
pH值对罗非鱼肌球蛋白乳化性的影响
2023-05-06 09:17:16张若兰周春霞洪鹏志刘唤明马焕塔黄晓冰
广东海洋大学学报 2023年2期
张若兰,周春霞,2,洪鹏志,2,刘唤明,2,刘 璐,马焕塔,黄晓冰
(1.广东海洋大学食品科技学院/广东省水产品加工与安全重点实验室/广东省海洋食品工程技术研究中心,广东 湛江 524088;2.海洋食品精深加工关键技术省部共建协同创新中心,大连工业大学,辽宁 大连 116034)
肌球蛋白是鱼肉中最重要的功能蛋白之一,约占肌肉总蛋白的30%和肌原纤维蛋白的55%,是影响肉制品质量最重要的蛋白质[1]。肌球蛋白分子由两条重链和四条轻链组成,形如“Y”字[2]。肌球蛋白作为两亲性分子,其分子结构中既含有亲水性基团又含有疏水性基团,能够在O/W 乳液界面形成吸附层,提高空间位阻并防止乳液中油滴聚集[3]。从动力学角度分析,蛋白质在油水界面达到稳定的过程有以下三个步骤:1)蛋白质扩散至油-水界面附近;2)蛋白质吸附到油-水界面,分子结构展开;3)界面上的蛋白质重排形成紧密的界面蛋白膜[4]。物理化学处理导致肌球蛋白构象的微小变化会极大地影响其乳化性[5],而调节溶液pH 值是一种使蛋白质变性和展开的常用方法[6]。不同pH 值可通过改变肌球蛋白的质子化程度和表面电荷来改变肌球蛋白的结构[7]。在极端酸性或碱性条件下,肌球蛋白棒状尾部发生完全解离或部分解离,头部构象发生显著变化,使其呈现“熔球态”[8]。不同的pH 值条件下肌球蛋白构象明显不同,调节蛋白溶液的pH 值可达到改变肌球蛋白功能特性的目的[9]。肌球蛋白经酸碱处理后,在静电斥力的作用下分子结构展开,暴露出蛋白分子内部的疏水基团和残基[7],可更好地吸附在油滴表面,形成致密的界面膜,并提供一定的空间位阻防止油滴的聚集[4]。L-精氨酸辅助超声处理能导致肌球蛋白分子展开,分子柔韧性增强,使其更易吸附在油水界面处,提高乳液的物理稳定性[10]。近年来,乳液在功能性食品中作为递送系统受到广泛关注。鱼肌原纤维蛋白已被证实可作为乳液的天然乳化剂[11],肌球蛋白作为肌原纤维蛋白的主要成分,但是其乳化性质的研究相对较少,且主要集中在鳕(Gadus morhua)[12]、花鲢(Aristichthys nobilis)[10]等。我国作为罗非鱼生产第一大国,2020 年罗非鱼淡水养殖高达165 万t[13],罗非鱼可提供丰富的蛋白资源。因此,本研究以罗非鱼肌球蛋白为研究对象,研究pH 值对肌球蛋白乳化稳定性及界面蛋白组成和结构的影响,以期为为罗非鱼肌球蛋白在食品工业中的开发利用提供依据。
1 材料与方法
1.1 材料与试剂
新鲜罗非鱼,购于广东省湛江市湖光市场,取其背部白肉,分装,于-18 ℃贮藏备用。氯化钠、氯化钾、六水合氯化镁、硫酸铵、乙二胺四乙酸二钠:分析纯,购自广州化学试剂厂。5-腺苷三磷酸二钠盐(ATP)、二硫苏糖醇(DTT)、马来酸,购自广州齐云生物科技有限公司。乙二醇-双-(2-氨基乙醚)四乙酸(EGTA):分析纯,购自上海源叶生物科技有限公司。快速Lowry 法蛋白含量测定试剂盒,购自上海荔达生物科技有限公司。
