
不同禾本科作物中ZmWRKY79同源基因的鉴定与分析
2023-05-05 06:33卢源达杜云龙刘学思赵建屛吴洪钦李玉琢朱文强邢世军程在全钟巧芳
西南农业学报 2023年3期
卢源达,陈 玲,杜云龙,刘学思,柯 学,赵建屛,吴洪钦,刘 鑫,李玉琢,朱文强,邢世军,程在全,钟巧芳
(1.云南农业大学植物保护学院, 昆明 650224;2.云南省农业科学院生物技术与种质资源研究所/云南省农业生物技术重点实验室/农业部西南作物基因资源与种质创制重点实验室, 昆明 650205)
【研究意义】禾本科作物在中国的主要粮食作物中占据重要地位,保障其增产、稳产尤为重要[1]。近年来,人口数量逐年增加,极端天气频繁出现,作物遗传多样性连续下降等一系列问题给禾本科作物的增产稳产带来了极大挑战[2]。其中,非生物胁迫是制约产量的关键因素,例如:干旱、盐碱、低温、高温、洪涝和氧化胁迫等都会严重降低其产量。由此说明,研究发掘并利用禾本科作物中的抗逆相关基因对提高作物产量意义重大。【前人研究进展】植物转录因子(TFs)在植物遭受逆境胁迫时,调动机体复杂的信号网络来介导植物的抗逆性,在这个大家族中,WRKY转录因子发挥着极为重要的作用[3]。WRKY转录因子的称谓来源于其同源基因均含有高度保守的七肽结构基序WRKYGQK[4]。WRKY转录因子分为3类:I类(2个WRKYGQK七肽序列结构和1个锌指结构Cx4-5Cx22-23HxH)、II类(1个WRKYGQK七肽序列结构和1个锌指结构Cx4-5Cx22-23HxH)、III类(1个WRKYGQK七肽序列结构和1个锌指结构Cx7Cx23HxC)。自20世纪90年代从甜薯中克隆出第1个WRKY基因以来,在水稻、高粱、小麦、青稞、玉米、谷子等禾本科作物中陆续发现了大量的WRKY基因[5]。目前,从玉米中发现了123个WRKY基因(I类:28个,II类:65个,III类:30个);水稻中发现了94个WRKY基因(I类:19个,II类:48个,III类:27个);高粱中发现了97个WRKY基因(I类:11个,II类:70个,III类:16个)[4]。WRKY转录因子的保守基序与其靶基因的顺势调控元件W-box(T)(T)TGAC(C/T)结合从而介导植物生长及抗逆过程[5-6]。尽管在玉米中已经发现了123个WRKY基因,但大部分WRKY转录因子调控玉米抗逆反应的机制有待研究。近年WRKY转录因子的研究热点主要集中在耐盐胁迫、干旱胁迫、植物抗病相关的研究,如:ZmWRKY83参与植物抗病反应[7],ZmWRKY104[8-9]和ZmWRKY86正向调控盐胁迫反应[10],ZmWRKY79正向调节玉米植物保卫素生物合成基因表达,在干旱胁迫下通过调控ABA生物合成的功能,还参与其它非生物胁迫反应[11-12]。这4个转录因子中ZmWRKY79具有较强的发掘潜力。【本研究切入点】不同禾本科作物之间存在同源基因,这些基因往往具有相似的功能。对于ZmWRKY79的同源基因鉴定与分析目前尚无报道。【拟解决的关键问题】本研究将ZmWRKY79基因作为研究对象,利用生物信息学从不同的禾本科作物中鉴定ZmWRKY79的同源基因,同时也对这些基因进行序列比对、结构分析、进化分析、启动子调控元件预测、逆境胁迫下的转录分析,为发掘出新的功能基因奠定理论基础。
