
小麦秸秆及其生物炭对植烟土壤养分、酶活性及细菌群落结构的影响
2023-05-06 13:59李亮生陈帅伟陈享灯宋文静
江苏农业科学 2023年7期
张 燕, 李亮生, 陈帅伟, 李 军, 陈享灯, 宋文静
(1.中国农业科学院烟草研究所/农业农村部烟草生物学与加工重点实验室/中国农业科学院青岛烟草资源与环境野外科学观测试验站,山东青岛 266101; 2.中国农业科学院研究生院,北京 100081;3.福建省烟草公司龙岩市公司,福建龙岩 364000; 4.山东中烟工业有限责任公司,山东济南 250014)
据统计,我国每年秸秆产量9亿t,可加工为副产物的达5.8亿t,其中2.3亿t的副产物可以被利用,剩下的3.5亿t副产物会被丢弃。近年来我国工业化发展迅速,对于秸秆的需求越来越少,如何处理和合理利用这些废弃的秸秆资源已成为当前亟待解决的问题。作为一种重要的生物有机资源,秸秆中含有丰富的有机质及营养元素,包括氮、磷、钾、钙、镁等[1],秸秆腐解后产生的活性有机碳,可以为微生物提供充足的碳源,能够有效增加土壤微生物数量和微生物多样性[2]。秸秆还田后直接增加了土壤有机质含量,使得土壤微生物活性增强,从而提高了土壤酶活性,促进了土壤养分的转化[3]。
生物炭是由有机材料(如秸秆、木材、植物残渣和粪肥)在厌氧的条件下热解(300~1 000 ℃温度范围)生成[4]。生物炭具有孔隙度大、高抗生化分解等特性,常被用作土壤调节剂改善土壤理化性状,提升土壤肥力等[5-7]。在山东烟区,植烟土壤由于长期连作、有机物料不足等不合理的农业管理措施,土壤理化性状较差、微生物活性降低等土壤质量问题已成为制约山东烟区实现可持续发展的瓶颈。科学、合理地利用当地的小麦秸秆资源改善烟田土壤可能是实现山东烟区烟田土壤质量提升的重要途径。目前,关于秸秆及秸秆生物炭对山东烟区烟田氨氧化土壤微生物与土壤养分及相关酶活关联度分析的研究报道较少,本研究通过比较不同秸秆处理方式下植烟土壤的理化性状、养分状况、土壤酶活性以及土壤氨氧化菌的群落组成等的差异,为秸秆资源的合理利用提供理论依据。
1 材料与方法
1.1 试验区概况