1.2 仪器与设备
Avanti-J-26 sxp 型高速冷冻离心机,美国贝克曼公司;UV757 紫外分光光度计,上海精科实业有限公司;IKA T-18 均质分散机,东南科仪器有限公司;AH-NANO 超高压纳米均质机,加拿大ATS仪器公司;NanoBrook Omni 激光粒度及Zeta 电位仪,美国布鲁克海文仪器公司;Chirascan V100 圆二色谱仪,荧光应用光物理公司;FV3000 激光扫描共聚焦显微镜,日本奥林巴斯仪器公司。
1.3 实验方法
1.3.1 肌球蛋白的提取 肌球蛋白的提取参考周春霞等[14]的方法。提取后的蛋白保存于4 ℃条件下,并于72 h内使用。肌球蛋白浓度采用Lowry法试剂盒检测,通过SDS-聚丙烯酰胺凝胶电泳(SDSPAGE)和凝胶成像系统计算蛋白质纯度高于86%。
将提取的肌球蛋白用透析液(20 mmol/L Na2HPO4-NaH2PO4,0.6 mol/L NaCl)稀释为10 mg/mL。使用1 mol/L NaOH 或1 mol/L HCl分别调节pH 值至2.0、5.0、7.0和11.0,用于乳化活性检测及乳液制备实验。
1.3.2 乳化活性指数(Emulsifying Activity Index,IEA)和乳化稳定性指数(Emulsion stability Index,IES)的测定 参考Lin等[15]的方法并修改,通过浊度法测定肌球蛋白的乳化活性指数与乳化稳定性指数。分别取不同pH 值的肌球蛋白溶液(6 mL)与2 mL 大豆油于50 mL的塑料量杯中,在12 000 r/min条件下均质1 min,立即取新鲜制备的乳液20 μL 并分散在4 mL 含有质量分数0.1%的SDS 溶液中。迅速读取500 nm 处的光密度,记为D0。样品静置10 min 后,在相同位置再次取样20 μL,重复上述步骤将光密度记为D10。乳化活性指数IEA(单位为m2/g)和乳化稳定性指数IES(单位为min)分别按下列公式计算:
其中,D0为样品初始光密度值;D10为样品放置10 min后的光密度值;N为稀释倍数,N=200;C为蛋白质量浓度,C=5.0 g/mL;φ为油相比例,φ=0.2;Δt=10 min。
1.3.3 肌球蛋白-大豆油乳液的制备 准确量取180 mL上述不同pH 值的肌球蛋白溶液(10 mg/mL),分别加入20 g 大豆油,于10 000 r/min 的条件下均质分散1 min 形成粗乳,然后通过高压均质(60 MPa,循环4 次,4 ℃)制备肌球蛋白-大豆油乳液,分装,于4 ℃贮藏。