1 材料与方法
1.1 同源基因鉴定和染色体定位
利用NCBI(National Center for Biotechnology Information,https://www.ncbi.nlm.nih.gov/)中的Nucleotide数据库获取ZmWRKY79基因编码区域的核苷酸序列,利用blast工具对其进行检索进而获得同源基因的编码区域序列,进一步利用NCBI中Gene数据库获取同源基因染色体定位信息和启动子序列。根据染色体定位信息,利用MG2C(http://mg2c.iask.in/)进行染色体图谱绘制[13]。
1.2 系统进化树构建和共线性分析
利用BioXM2.6获取基因家族结构信息,并利用WPS绘制基因结构图。利用SMART(http://smart.embl-heidelberg.de/)对蛋白保守结构域进行预测[14]。利用DNAman对保守结构域的氨基酸序列进行多序列比对。利用Clustal(https://www.ebi.ac.uk)对氨基酸序列进行比对获得多重比对程序,再利用MEGA 7.0软件建立NJ(邻接法)系统发生树[15]。
1.3 蛋白保守基序(Motif)和启动子预测
利用在线软件MEME 5.0(http://meme-suite.org)对ZmWRKY79同源基因的蛋白序列进行保守基序分析,参数值设定为motif大小在6~60 aa,输出6个motif。将ZmWRKY79同源基因的启动子序列上传至在线软件PlantCARE(http://bioinformatics.psb.ugent.be)进行顺式作用调控元件预测[16],并用TBtools绘制相关热图[17]。
1.4 亚细胞定位转录组数据获取
利用Crop-PAL(https://crop-pal.org/)进行亚细胞定位预测[18]。将玉米中ZmWRKY79基因同源基因的核苷酸序列上传至qTeller数据库(https://qteller.maizegdb.org/index_B73v4.php),进而获得供体材料B73苗期在接受非生物胁迫(冷、热、干、盐、紫外线)处理下所对应基因的转录组数据[19],并利用GraphPad Prism 8.0软件绘制相应的柱状图。
2 结果与分析
2.1 ZmWRKY79同源基因鉴定