试验区位于山东潍坊烟草有限公司诸城分公司洛庄烟站(36°1′N,119°6′E),试验区年日照时数在 2 500 h 以上,年平均气温12 ℃以上,年平均降水量750 mm左右,年平均无霜期230 d左右。土壤基础理化性状如表1所示。
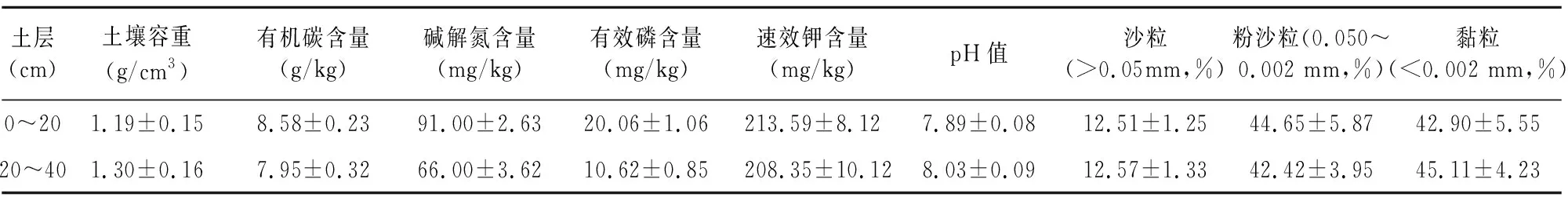
表1 土壤基础理化性状
1.2 试验设计
试验设置3个处理:(1)CK,常规施肥;(2)WS,CK+施用小麦秸秆,还田量6.75 t/hm2;(3)BC,CK+施用小麦秸秆生物炭,还田量2.25 t/hm2。每个处理3次重复,随机区组设计,每个小区面积为 48 m2。小麦秸秆及其生物炭的理化性状详见表2。
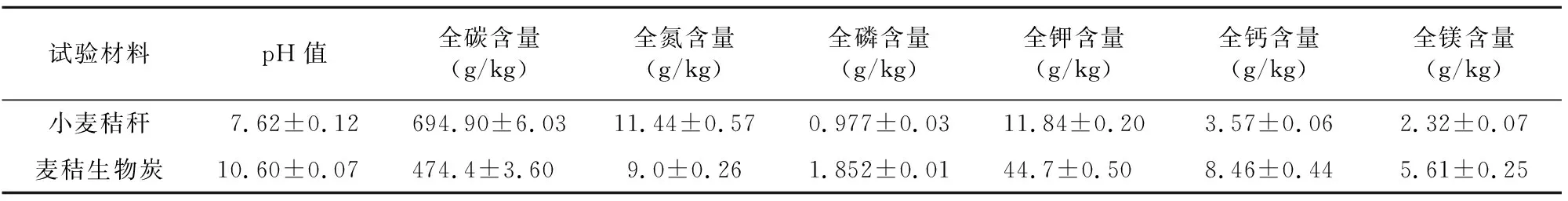
表2 小麦秸秆及其生物炭的理化性状
在2021年3月20日烟田起垄前,撒施小麦秸秆或生物炭,施肥种类及用量情况:复合肥(含N、P2O5、K2O分别为10%、10%、20%)施用量为 600.0 kg/hm2,硫酸钾(含K2O 50%)施用量为 105.0 kg/hm2,磷酸二铵(含N、P2O5分别为16.5%、44.5%)施用量为60.0 kg/hm2。
供试烤烟品种为NC55,移栽时间为2021年5月8日,烤烟的株行距为1.2 m×0.5 m。
1.3 样品采集与测定
1.3.1 土壤的采集及土壤养分的检测 2021年7月20日,于烟叶采烤结束后,按照5S法,用土钻在不同小区于烟垄垄体正中收集耕层土样。土样过 2 mm 筛,用于测定土壤酶活性;其余土壤样品经风干后,测定土壤化学指标;新鲜土样用于检测土壤微生物群落结构及多样性。总有机碳(TOC)、土壤全氮(TN)、碱解氮(AN)、土壤有效磷(AP)、土壤有效钾(AK)含量,土壤pH值均依照土壤农业化学分析方法[8]测定。
1.3.2 土壤酶活性的检测 土壤酶活性[9]测定:土壤蔗糖酶活性选用3,5-二硝基水杨酸比色法测定;土壤脲酶活性选用靛酚比色法测定;过氧化氢酶活性采用紫外分光光度法测定;中性磷酸酶活性采用磷酸苯二钠比色法测定。
1.3.3 土壤微生物提取及高通量测序 采用Fast DNA SPIN Kit for Soil 土壤 DNA 提取试剂盒按步骤提取。利用 Illumina 公司的 MiSeq PE250平台进行测序(上海美吉生物医药科技有限公司)。