1.3.4 乳液粒径和电位的检测 参考陈艾霖等[16]的方法并稍作修改。为避免多重散射,将新鲜制备的乳液使用相应pH 的缓冲溶液(20 mmol/L Na2HPO4-NaH2PO4,0.6 mol/L NaCl)稀释500 倍,通过动态光散射分析乳液平均粒径。乳液和水的反射系数分别设置为1.46 和1.33,在25 ℃的条件下进行测量。样品平衡时间为120 s。
上述同样的样品用相应pH 值蒸馏水稀释200倍,使用Nano Brook Omni激光粒度及Zeta电位仪在25 ℃下测量Zeta-电位值。样品平衡时间为120 s。
1.3.5 乳析指数(Creaming Index,IC)的测定 参考Shi等[10]的方法,将均质后的乳液立即转移到样品瓶(内径2 cm,高7 cm)中,并放置于4 ℃冰箱内。拍照记录乳液制备7 d内分层情况。乳析指数计算如下:
其中,Ht为乳液的总高度,Hs为底部上清液的高度。
1.3.6 乳液微观结构的观察 分别采用光学显微镜和激光共聚焦显微镜观察乳液的微观结构。将乳液稀释10倍,并吸取10 μL置于载玻片上,盖上盖玻片,使用40 倍物镜的光学显微镜观察并拍照[17]。另取10 μL 乳液,同时加入10 μL 尼罗红染液和20 μL尼罗蓝染液,避光反应15 min,随后吸取10 μL 置于载玻片上,盖上盖玻片并排除起泡,置于100倍油镜下使用激光共聚焦显微镜(confocal laser scanning microscopy,CLSM)观察并拍照,其中激发波长为633 nm和488 nm,拍照分辨率为1 024×1 024[3]。
1.3.7 界面吸附蛋白的组成分析 参考Li 等[18]的方法并稍作修改,取不同pH值乳液离心(10 000 r/min,30 min),取上层乳析层,加入4 倍体积相应pH 值磷酸盐缓冲溶液,混匀,调节蛋白浓度至1 mg/mL,进行SDS-PAGE 分析。蛋白溶液分别与上样缓冲液1∶1 混合,煮沸4 min,每孔进样10 μg,凝胶电泳于恒压下(浓缩胶中为80 V,分离胶中为120 V)进行。制备好的凝胶,用染色液在室温下染色1 h,然后用蒸馏水进行脱色,直到出现清晰的条带。
1.3.8 界面蛋白的解吸 参考Li等[4]的方法,使用吐温-20 通过竞争吸附的方法解吸出油滴表面的肌球蛋白,研究界面蛋白的变化。取20 g 乳液,超速离心(20 000 ×g,1 h,4 ℃)分离乳化层。收集乳化层并用去离子水清洗两次。随后将清洗后的乳化层与磷酸盐缓冲溶液(含体积分数2%的吐温20)充分混合,使吐温20 充分置换油滴表面的肌球蛋白,再次离心(20 000×g,1 h,4 ℃),下层清液即为界面吸附蛋白溶液。
1.3.9 二级结构的测定 参考Wu 等[19]的方法,将肌球蛋白乳液以及解吸后的界面蛋白用对应pH 值的缓冲液稀释至0.1 mg/mL,进行圆二色谱(circular dichroism,CD)扫描。波长范围为190~260 nm,上样量0.2 mL,测定分辨率0.5 mm,扫描速度为100 nm/min,灵敏度为0.02°,响应时间为0.25 s,以缓冲溶液做空白,实验值为三次扫描的均值。二级结构含量使用CDNN软件计算。