通过在线网站NCBI获取ZmWRKY79基因的CDS序列(ID:LOC100193434)和氨基酸序列(ID:NP_001132026.2)。以ZmWRKY79基因的蛋白序列为模板,在NCBI的蛋白数据库中进行blastp检索,结果显示有50条蛋白序列与NP_001132026.2相似度超过49%。对这50条序列进行蛋白结构域(WRKY结构域和锌指蛋白结构域)预测,结果显示,50条序列中,只有9条含有完整的WRKY结构域(WRKYGQK)和锌指蛋白结构域(CX7CX23HC),将这9条序列暂定为ZmWRKY79的同源基因(表1),经比较分析,这些成员为第III类WRKY转录因子,这些成员依次是玉米(Zeamays)B73基因组中的NP_001351600.1(ZmWRKY53)和NP_00115 1583.1(ZmWRKY74)、高粱(Sorghumbicolor)BTx623基因组中的XP_002460069.1(SbWRKY54)和XP_002460068.2(SbWRKY17)、谷子(Setariaitalica)Yugu1基因组中的XP_004956613.1(SiWRKY70)、水稻(Oryzasativa)Nipponbare基因组中的XP_015651250.1(OsWRKY30)、普通小麦(Triticumaestivum)Chinese Spring基因组中的XP_044388521.1(TaWRKY19-like)、野生二粒小麦(Triticumdicoccoides)Atlit2015基因组中的XP_037434168.1(TdWRKY41)、青稞(Hordeumvulgaresubsp)vulgare基因组中的XP_044985695.1(HvWRKY19-like)。
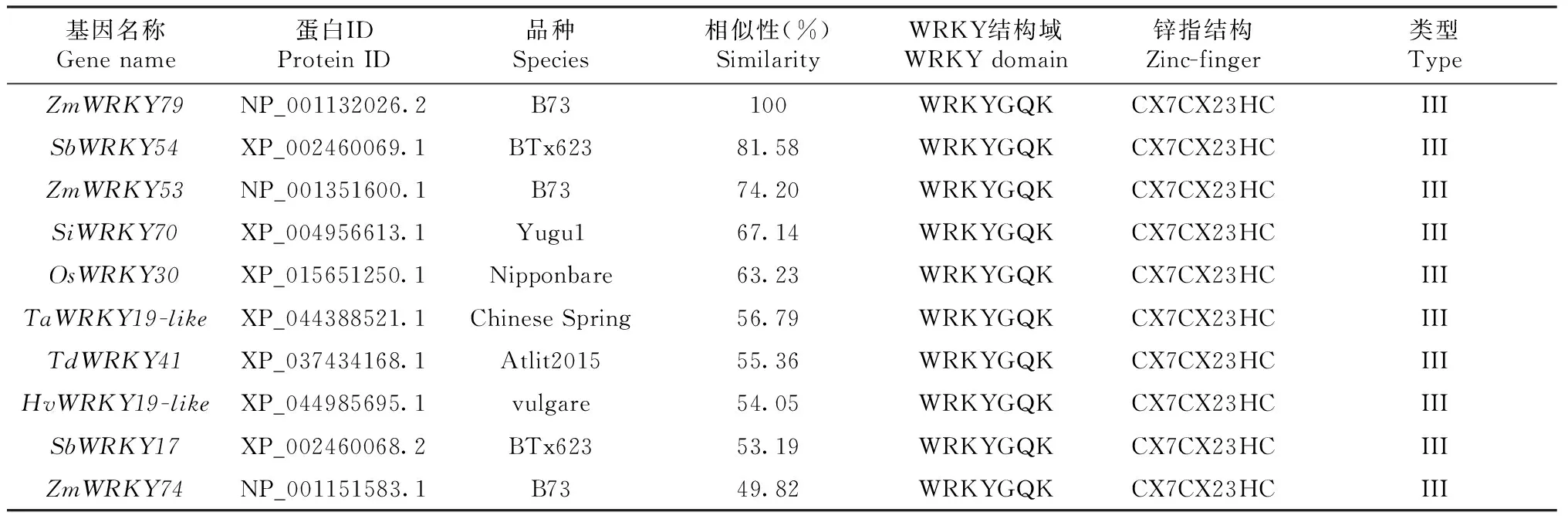
表1 ZmWRKY79同源基因序列信息Table 1 The sequence information of ZmWRKY79 homologous genes
2.2 ZmWRKY79同源基因在染色体上的分布
将10个成员锚定在染色体上(图1),发现ZmWRKY53、ZmWRKY74、ZmWRKY79依次位于玉米(2n=20)2号染色体物理位置180.59 Mb处、7号染色体物理位置96.76和96.86 Mb处。SbWRKY17和SbWRKY54依次位于高粱(2n=20)2号染色体物理位置55.13和55.19 Mb处。SiWRKY70位于谷子(2n=18)II号染色体物理位置27.23 Mb处。TdWRKY41位于野生二粒小麦(2n=4x=28)5A号染色体物理位置332.19 Mb处。TaWRKY19-like位于普通小麦(6n=42)5B号染色体物理位置337.77 Mb处。HvWRKY19-like位于青稞(2n=14)5H号染色体物理位置394.40 Mb处。OsWRKY30位于水稻(2n=24)9号染色体物理位置10.13 Mb处。

图1 ZmWRKY79同源基因的染色体定位Fig.1 Chromosomal localization of the ZmWRKY79 homologous genes
2.3 ZmWRKY79同源基因进化分析
