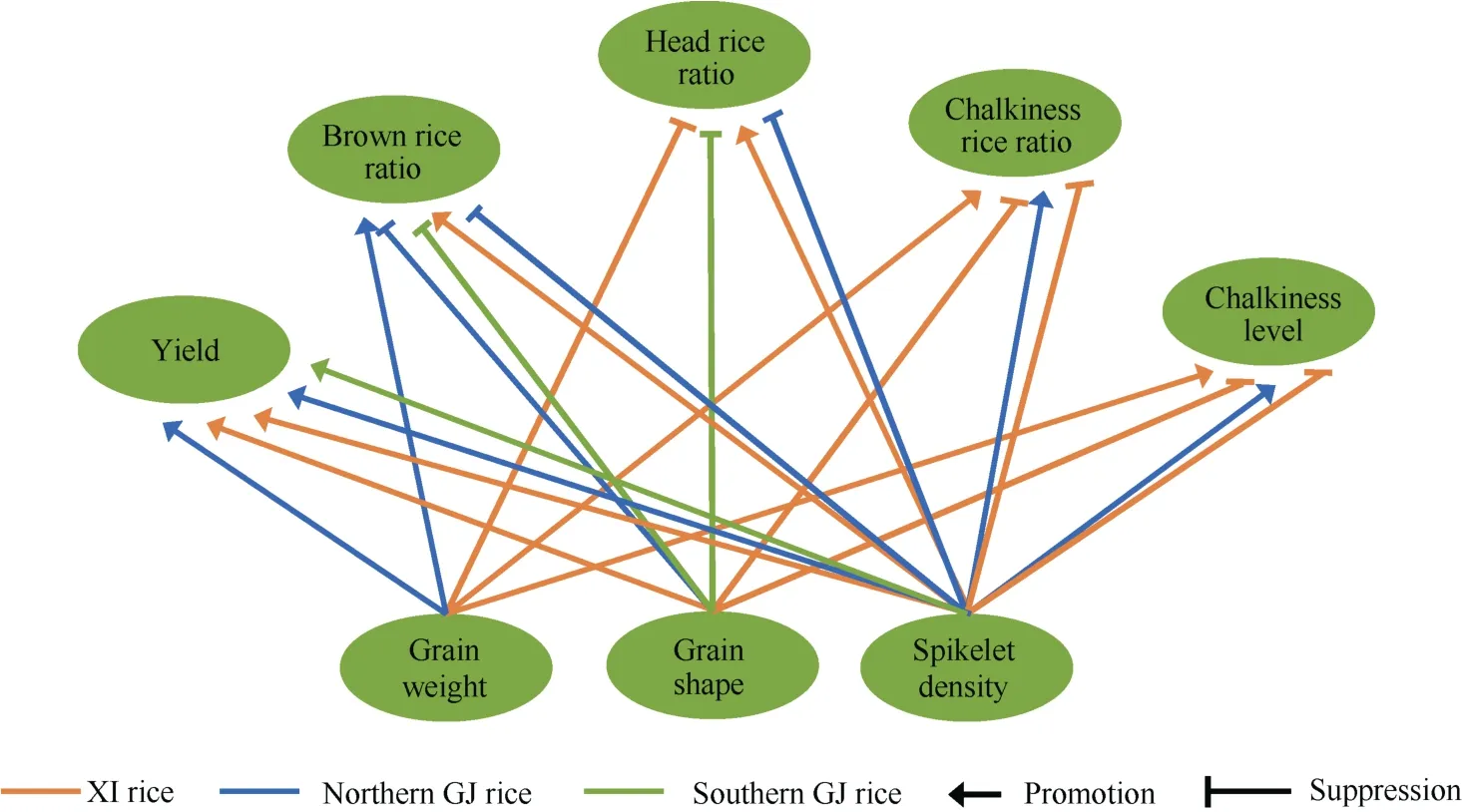
图4 籼粳稻粒型和穗型与产量和品质的关系Fig. 4 Relationship among grain shape, panicle architecture, yield, and quality of XI and GJ rice variety
4 水稻株型遗传基础研究
近年来分子生物学研究成功克隆了大量株型调控主效基因, 水稻株高、分蘖数、分蘖夹角、穗型的基因调控网络逐渐清晰。在20世纪60年代的绿色革命中,sd1基因编码蛋白是赤霉素合成途径中的关键酶, 催化GA53转换为GA20, 作为重要的矮秆基因在生产上大面积应用[37]。独脚金内酯(strigolactones,SLs)作为一类新型的植物激素, 生物合成的关键基因DWARF27(D27)、D17、D10、Os-MAX1s及信号途径相关基因D3、D14和D53在水稻株高和分蘖调控中起关键作用[38]。
分蘖数和分蘖角度直接影响水稻株型,D14可以与D3、D53结合,D53泛素化后被26S蛋白酶体识别并降解, 解除D53对SLs下游响应因子的抑制作用, 同时影响水稻株高和分蘖数[39]。D53与水稻理想株型基因IPA1互作抑制IPA1的转录活性[40]。SLs和油菜素内酯(brassinosteroids, BRs)信号途径都可以通过调节D53和BRs信号途径中转录复合体OsBZR1-RLA1-DLT的稳定性影响水稻Teosinte Banched1(OsTB1)的表达, 进而控制水稻分蘖数[41]。PROG1是匍匐生长、低产的野生稻向直立、高产的栽培稻驯化的重要株型改良位点,Tiller Angle Controlling 1(TAC1)和Tiller Inclined Growth 1(TIG1)是籼粳分化中决定分蘖角度的重要选择位点[12,42-44]。LAZY1(LA1)是从水稻散生材料中克隆的调控分蘖角度的关键基因, 通过分析水稻茎重力反应过程中的动态转录组, 确立了一条以LAZY1(LA1)为核心介导的HEAT STRESS TRANSCRIPTIONAL FACTOR 2D(HSFA2D)-LA1-WUSCHEL RELATED HOMEOBOX 6(WOX6)/11水稻分蘖角度分子途径[45-46], 为阐明水稻分蘖角度形成的分子网络奠定了基础(图5)。