1.4 数据统计分析
所有实验均重复3 次,结果以平均值± 标准差表示。数据使用Origin 2018 软件作图,数据间的显著性差异采用SPSS Statistics 22.0 软件进行方差分析(ANOVA),并通过Duncan 法检验平均值间的显著性,显著性水平α=0.05。
2 结果与分析
2.1 pH 值对肌球蛋白乳化活性指数和乳化稳定性指数的影响
IEA通过单位质量所能稳定的油水界面面积来表征蛋白质乳化能力,可以反映蛋白质在界面处的吸附能力[4]。IES与蛋白质在一定时间内保持乳液物理稳定性有关,可以反映蛋白质抵抗不稳定性如聚集、絮凝等能力[10]。肌球蛋白在不同pH值条件下的IEA和IES结果(图1)显示,当pH 5.0(肌球蛋白的等电点附近)时,肌球蛋白的IEA和IES值最小,因为此时肌球蛋白表面电荷量少,静电斥力的减小使乳液中的蛋白质絮凝、聚集,导致乳化性降低。等电点附近的低溶解度降低蛋白质在油水界面的吸附能力,从而降低蛋白质的乳化性能[20]。而当pH 值远离等电点后,肌球蛋白表面静电荷含量增加,使蛋白质的溶解性增加,提高肌球蛋白的IEA和IES值,随着酸碱度的增加,在电荷排斥的作用下肌球蛋白分子逐渐展开,氨基酸残基中的疏水基团靠近油相,亲水基团靠近水,蛋白质分子结构展开后,与油滴之间的相互作用增强,提高乳化活性[21]。
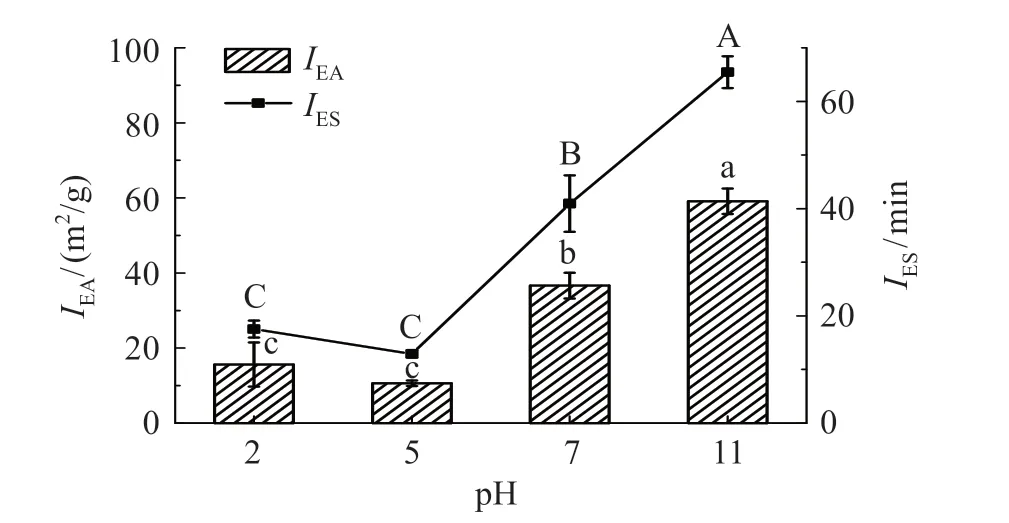
图1 pH值对罗非鱼肌球蛋白IEA和IES的影响Fig.1 Effect of pH value on the IEA and IES of tilapia myosin
2.2 pH值对肌球蛋白乳液平均粒径及电位的影响
乳液平均粒径的大小可以反映蛋白质的乳化能力[22],乳液粒径越小,乳化能力越强。不同pH 值条件下制备的肌球蛋白-大豆油乳液,其平均粒径结果见图2。在不同pH 值条件下,新鲜制备的乳液粒径大小有显著性差异。在pH 5.0 时,肌球蛋白乳液粒径最大,乳液稳定性差;当pH 值远离等电点时,乳液粒径减小(P<0.05)。蛋白乳液的稳定主要由蛋白质提供的空间位阻和液滴之间的静电斥力决定[23]。在等电点附近,肌球蛋白分子表面静电荷接近于零,静电斥力下降使肌球蛋白分子之间发生疏水聚集,与油滴的结合位点减少,蛋白在油滴表面的吸附能力弱,不能形成致密、稳定的界面膜来提供足够的空间位阻使乳液液滴分散。当pH 值远离肌球蛋白等电点时,肌球蛋白开始失去大部分的侧链连接,形成“熔融球体”[8],溶解度增加,可迅速扩散到油-水界面形成界面膜[24],且蛋白吸附能力强,可形成稳定的乳液。
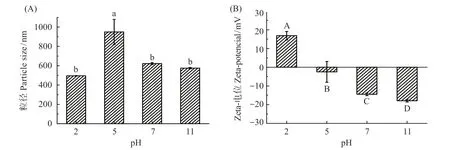