2.3.1ZmWRKY79同源基因结构分析 对10个成员的基因结构进行可视化分析,发现ZmWRKY79的同源基因核苷酸大小在1206~1792 bp,其中ZmWRKY79基因最短为1206 bp,SbWRKY17基因最长为1792 bp。由图2可知,基因大小与外显子数量无关,10个成员中,只有来自小米的SiWRKY70成员含有2个外显子和1个内含子,其余成员都含有3个外显子和2个内含子,这些成员中,除SbWRKY54第1外显子比较长之外,其余成员都是第3个外显子最长,长度介于498~648 bp,第2外显子最短介于120~135 bp,第1外显子次之介于251~422 bp。由此说明,ZmWRKY79同源基因的基因结构相似度很高。
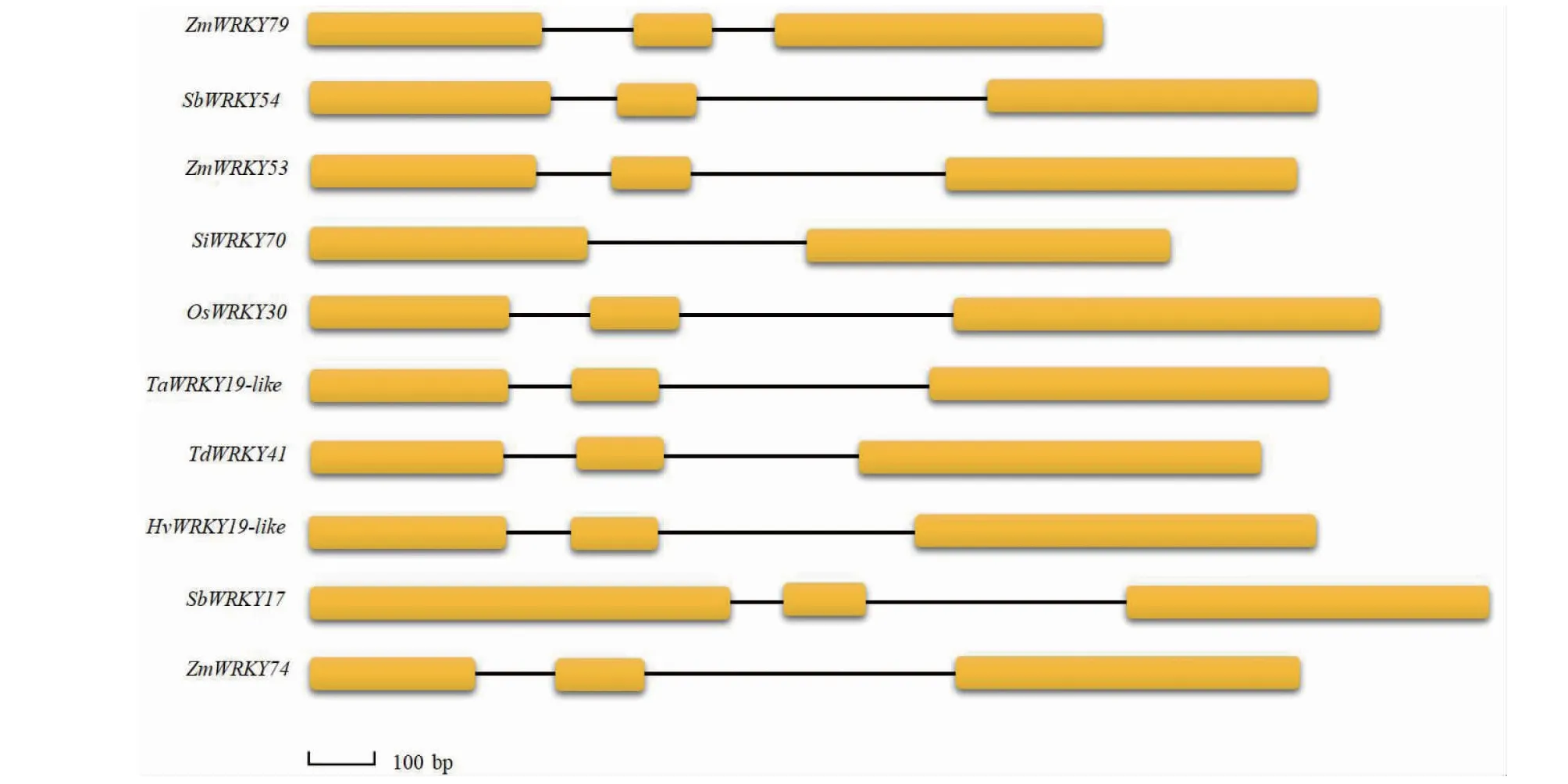
图2 ZmWRKY79基因的同源基因基因结构Fig.2 Gene structure of ZmWRKY79 homologous genes
2.3.2ZmWRKY79同源基因功能结构域氨基酸序列分析 对ZmWRKY79同源基因的氨基酸序列进行多重比对(图3),发现WRKY79蛋白比较保守,尤其七肽基序保守性最强,10个成员在该区域均未发生任何变异,虽ZmWRKY79蛋白21个变异位点就有14个位于锌指蛋白结构域中,但这些变异并不影响ZmWRKY79蛋白类型。由此说明ZmWRKY79同源WRKY功能结构域非常保守。

a代表WRKY结构域;b代表锌指蛋白结构域。a represents the WRKY domain; b represents the zinc finger protein domain.图3 ZmWRKY79同源基因保守结构域的氨基酸序列比对Fig.3 Amino acid sequence comparison of the conserved structural domains of ZmWRKY79 homologous genes
2.3.3ZmWRKY79同源基因的蛋白保守性分析 利用保守基序预测工具MEME对供试的10个ZmWRKY79同源基因的蛋白进行保守基序预测(图4),结果预测到6个保守基序,6个保守基序大小介于15~50个氨基酸,其中Motif1(WRKYGQKEILGAKHPRGYYRCTHRHSQGCAATKQVQRTDEDPALFDVIYH)在WRKY功能结构域内,Motif5(HNPDAQSHLQSLSAGLTVKTEGLPAAPAQQQ)和Motif6(ENWGVSPATSDSNHV)在锌指结构域内。与此同时,供试的10条蛋白序列均拥有这6个保守基序,除SbWRKY54成员的6个保守基序位置变化比较大外,其余成员的6个保守基序排序和位置都基本一致。
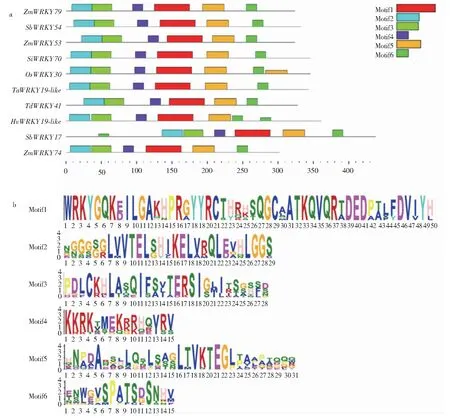