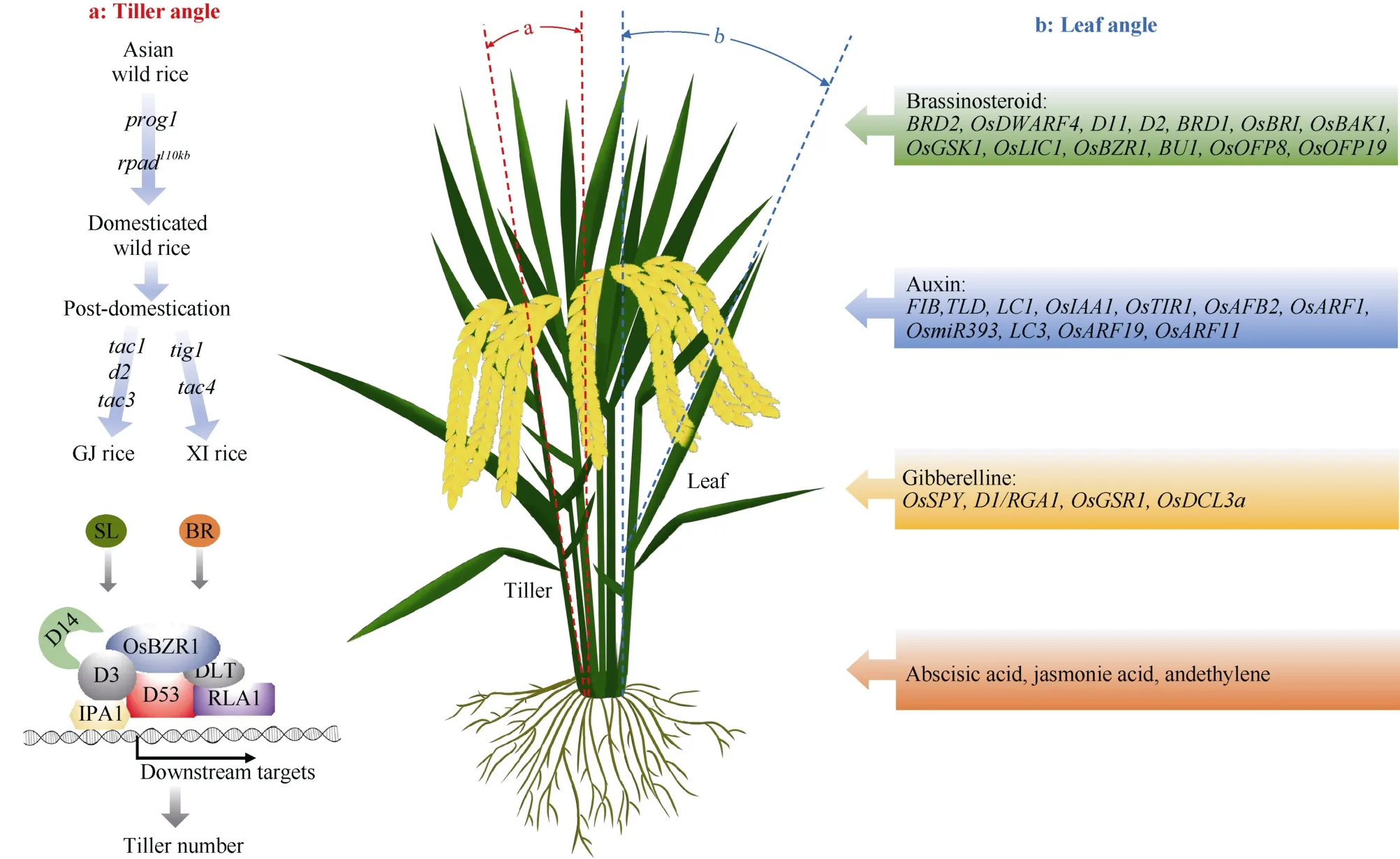
图5 分蘖夹角、分蘖数和叶片夹角的分子网络示意图Fig. 5 Proposed molecular mechanism of tiller angle, tiller number, and leaf angle
叶片夹角是水稻株型构成的一个重要农艺性状,适当直立的叶片适合水稻高密度种植, 叶夹角受油菜素内酯、生长素、赤霉素以及这些激素的互作调控。参与油菜素内酯途径的基因主要有BRD2[47]、OsDWARF4[48]、D11[49]、D2[50]、BRD1[51]、OsBRI[52]、OsBAK1[53]、OsGSK1[54]、OsLIC1[55]、OsBZR1[56]、BU1[57]、OsOFP8[58]、OsOFP19[58]等。生长素(IAA)作为植物中重要的信号分子, 也参与到叶片夹角的调控, 其中主要基因有FIB[59]、TLD[60]、LC1[61]、OsIAA1[62]、OsTIR1[63]、OsAFB2[63]、OsARF1[64]、OsmiR393[63]、LC3[65]、OsARF19[66]和OsARF11[66]。赤霉素(GA)对于植物生长发育具有重要作用, 同样具有调控叶片夹角的功能, 其中已克隆的主要调控基因有OsSPY[67]、D1/RGA1[68]、OsGSR1[69]和OsDCL3a[70]。除了以上3种植物激素外, 脱落酸(abscisic acid,ABA)、茉莉酸(jasmonie acid, JA)和乙烯(ethylene,ETH)也参与调控水稻叶片夹角[71]。ABA通过拮抗BR的生物合成与信号传导影响叶片夹角, 茉莉酸甲酯通过抑制BR的合成和信号传导影响叶片夹角, 而乙烯可能通过与BR的互作影响叶片夹角[72](图5)。
在茎秆和叶片性状相关研究不断深入基础上,水稻穗发育的分子调控机制也取得了重要进展。调控一次枝梗形成的主要基因有Gn1a、LOG、DST、IPA1/WFP、OsSHI1、OsOTUB1、OsSPL17、OsNAC2、FON2/4、FON1等[73-84], 而调控二次枝梗形成的主要基因有FZP、OsMFT1、NAL1/SPIKE、GNP1、TAW1、GN2、OsGRF6、GSN1、GLW7和NOG1[85-96]。