图2 pH值对罗非鱼肌球蛋白-大豆油乳液粒径和Zeta-电位的影响Fig.2 Effect of pH value on particle size and Zeta-potential of tilapia myosin-soybean oil emulsion
Zeta 电位是表征蛋白质表面电荷量的直接指标[11]。在O/W 乳液中,乳液液滴所带的电荷来自于覆盖在油滴表面的蛋白质,电位绝对值越大表示液滴之间静电斥力越大,乳液液滴之间越分散[25]。图2显示,随着pH 值的增加,电位绝对值呈现“V”字型趋势。pH 2.0 时,肌球蛋白乳液带正电荷;当pH 5.0时,乳液电位绝对值最小;而当pH 值增加至7.0 与11.0时,乳液带负电荷。在pH 5.0时,肌球蛋白分子间静电斥力小,蛋白分子间因疏水相互作用而发生聚集,表面疏水性降低,乳液的平均粒径最大。当pH 值处于极端条件时,蛋白质分子发生解离、展开和重排,蛋白质的表面净电荷增加,液滴之间的静电斥力增大,抑制蛋白质分子的聚集,形成的乳液粒径小[26]。在pH 值为2.0、7.0 和11.0 时,肌球蛋白在油-水界面的吸附能力较强,能形成良好的界面膜,并且可提供稳定乳液的静电斥力。
2.3 pH值对肌球蛋白乳液贮藏稳定性的影响
乳液是一种热力学不稳定体系,随着贮藏时间的延长,乳液体系中颗粒运动加快,使油-水界面处吸附的肌球蛋白分子发生重排或脱落,从而改变了其界面结构。图3(a)显示pH 值对肌球蛋白乳液贮藏期时粒径变化情况。4 ℃下贮藏7 d,四种肌球蛋白乳液粒径均显著增大。极端pH 值条件下的肌球蛋白分子结构展开,表面疏水性增加,有利于促进界面处蛋白质吸附至油滴[5],使其稳定的乳液在储藏期粒径变化小,不发生分层。由于布朗运动、重力等因素的影响,乳液中的液滴处于连续运动状态并相互碰撞[10]。较大颗粒的液滴比较小的液滴移动速率更快,液滴间相互碰撞的机率增加,更容易发生聚集现象,因此,pH 5.0条件的乳液平均粒径增长明显。
乳液在贮藏过程中,可以通过记录乳析指数的变化来表征乳液的乳化稳定性[27]。乳析指数越小乳液越稳定。由图3(b)可见,4 ℃下贮藏7 d,只有pH 5.0条件下的肌球蛋白乳液出现分层,且随着贮藏时间的增加,乳析指数增大。由图3(c)也可看出,新鲜制备的肌球蛋白乳液都呈现均匀的白色,在贮藏过程中,pH 5.0 条件下的肌球蛋白乳液发生明显的分层,上层为白色的乳析层,下层为透明的水层。乳液的长期稳定性主要依赖于油水界面吸附蛋白膜的强度与厚度[28]。肌球蛋白在油滴上的吸附是可逆的[3],随着贮藏时间的延长,蛋白质聚集增加会导致无法形成良好的界面膜包裹油滴[11]。当肌球蛋白体系pH 值处于远离等电点时,其稳定的乳液贮藏稳定性良好,在7 d 内不会发生分层现象。这一结果表明,在pH 2.0、7.0 和11.0 时,肌球蛋白在油-水界面的吸附能力强,可形成致密的界面膜防止液滴的聚集,并且在较大的静电斥力作用下,乳液贮藏稳定性得到进一步提高。