a:ZmWRKY79同源基因的保守基序; b:ZmWRKY79同源基因的蛋白序列标识。a:Conserved motif of ZmWRKY79 homologous genes; b:Protein sequence identity of ZmWRKY79 homologous gene.图4 ZmWRKY79同源基因的保守基序分布和蛋白序列标识Fig.4 Conserved motif distribution and protein sequence identity of the ZmWRKY79 homologous genes
2.3.4ZmWRKY79同源基因的系统发育 利用MAGA软件对供试的10条氨基酸序列构建系统发生树(图5)。系统发生树结果显示,供试的ZmWRKY79蛋白同源序列可被分为2大类,其中来自玉米、高粱和小米的6个成员ZmWRKY53、SiWRKY70、ZmWRKY79、SbWRKY54、SbWRKY17、ZmWRKY74归属为I类,6个成员中,ZmWRKY79、SbWRKY54的亲缘关系最近,二者遗传距离为0.143(表2);来自水稻、小麦和青稞的4个成员OsWRKY30、HvWRKY19-like、TaWRKY19-like和TdWRKY41归属为II类,4个成员中,HvWRKY19-like、TaWRKY19-like的亲缘关系最近,二者遗传距离为0.083(表2)。
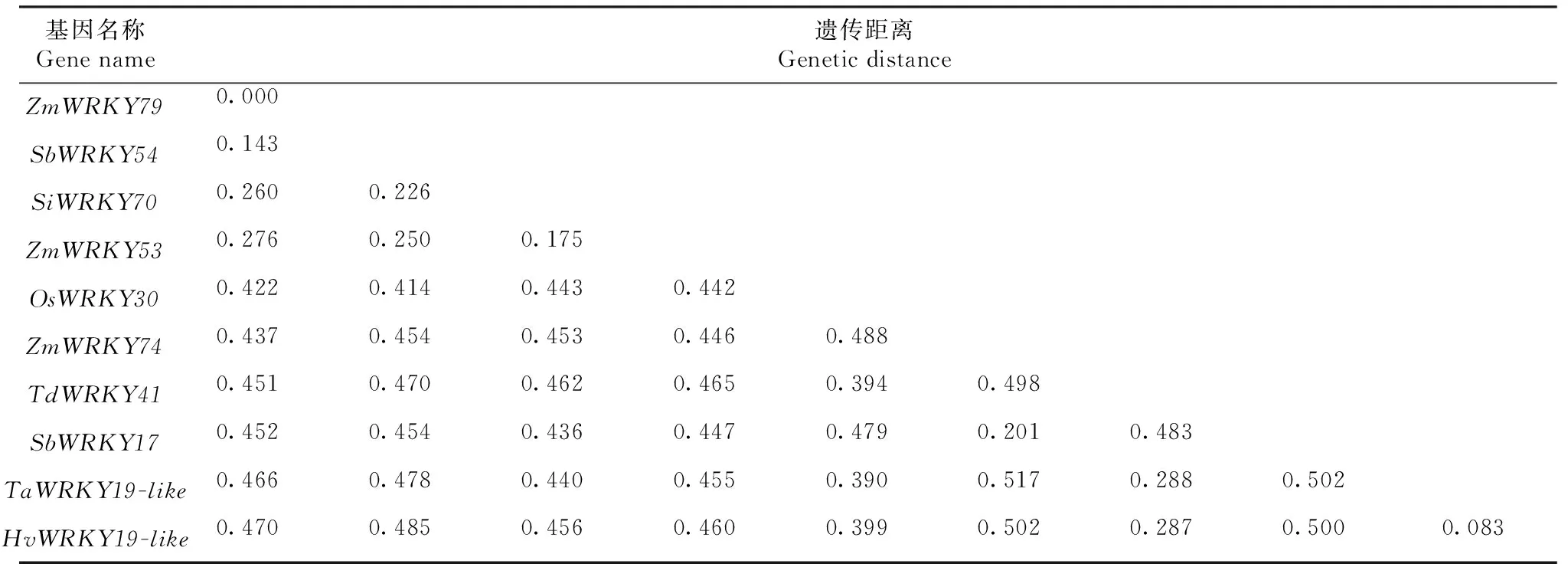
表2 ZmWRKY79同源基因的遗传距离Table 2 Genetic distance of ZmWRKY79 homologous genes

图5 ZmWRKY79蛋白同源序列的系统进化树Fig.5 Phylogenetic tree of ZmWRKY79 protein homologous sequences
2.3.5ZmWRKY79同源基因进化选择压力和亚细胞定位预测 为进一步明确ZmWRKY79同源基因的进化情况,利用DnaSP6对供试的10条核酸序列进行进化选择压力分析,从而获得Ka(非同义替换率)和Ks(同义替换率)(表3),结果显示本研究中的9个成员相对于ZmWRKY79基因,Ka小于Ks,Ka/Ks比值小于1,由此说明ZmWRKY79同源基因成员间在进化过程中以纯化选择为主。利用Crop-PAL对参试的10个蛋白进行亚细胞定位分析(图6),结果显示10个成员均定位在细胞核,由此说明该基因的成员主要在细胞核发挥功能。

图6 ZmWRKY79同源基因的亚细胞定位Fig.6 Subcellular localization of ZmWRKY79 homologous genes
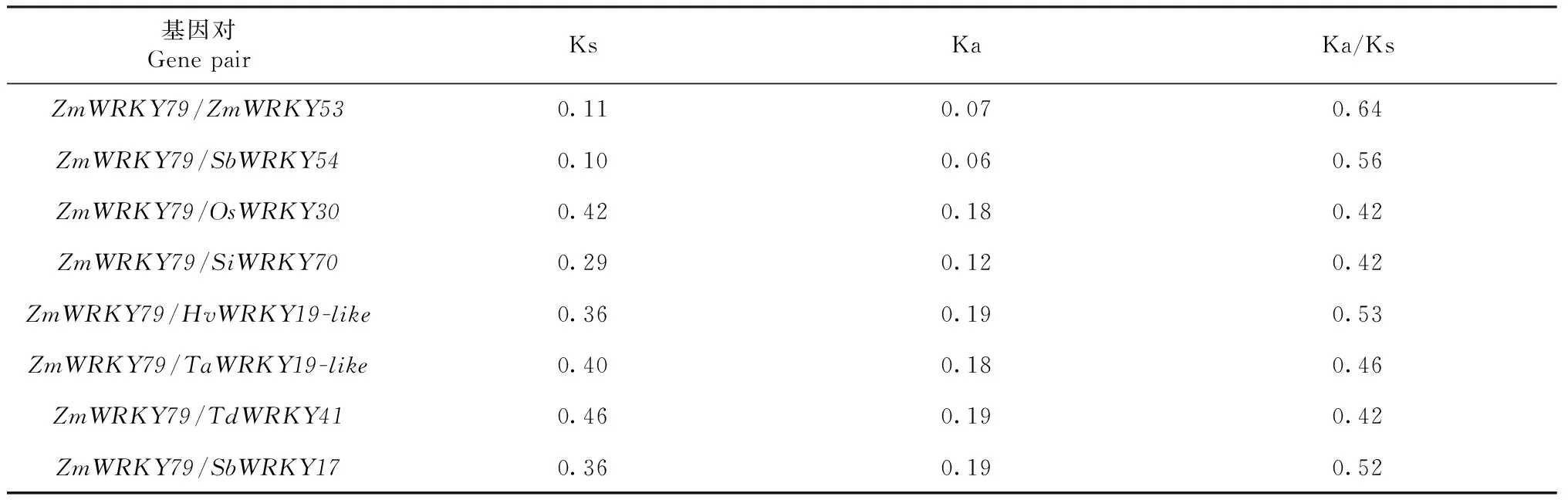
表3 ZmWRKY79同源基因进化选择压力Table 3 Evolutionary selection pressure of ZmWRKY79 homologous gene