1.4 数据分析
应用QIIME软件处理高通量测序结果;利用Mothur软件包以97%为阈值对细菌基因序列划分分类单元,并计算细菌多样性指数和丰富度指数;应用SPSS进行土壤细菌群落组成和相对丰度分析,采用Origin 2021处理数据和制作图表。
2 结果与分析
2.1 秸秆及秸秆生物炭对土壤速效养分和pH值的影响
由图1所示,与CK相比,WS处理显著提高了土壤碱解氮和有效磷含量,增幅分别为78.11%和100.81%,而BC处理养分含量变化较小。各处理间土壤速效钾含量和pH值无显著差异。
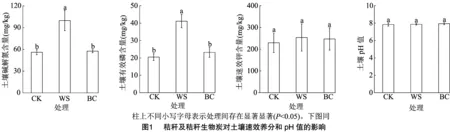
2.2 秸秆及秸秆生物炭对土壤全效养分的影响
如图2所示,WS处理土壤全氮含量较CK显著提高了27.65%。与CK处理相比,WS和BC处理均显著提高了土壤总有机碳含量,增幅分别为20.71%、74.93%。 BC处理较CK处理土壤全钾含量显著提高了21.93%,而WS处理土壤全钾含量变化较小。

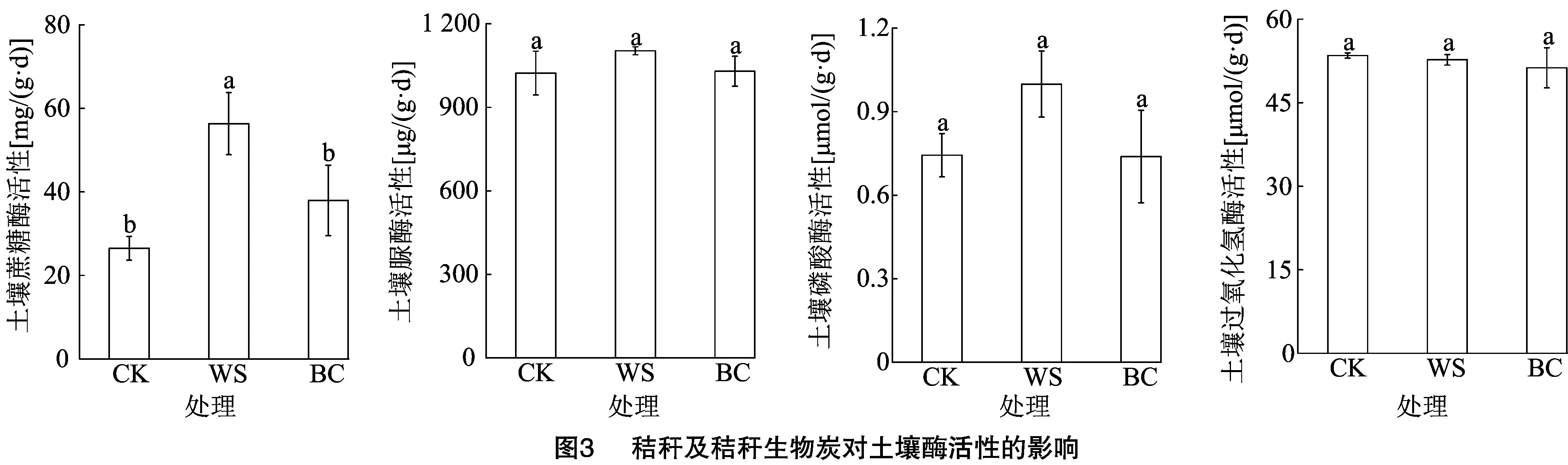
2.3 秸秆及秸秆生物炭对土壤酶活性的影响
如图3所示,与CK相比,WS处理显著提升了蔗糖酶活性,增幅为112.54%,BC处理与CK处理差异不显著。各处理间土壤脲酶、磷酸酶和过氧化氢酶活性无显著差异。
2.4 秸秆及秸秆生物炭对土壤细菌群落的影响
2.4.1 微生物多样性 由表3可知,WS处理中16S基因型细菌Shannon 指数、ACE指数均高于CK处理和BC处理,但差异并不显著,Chao1指数显著高于CK处理和BC处理,与CK相比,增幅为14.19%。各处理nosZ基因型反硝化细菌、AOA和AOB的群落多样性和丰富度指数差异均不显著。
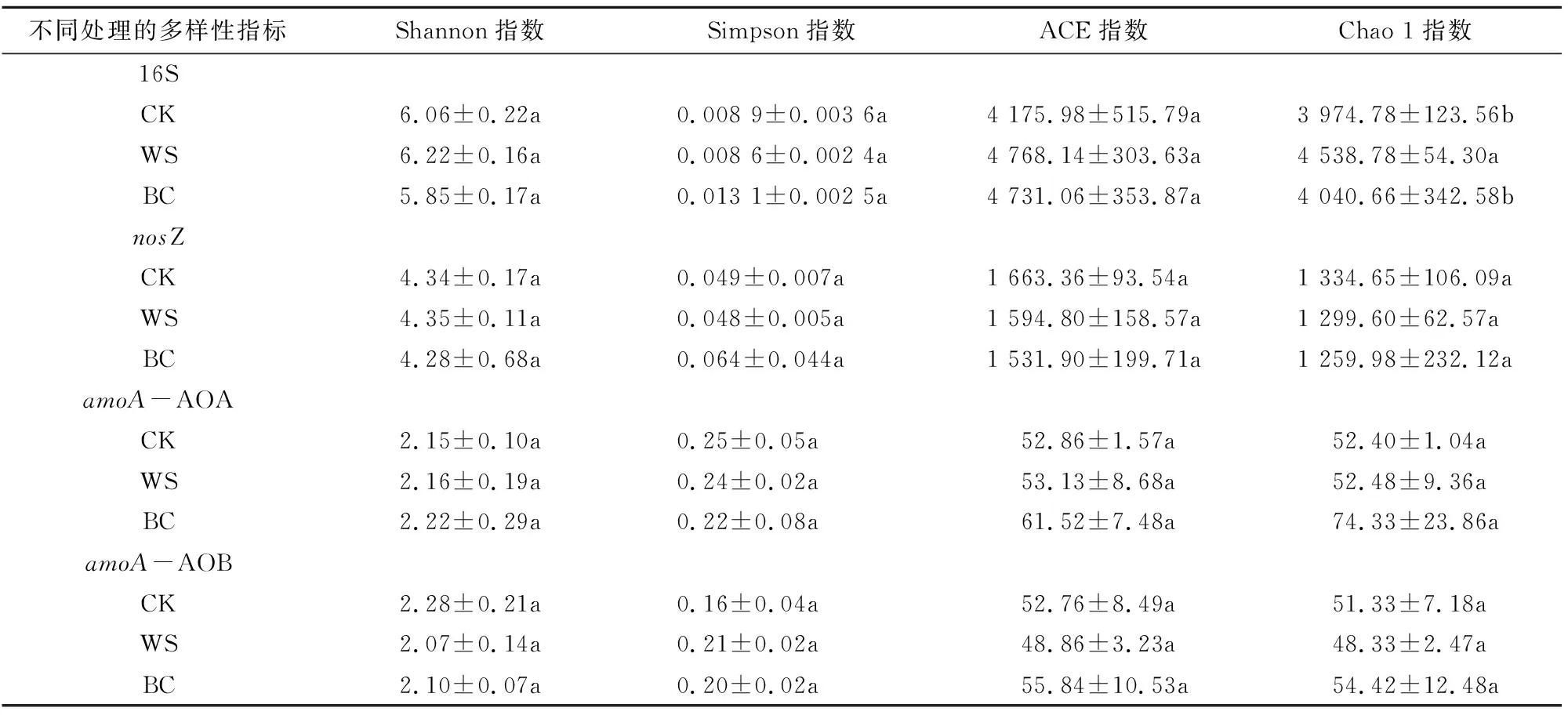
表3 秸秆及秸秆生物炭对土壤微生物和细菌种类丰富度和多样性指标的影响
2.4.2 土壤细菌的群落结构分析 对不同处理优势细菌门水平的分析表明(图4),3种处理中nosZ基因型反硝化细菌群落的优势菌门为变形菌门(Proteobacteria),AOA群落的优势菌门为泉古菌门(Crenarchaeota),16S基因型细菌的优势菌门为放线菌门(Actinobacteria),AOB群落的优势菌门为变形菌门。