水稻直立穗型基因DEP1编码G蛋白Gamma亚基, 是重要的穗部性状调控因子, 其获得功能型等位基因提供分生组织活性, 显著增加每穗粒数[97-100], 同样调控直立穗型的基因还有EP2、EP3和DEP3[101-103], 此外SPED1、APO1、APO2/RFL、SP1、ASP1/OsREL2都在穗型调控中起着重要作用[104-108]。
5 讨论与展望
关于水稻特别是北方粳稻理想株型模式及其主要参数, 本团队已在近期另一篇论文[109]中详细介绍, 下面简要讨论水稻株型相关研究几个问题及发展方向。
籽粒长宽比是籼粳稻重要形态差异, 历来是亚种分类主要指标, 至今仍然是籼稻长宽比基本>2.5而粳稻大多<2.0[34], 但是籼粳亚种粒形差异的生理生态与演化机制还不清晰。游修龄[110]认为南方籼稻区昼夜温差小, 夜间温度高呼吸作用强, 积累的光合产物少, 因而粒重偏低呈细长形, 同理北方粳稻表现为短圆粒形。根据图4可以推测, 目前籼粳稻之所以粒形表现显著差异, 是因为籼稻长宽比大既有利于高产又有利于改善外观品质, 同时对加工品质负面作用较小; 而粳稻长宽比大则显著降低糙米率, 改善外观品质的作用有限; 同处较低纬度的南方粳稻, 增加长宽比将显著降低糙米率和整精米率,既失去了北方粳稻加工品质优势, 又对改良外观品质作用不大, 因此南方粳稻不能像籼稻那样选择细长粒形。近年来南方籼稻千粒重已经明显大于北方粳稻, 籼粳稻千粒重平均值超过26 g, 而北方粳稻只有25 g左右[34]。从另一个角度思考, 水稻亚种分化早期南方籼稻也可以采取减少每穗粒数, 或较小短圆粒形策略应对低纬度不利于光合产物积累的生态环境, 或者说籼稻演化为细长粒形只是随机现象,粒型的籼粳亚种间差异及其与生态环境的关系, 是一个值得深入思考的科学问题。
松尾系统总结了相关领域长期研究成果[111], 认为从物质生产与分配角度分析, 株型改良仍然是进一步提高水稻产量的主要途径: 第一, 叶片直立是迄今改善群体结构的核心, 今后仍然有进一步改良的潜力; 第二, 在保持抗倒伏性的同时降低叶面积密度, 可以通过在保持抗倒伏性前提下增加株高实现, 还可以通过长而直立的剑叶配合短而粗的倒一节间特别是穗颈实现; 第三, 在兼顾穗本身受光的同时, 降低穗在冠层中相对位置以减少对叶片的遮光。