2.4 ZmWRKY79同源基因启动子分析
将从NCBI的Gene数据库获取的ZmWRKY79同源基因编码区域上游1500 bp核苷酸序列作为启动子序列。利用在线分析软件PlantCARE预测启动子区域顺式作用调控元件(图7)。结果显示,ZmWRKY79基因及其同源基因拥有多个顺式作用元件,主要分为4类:组织特异性元件(B、C、E、G、O)、应激响应元件(H、K、T)、激素响应元件(D、F、L、M、R、S)、其它顺式作用元件(A、I、J、N、P、Q)。值得注意的是ZmWRKY79同源基因的启动子中均具有多个MeJA响应元件和脱落酸响应元件,由此说明,ZmWRKY79同源基因可能像ZmWRKY79一样可参与激素响应反应。与此同时,大部分ZmWRKY79同源基因拥有MYB干旱胁迫结合位点,说明这些基因可能参与干旱胁迫反应。
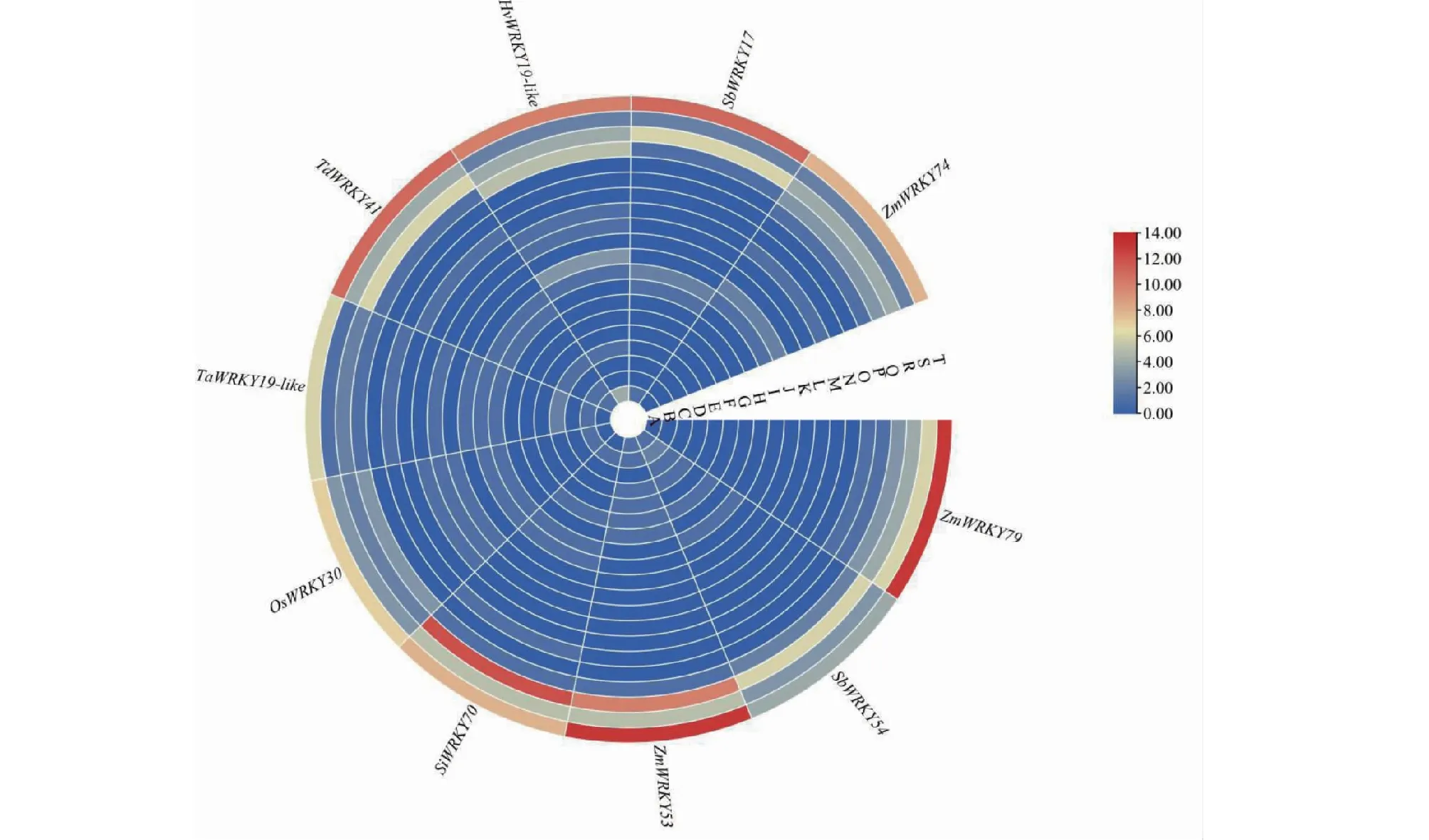
A:转录起始-30区核心启动子元件;B:分生组织表达相关元件;C:分生组织特异性激活相关元件;D:赤霉素响应元件;E:胚乳特异性表达元件;F:参与玉米素代谢调控元件;G:根特异性调控元件; H:低温响应元件;I:MYB干旱胁迫结合位点;J:MYBHv1结合位点;K:防御与应激响应元件;L:水杨酸响应元件; M:生长素响应元件;N:参与昼夜节律调控的顺式作用调节元件;O:种子特异性调控元件;P:ATBP-1结合位点;Q:厌氧诱导元件;R:MeJA响应元件;S:脱落酸响应元件;T:光响应元件。A:Core promoter element around -30 of transcription start; B:Meristem expression related element; C:Cis-acting regulatory element related to meristem specific activation; D:Gibberellin response element; E:Endosperm-specific expression element; F:Cis-acting regulatory element involved in zein metabolism regulation; G:Root-specific regulatory elements;H:Low temperature response element; I:Drought stress binding site for MYB; J:MYBHv1 binding site; K:Defensive and stress response element; L:Salicylic acid response element; M:Auxin response element; N:Cis-acting regulatory element