对不同处理优势细菌属水平的分析表明,3种处理中nosZ基因型反硝化细菌群落的优势菌属为unclassified_k_norank_d_Bacteria,16S基因型细菌的优势菌属为norank_c_Soil_Crenarchaeotic_Group_SCG,AOA群落的优势菌属为norank_c_environmental_samples_p_Crenarchaeota,AOB群落的优势菌属为norank_f_environmental_samples。秸秆及秸秆生物炭还田后的土壤优势菌群组成与常规施肥一致,但优势细菌群分类组成比例存在差异。
2.4.3 土壤细菌的群落组成分析 由图5可知,第1主成分 PC1 对16S基因型细菌群落组成差异的解释度为27.50%,第2主成分 PC2 对16S基因型细菌群落组成差异的解释度为20.86%,PC1和PC2可累积解释48.36%的变异度。秸秆及秸秆生物炭还田使植烟土壤中16S基因型细菌群落结构发生改变,但对nosZ基因型反硝化细菌(PC1=30.16%,PC2=20.50%)、amoA-AOA(PC1=60.78%,PC2=23.32%)、amoA-AOB(PC1=37.89%,PC2=33.60%)的细菌群落结构改变不显著。

2.5 影响植烟土壤细菌群落的因子分析
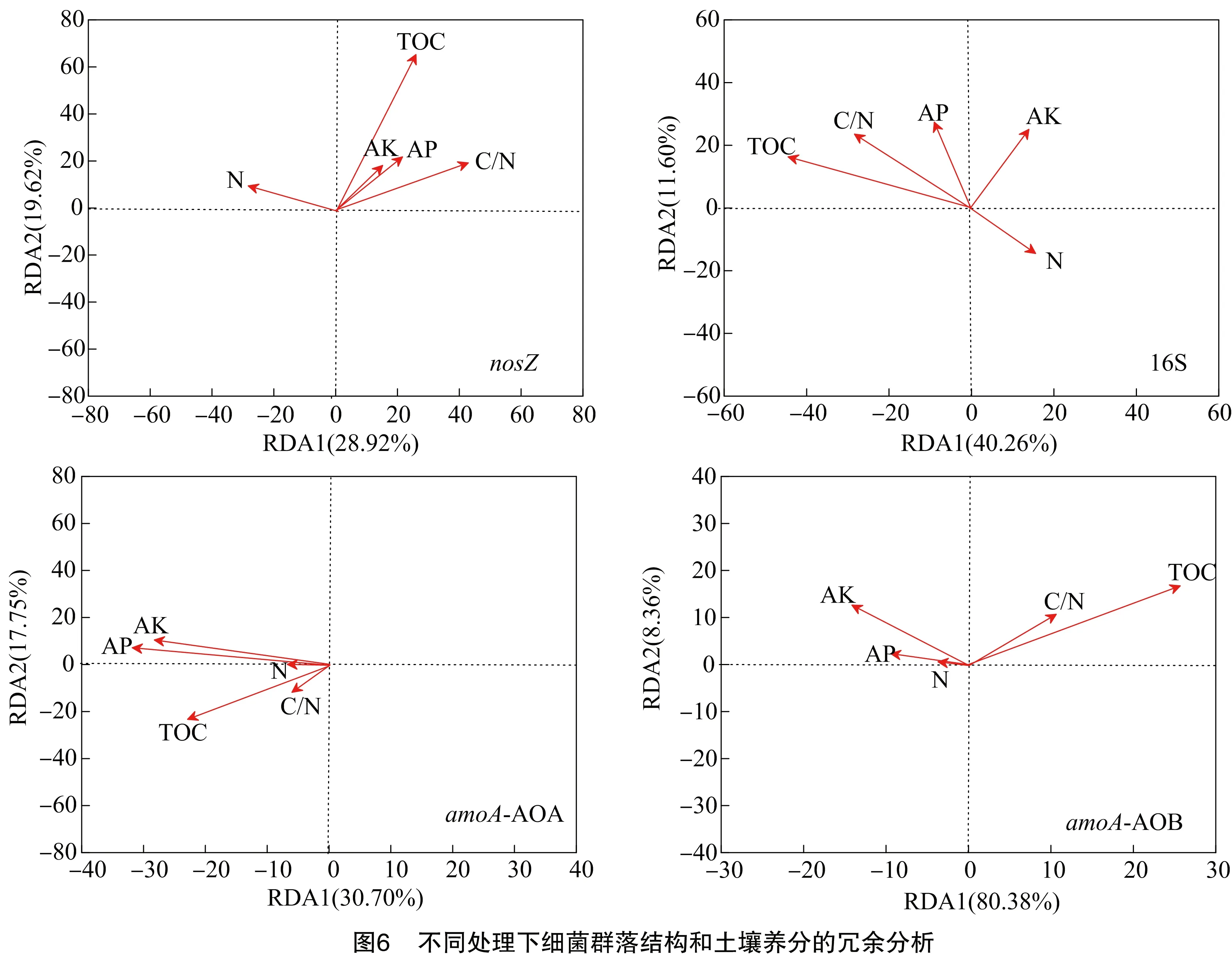
冗余分析结果显示(图6),在不同的处理中,总有机碳含量是引起nosZ基因型反硝化细菌群落变化(r2=0.733,P<0.05)的主控因子,也是引起16S 基因型细菌群落变化(r2=0.627,P<0.05)的主要环境因子,有效磷(r2=0.424,P<0.05)是影响AOB群落变化的主要环境因子。
3 讨论
3.1 秸秆及秸秆生物炭还田对土壤养分的影响
本研究结果表明,秸秆还田显著提升了植烟土壤碱解氮、有效磷含量,这与邹清祺等的研究结果[10-13]一致。秸秆还田对土壤养分含量的提升作用与秸秆矿化分解后向土壤中释放了大量养分元素有关[14]。此外,秸秆腐解形成的腐殖质具有溶磷和解钾的双重作用,也能够通过活化土壤中的磷、钾元素来提升土壤中的速效养分含量[13]。