图3 不同pH值条件下罗非鱼肌球蛋白-大豆油乳液贮藏过程中粒径、乳析指数及样品的变化Fig.3 Changes in particle size,creaming index and sample of tilapia myosin-soybean oil emulsions during storage under different pH values
2.4 pH值对肌球蛋白乳液微观结构的影响
图4 显示,不同pH 值条件下的乳液液滴直径不同。pH 5.0时乳液发生明显聚集,且聚集体较大,分布广泛,与粒径和Zeta-电位检测结果一致;pH 2.0时乳液有部分聚集;当pH值大于等电点时,乳液的光学显微镜图显示乳液液滴呈现球状且分布均匀,无明显的液滴聚集的情况。在pH 7.0与pH 11.0时,油滴被肌球蛋白完全包裹并均匀分布以形成稳定的乳液,与粒径检测结果一致。
激光共聚焦显微镜(CLSM)可用于肌球蛋白乳液中油滴(红色)和蛋白质(绿色)分布的可视化[5]。从图4 中可发现大小不同的油滴分布在水相中,油滴表面被肌球蛋白包围。在pH 5.0 条件下的乳液液滴聚集明显,此时肌球蛋白间静电斥力小,形成较大的蛋白质聚集体,不能很好包裹油滴使其分散。在pH 7.0、2.0 和11.0 条件下,肌球蛋白分子部分展开,与油滴结合紧密,乳液液滴均匀且尺寸更小,表明蛋白质能够快速吸附在油-水界面形成完整的界面膜,有效防止液滴的聚集从而形成更小的液滴[29],蛋白质的相互作用可以形成多层蛋白质吸附,并在油滴之间产生强大的空间位阻,从而提高乳液的稳定性[30]。
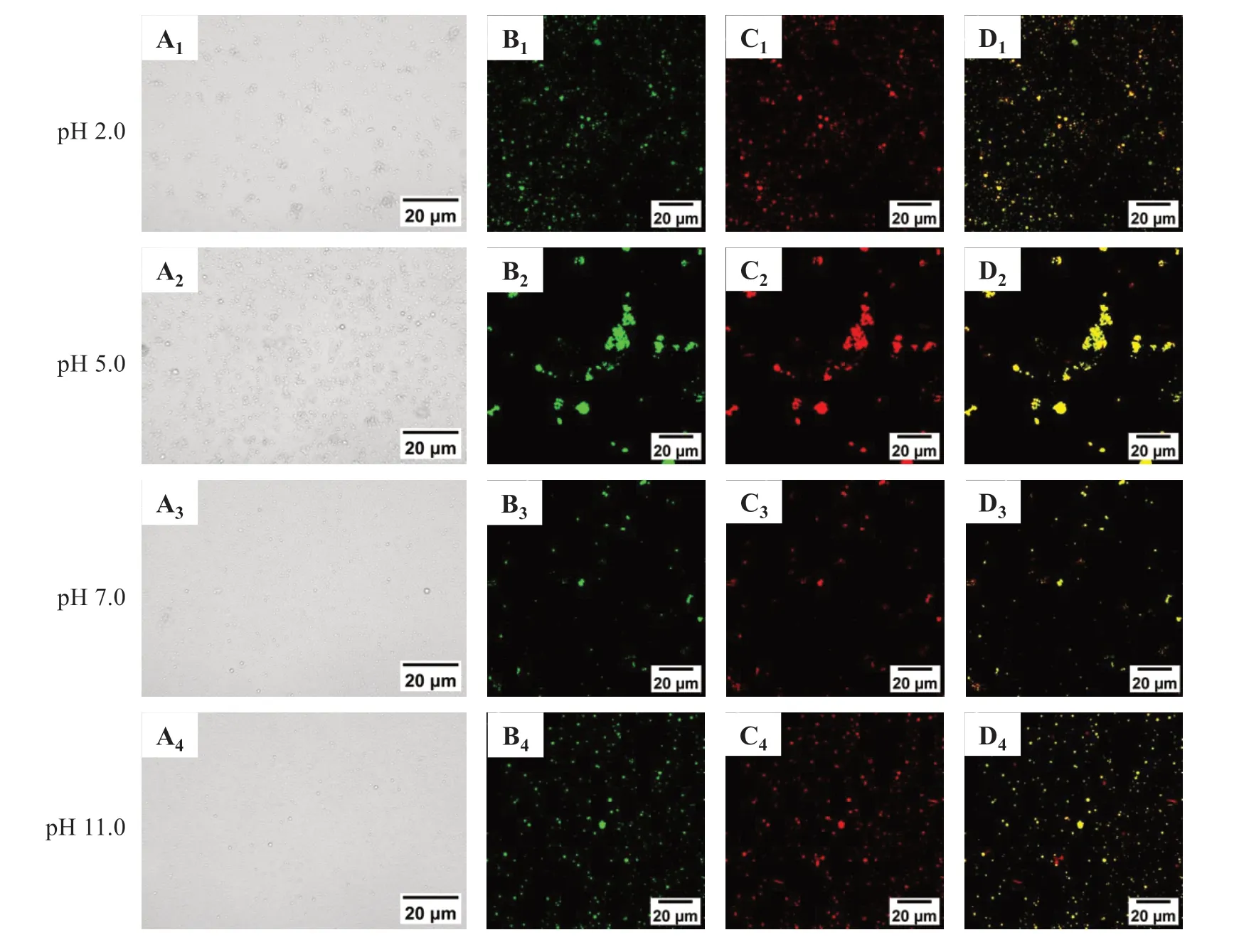
图4 pH值对罗非鱼肌球蛋白-大豆油乳液微观结构的影响Fig.4 Effect of pH value on the microstructure of tilapia myosin-soybean oil emulsion
2.5 pH值对肌球蛋白乳液界面蛋白组成的影响
通过SDS-PAGE 分析吸附在油-水界面的蛋白质组成,结果如图5 所示。未经处理的肌球蛋白在220 ku 附近显示明显的肌球蛋白重链(myosin heavy chain,MHC)条带,在30 ku 附近有一条轻微的原肌球蛋白条带,在15~22 ku 附近显示肌球蛋白轻链(myosin light chain,LHC)条带。乳液界面吸附肌球蛋白主要包括MHC 和LHC。但与未经处理的肌球蛋白相比,不同pH 值条件下肌球蛋白乳液体系中界面蛋白的MHC 和LHC 条带颜色和宽度均有所变化。蛋白质发生降解主要表现为蛋白质条带的模糊、弱化和扩展[17],这表明pH 值的变化及高压均质乳化等可能对蛋白质的交联降解有影响。在pH 11.0 时,乳液的稳定性较好,经离心后没有分离出明显的乳析层,导致此时的电泳条带颜色较浅;而在蛋白质等电点时,肌球蛋白分子展开受到抑制,不利于界面蛋白膜的形成[31],此时蛋白质分子间静电斥力最小,蛋白质包裹的油滴相互靠近并发生聚集,导致肌球蛋白发生聚集和沉淀,电泳条带变窄,与粒径检测结果一致。油滴表面的界面蛋白膜由肌球蛋白构成,因此,肌球蛋白发生交联会使此条件下的蛋白乳液稳定性下降,可能发生分层等情况。