involved in circadian control; O:Seed specific regulatory element; P:ATBP-1 binding site; Q:Anaerobic induction element; R:MeJA response element; S:Abscisic acid response element; T:Light response element.图7 ZmWRKY79的同源基因启动子区域顺式作用元件预测Fig.7 Predicted cis-acting elements in the promoter region of ZmWRKY79 homologous genes
2.5 非生物胁迫下ZmWRKY79同源基因表达情况
将参试的10个基因输入qTeller数据库,共获得ZmWRKY53、ZmWRKY74和ZmWRKY79共3个成员在各处理(高温胁迫、低温胁迫、盐胁迫、紫外线胁迫、干旱胁迫)下的转录组数据(图8)。结果显示,3个成员中,ZmWRKY79基因受多种非生物胁迫诱导表达,其在高温、低温、高盐、紫外线、干旱处理下均显著上调表达,而ZmWRKY53只在低温胁迫下上调表达,ZmWRKY74在低温和紫外线胁迫下上调表达。
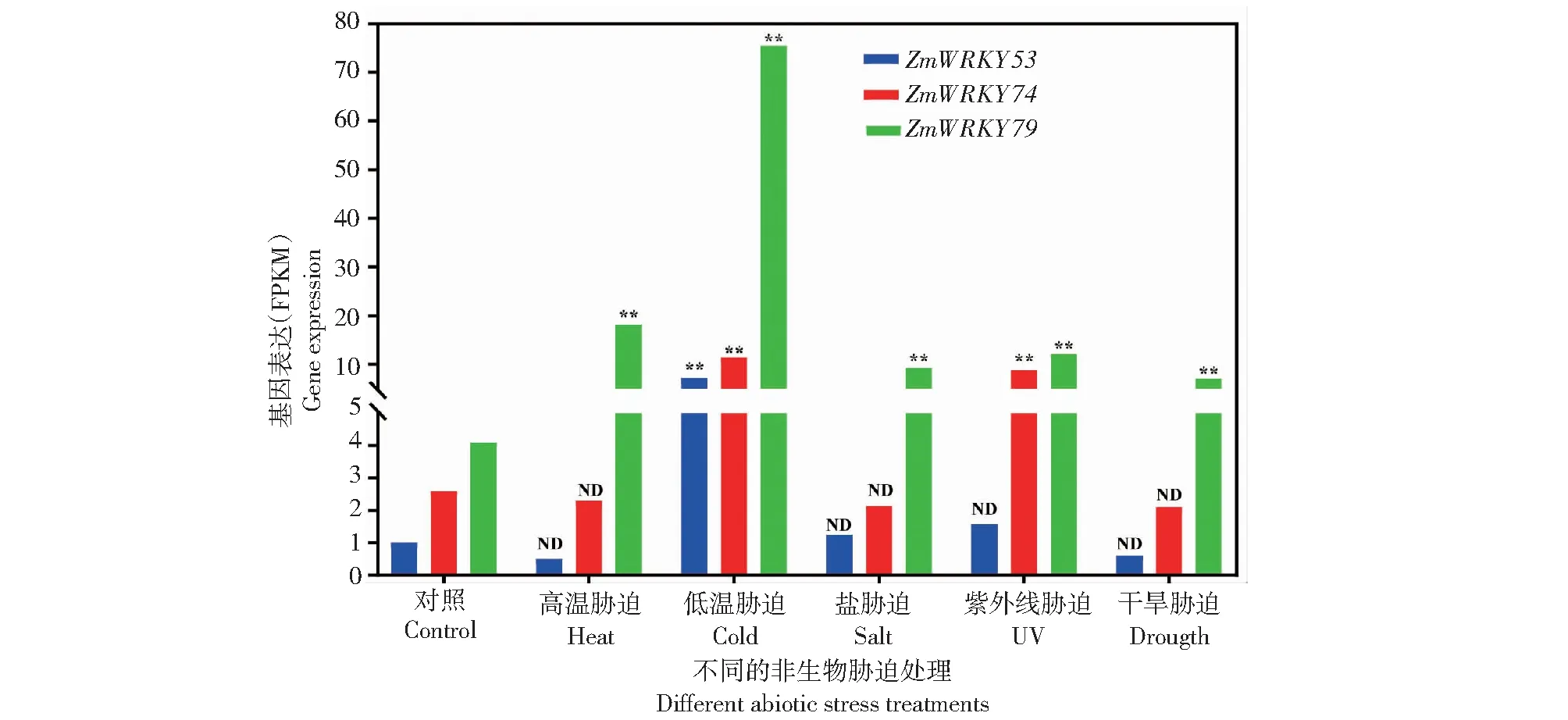
ND代表没有差异;**代表存在显著性差异。ND represents no difference; ** represents the existence of significant indigenous differences.图8 非生物胁迫下ZmWRKY79基因同源基因表达Fig.8 Expression of ZmWRKY79 homologous genes under abiotic stress
3 讨 论
3.1 不同禾本科作物中存在玉米ZmWRKY79基因的同源基因
全球每年因非生物胁迫导致农作物产量减少一半,生物胁迫导致农作物产量减少约20%[11]。禾本科作物作为农业生产中不可替代的角色,也会遭受各种胁迫导致产量的减少,其中非生物胁迫的影响要远远大于生物胁迫[4,6]。WRKY转录因子能够在植物遭受到外界胁迫时调动机体防御系统来适应生境[4,6]。玉米作为重要的粮饲作物,常年遭受极端天气和病原菌的胁迫,产量和品质深受影响[22]。ZmWRKY79在玉米响应逆境胁迫中发挥着重要作用。然而,在一般情况下,同源基因成员间功能可能基本相似,为此本研究通过生物信息学方法在不同物种中寻找ZmWRKY79基因的同源基因,以期获得更多具有利用潜力的WRKY基因。ZmWRKY79基因在数据库中的检索结果显示,在玉米、水稻、高粱、小米、青稞、小麦中找到9个ZmWRKY79基因的同源基因,这说明不同禾本科作物中存在玉米ZmWRKY79基因的同源基因,也预示着这些基因可能发挥着类似的功能。本研究挖掘到的这些基因可作为农作物非生物逆境抗性的重要基因资源。