研究表明,土壤添加生物炭后有利于改善土壤养分状况,增加土壤孔隙度,促进土壤团聚体的形成,构建良好的土壤结构[15]。本研究中,秸秆生物炭处理显著提升了土壤中的总有机碳含量,这与Liu等利用商业木炭残留物制备的生物炭改良玉米田沙性土的研究结果[16]一致。添加生物炭短期内即能提高土壤中的TOC含量的主要原因在于生物炭的芳环结构使其具有高度的稳定性和抗分解特性[17],这使其在土壤中难以为土壤微生物所分解利用。
3.2 秸秆及秸秆生物炭还田对土壤酶活性的影响
土壤酶活性在土壤碳氮代谢和养分循环中发挥重要作用,能够表征土壤养分代谢强度[18]。靳玉婷等在巢湖地区利用秸秆还田改良稻-油轮作农田土壤发现,秸秆还田使水稻季和油菜季土壤磷酸酶和脲酶活性明显提升,且油菜季增幅高于水稻季[19]。程曼等在山西省对黄土旱塬区的研究发现,秸秆直接还田对玉米田脲酶和磷酸酶活性影响较小[20]。在本研究中,小麦秸秆还田提高了土壤蔗糖酶活性,主要是由于:一是秸秆还田丰富了微生物提供可利用碳源的种类和数量[21-22],刺激了微生物代谢;二是秸秆还田改善了土壤理化特性,优化了微生物生活环境,提供了土壤酶载体;三是秸秆还田与化肥配施还能够加速土壤自身有机碳的矿化分解来提升土壤酶活性[23]。
生物炭对土壤酶活性影响因材料来源、制备工艺、土壤类型等的不同而存在较大差异[24-26]。杜倩等在四川凉山的研究发现,玉米生物炭和油菜生物炭明显提升了不同烤烟生育期土壤蔗糖酶、脲酶、蛋白酶活性[26]。张桃香等通过室内试验研究发现,低温条件(300 ℃)较高温条件(600 ℃)下制备的生物炭更有利于南方茶林红壤土脲酶、酸性磷酸酶等酶活性的提高[27]。在本研究中,与秸秆还田对土壤蔗糖酶的提升不同,秸秆生成的生物炭处理对蔗糖酶活性影响较小。这一研究结论与 Mierzwa-Hersztek等的研究结论[28]一致。导致这一现象的根本原因在于秸秆转化为生物炭后有机碳的存在形态发生改变,致使其对土壤微生物活性的影响与秸秆直接还田处理存在较大差异[29]。此外,生物炭表面的官能团对活性底物和土壤酶的吸附也会限制土壤酶活性的提升[30]。