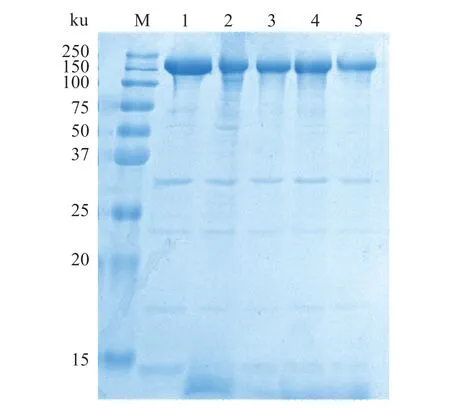
图5 pH值对罗非鱼肌球蛋白-大豆油乳液界面蛋白组成的影响Fig.5 Effect of pH value on the interfacial protein composition of tilapia myosin-soybean oil emulsion
2.6 pH值对乳液及界面肌球蛋白二级结构的影响
为进一步了解乳液及界面肌球蛋白的二级结构及其与乳液稳定性的关系,使用CD 光谱分析不同pH 值肌球蛋白乳液中界面蛋白的二级结构。肌球蛋白棒状尾部95%是由α-螺旋结构构成,肌球蛋白α-螺旋含量的变化可反映杆状尾部的结构[32]。由图6 可见,未经处理的肌球蛋白CD 光谱在208 nm和222 nm 处有两个明显的负峰,这是α-螺旋的特征峰[33]。经计算可得,未处理的罗非鱼肌球蛋白二级结构包括67.93% 的α-螺旋、3.53% 的β-折叠、12.60%的β-转角和13.63%的无规则卷曲。鱼的种类不同,其肌球蛋白含量也有所变化。大眼金枪鱼(Thunnus obesus)的肌球蛋白含有65%的α-螺旋结构,而明太鱼(Gadus chalcogrammus)肌球蛋白α-螺旋结构含量仅为38%[32]。Liu 等[34]研究发现,pH 值对肌球蛋白α-螺旋结构含量的影响主要是通过改变蛋白质分子带电氨基酸和表面的α-羟基及α-氨基末端的质子化程度来实现。CD 光谱显示乳液稳定性与α-螺旋含量呈正相关[19]。α-螺旋结构在油滴表面的暴露有利于蛋白质和油滴之间形成较强的相互作用,较高含量的β-折叠表明相邻蛋白质分子之间有更多的交联,形成蛋白质界面层,为乳液提供一定的空间位阻,体系的稳定性增强,但是当β-折叠结构含量过高时,最外层的蛋白质可能会表现出一些聚集,从而导致乳液液滴的不稳定[4]。
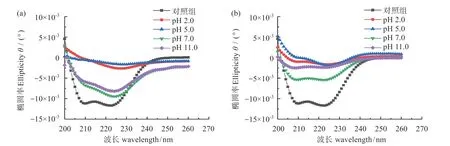
图6 不同pH值条件下肌球蛋白在乳液(a)与界面吸附(b)状态下的CD光谱Fig.6 CD spectra of myosin in emulsion(a)and interface adsorption(b)states at different pH values