尽管穗本身光合能力很低, 但只对穗遮光却导致成熟率降低。籼粳稻地理分布不同, 生态环境有明显差异, 穗型与生态环境的关系及其对产量和品质的影响还需要进一步深入研究。一般日本粳稻剑叶较短, 直立穗型粳稻尤其明显, 对籽粒灌浆的贡献也受到限制, 粳稻直立穗型配合籼稻窄、凹、直、长、厚剑叶的可能性及其效果值得探讨。
根系性状的重要性越来越受到关注, 但是根系性状的复杂性决定了比地上部分更难精准鉴定表型[19]。稻田土壤理化特性比较均匀, 水稻根系分布范围较小, 土芯法等配合根系扫描仪等设备以及合理的田间试验设计, 基本能满足栽培生理研究需要,但是达不到遗传分析等更精准要求[20]。“理想根型”还停留在定性或概念层面, 明显滞后于地上部分茎、叶、穗型研究, 不足以满足遗传生理研究和育种实践需要, 开发高效的田间根系表型鉴定方法,是今后打破水稻根系研究瓶颈, 实现“理想根型”育种实践的关键技术问题[112-113]。
水稻株型看似简单的形态学范畴问题, 实际上是一个复杂的系统工程。迄今除地下部分根系性状研究相对薄弱外, 地上部分茎秆、叶片和穗部性状生理、生态及遗传机制研究都相当深入。但是应该看到, 上述器官之间不但存在外在生理生态相互影响, 还存在内在形态建成遗传调控密切相关。另外,过去主要围绕高产建立的株型模式, 未必适应兼顾高产、优质、高效、绿色的新目标。今后需要充分利用现代生物学特别是生物信息、表型组、基因编辑、分子设计育种、全基因组选择等理论与技术最新成果, 解析各器官之间的外在与内在相互关系,构建和优化对水稻精准株型模式[114]。在此基础上明确分子机制与调控网络并付诸株型育种实践, 实现更高水平全方位的“水稻理想株型”。
猜你喜欢
古今农业(2022年2期)2022-08-15
作物学报(2022年7期)2022-05-11
安徽农学通报(2022年8期)2022-05-06
作物学报(2021年10期)2021-08-05
北方水稻(2021年2期)2021-04-26
中国农业信息(2017年12期)2017-09-03
湖北农业科学(2017年6期)2017-04-26
上海农业学报(2017年4期)2017-04-10
养生保健指南(2016年12期)2017-01-06
农学学报(2016年2期)2016-12-22