3.3 秸秆及秸秆生物炭还田对细菌群落的影响
土壤微生物多样性被公认为是维持土壤健康的关键因素。本研究发现,秸秆还田使16S基因型细菌群落多样性指数中的Shannon指数、ACE指数和Chao 1指数显著提升。这表明,秸秆还田有助于提升土壤16S基因型细菌群落丰富度和多样性。胡蓉等通过对稻麦轮作系统农田土壤的研究也发现,秸秆还田后,水稻根际细菌群落Chao 1丰富度指数和Shannon多样性指数均明显增加[31]。土壤细菌群落门水平的相对丰度比较发现,变形菌门为nosZ基因型反硝化细菌群落、16S基因型细菌群落和AOA细菌群落共同的优势菌门。
主成分分析发现,秸秆还田和秸秆生物炭处理均改变了16S基因型细菌的群落结构。赵凤艳等的研究也发现,添加不同有机物料后,设施番茄长期连作土壤的微生物群落结构发生了改变[32]。微生物也会反作用于生存的环境,微生物多样性的任何损失也可能会减少多功能性,从而对陆地生态系统提供的服务产生负面影响[33]。在本研究中,秸秆和生物炭处理改变了土壤总有机碳含量,冗余分析进一步指出总有机碳是引起16S 基因型细菌群落发生变化的主要环境因子。
4 结论
秸秆还田提高了植烟土壤总有机碳、碱解氮、有效磷和全氮含量,增加了植烟土壤蔗糖酶活性,提升了16S基因型细菌的丰富度和多样性;添加生物炭显著提高了土壤总有机碳和全钾含量;秸秆还田及其生物炭处理使土壤 16S基因型细菌群落结构发生了改变,总有机碳是引起16S 基因型细菌群落变化的主控环境因子。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29
中国比较医学杂志(2020年4期)2020-05-26
水生生物学报(2019年4期)2019-07-20
生物安全学报(2019年3期)2019-02-15
川北医学院学报(2019年6期)2019-02-10
水生生物学报(2015年1期)2015-02-28
现代检验医学杂志(2015年6期)2015-02-06
实验动物与比较医学(2014年5期)2014-02-28
河南科技(2014年18期)2014-02-27
中国糖料(2013年1期)2013-01-22