与未经处理的罗非鱼肌球蛋白(对照组)相比,乳液状态和界面吸附状态下肌球蛋白的二级结构含量变化趋势一致,即α-螺旋结构含量显著下降,表明高压均质制备乳液过程中的强烈剪切、撞击和空穴作用等可能使肌球蛋白的α-螺旋结构部分被破坏。表1显示,在pH 7.0时,乳液肌球蛋白的α-螺旋质量分数为(54.90±2.56)%,远离中性pH值条件时,界面肌球蛋白α-螺旋含量均显著下降。由于肌球蛋白尾部对酸性pH 环境敏感,因此,在pH 2.0 和pH 5.0时,肌球蛋白尾部螺旋结构解离,易形成β-折叠结构[35],较高的β-折叠结构含量可能使蛋白质分子在界面发生聚集,影响乳液的稳定性,且pH 5.0时,肌球蛋白分子表面净电荷接近于零,分子间静电斥力的下降使肌球蛋白发生疏水聚集,故该条件下肌球蛋白的乳化活性与乳化稳定性最差。与酸性处理相比,肌球蛋白的二级结构在碱性pH 条件下的变化较小[32]。pH 11.0 时,肌球蛋白分子结构展开,表面疏水性增强,疏水基团的暴露有利于蛋白质分子吸附在油滴表面,提高肌球蛋白的乳化性,且此时β-折叠结构含量的适当增加有利于蛋白质间的相互作用,能提供一定的空间位阻稳定乳液,提高乳液贮藏稳定性。
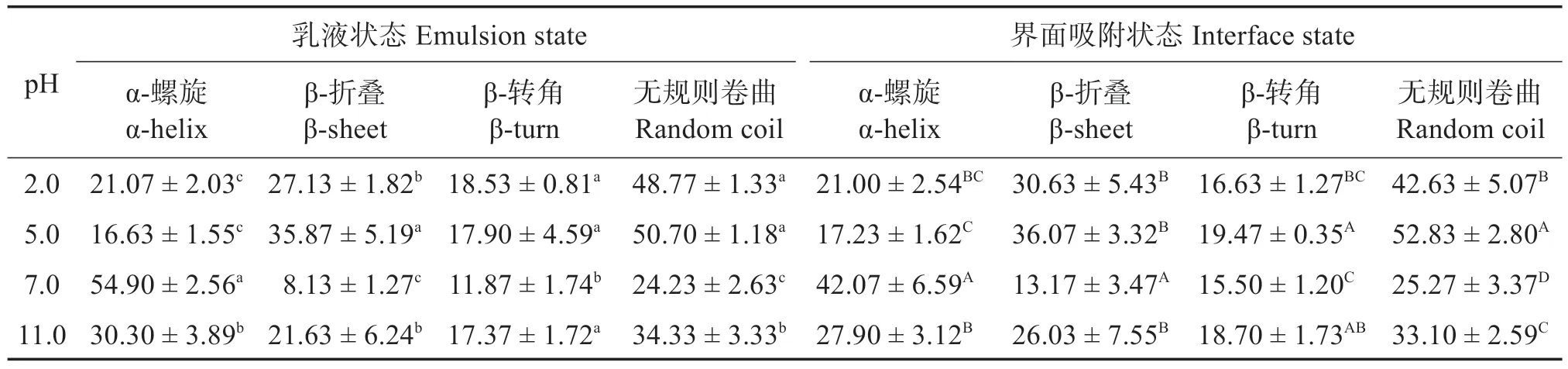
表1 pH值对肌球蛋白在乳液与界面吸附状态下的二级结构质量分数的影响Table 1 Effect of pH value on the secondary structure content of myosin in emulsion and interface adsorpotion states %
3 结论
pH 值的变化会显著影响肌球蛋白的乳化活性及乳液的稳定性。当pH 值为7.0 时,肌球蛋白经高压均质处理后二级结构变化较小,蛋白质分子完全包裹油滴并均匀分布,乳液贮藏7 d 不分层。当pH值接近肌球蛋白等电点(pH 5.0)时,分子间静电斥力小,蛋白质分子在油-水界面发生聚集,未形成完整的界面膜,乳液液滴聚集,乳液稳定性差。而在极端pH 值(2.0 和11.0)条件下,肌球蛋白分子结构部分展开,蛋白质分子和油滴之间的相互作用增强,在油滴表面形成稳定的界面蛋白膜,乳液稳定性提高。且当pH 值为11.0时,β-折叠结构含量的适当增加更有利于蛋白质分子间的相互作用,提供一定的空间位阻使乳液更加稳定。因此,pH值的调节能改变肌球蛋白的分子结构,进而改善其乳化性。
猜你喜欢
小主人报(2022年18期)2022-11-17 02:19:56
实验技术与管理(2020年7期)2020-09-29 08:44:20
高校化学工程学报(2020年2期)2020-06-10 08:53:26
原子能科学技术(2020年1期)2020-03-30 08:39:02
中国石油大学学报(自然科学版)(2015年2期)2015-11-10 06:07:47
燃气涡轮试验与研究(2015年6期)2015-10-28 05:50:06
生命科学研究(2015年2期)2015-04-29 23:09:30
郑州大学学报(医学版)(2015年2期)2015-02-27 14:50:43
郑州大学学报(医学版)(2015年2期)2015-02-27 14:50:43
心血管病学进展(2015年4期)2015-02-22 07:56:44