3.2 ZmWRKY79及其同源基因可能发挥着相似的功能
本研究获得的9个ZmWRKY79同源基因,在功能上存在多样化,这可能与锌指结构域有着密切关系,相关研究表明,锌指结构增加了WRKY蛋白表达的多样性,从而可能使WRKY蛋白在植物防御过程中功能有所不同[4]。本研究发现ZmWRKY79及其9个同源基因的WRKY功能结构域相对保守,七肽基序完全一致,但其中的锌指结构域却存在大量变异,通过聚类分析发现,10个蛋白的聚类也与锌指结构区域相关,由此说明锌指结构区域可能是ZmWRKY79及其同源基因的功能位点。相关研究表明,WRKY家族基因在进化过程中经历了全基因组的加倍和串联复制,这些基因往往成簇分布在染色体上[21]。染色体定位显示ZmWRKY79同源基因分布在不同的染色体上,由于目前鉴定到的ZmWRKY79同源基因成员较少,这些基因在染色体上分布呈不均匀状态,存在热点区域,并未成簇分布,预示还可能存在新的ZmWRKY79同源基因。亚细胞定位结果显示ZmWRKY79同源基因均显示在细胞核,这与Fu等[11]研究类似。利用生物信息学方法预测WRKY转录因子的基本信息及生物学功能极大地缩减了研究时间[23]。然而,往往遗传距离较近的WRKY蛋白拥有近似的生物学功能[6,21]。启动子分析结果显示,ZmWRKY79同源基因的启动子中均具有激素响应调控元件,由此推测这些基因在受到外界胁迫时通过调控体内激素合成来对其响应。非生物胁迫表达结果显示,ZmWRKY53、ZmWRKY74、ZmWRKY79均会受到低温胁迫而上调表达。综上所述,本研究鉴定的这些基因可能对一种甚至多种非生物逆境具有抵御功能。今后的研究将聚焦于这些基因的作用机制研究,加速ZmWRKY79同源基因在育种中运用的进程。
4 结 论
本研究以ZmWRKY79基因作为研究对象,利用生物信息学方法从不同的禾本科作物中鉴定ZmWRKY79的同源基因,同时也对这些基因进行序列比对、结构分析、进化分析、启动子调控元件预测、逆境胁迫下的转录分析,结果显示ZmWRKY79同源基因可能具有抗逆功能,这将为分子育种提供更多的选择,并为挖掘更多ZmWRKY79的同源基因功能奠定理论基础。
猜你喜欢
华人时刊(2023年1期)2023-03-14
南方医科大学学报(2022年3期)2022-04-13
昆明医科大学学报(2022年1期)2022-02-28
汉字汉语研究(2021年2期)2021-08-30
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10
实用临床医药杂志(2021年13期)2021-01-10
三农资讯半月报(2020年15期)2020-08-25
汉字汉语研究(2019年2期)2019-08-27
河北书画研究(2016年3期)2016-04-28
中华胰腺病杂志(2015年5期)2015-12-08
