
霞多丽葡萄果实降异戊二烯香气物质积累及代谢酶活变化分析
2023-05-04 05:30:24崔媛媛陈春霞冯丽丹李霁昕把灵珍孔祥锦蒋玉梅
浙江农业学报 2023年4期
张 祯,崔媛媛,陈春霞,冯丽丹,赵 勇,李霁昕,把灵珍,孔祥锦,张 煜,蒋玉梅,*
(1.甘肃农业大学 食品科学与工程学院,甘肃省葡萄与葡萄酒工程学重点实验室,甘肃省葡萄与葡萄酒产业技术中心,甘肃 兰州 730070; 2.甘肃莫高实业发展股份有限公司生态农业示范种植园区,甘肃 武威 733006)
香气是衡量葡萄和葡萄酒品质的重要指标,葡萄酒的品种香主要来自葡萄果实,葡萄果实香气在葡萄的生理生化代谢过程中生成积累,受葡萄品种、生长环境、气候、栽培管理模式和成熟度等因素的影响[1]。目前葡萄果实中已确定的香气物质有600多种,包括醛、醇、酯、酮、萜类和吡嗪等多类物质[2-3]。其中降异戊二烯香气物质阈值较低,通常具有苹果、覆盆子、木瓜、紫罗兰等花果香气特征,是霞多丽葡萄特征香气的重要组成,主要由类胡萝卜素在类胡萝卜素裂解双加氧酶(carotenoid cleavage dioxygenase,CCD)的酶促或非酶作用下降解代谢生成[4],包括C9、C10、C11和C13降异戊二烯香气物质,以游离态和糖苷结合态2种形式存在[5]。
CCD是类胡萝卜素酶促降解代谢生成降异戊二烯香气物质的关键酶[6]。研究发现葡萄品种[7]、栽培措施[1]、生长环境[8]和成熟度[9]等因素均会影响降异戊二烯香气物质的积累。玫瑰香和黑比诺葡萄发育过程中降异戊二烯香气物质总量整体呈先升后降趋势,结合态含量高于游离态[9-10]。目前葡萄果实中发现的CCD基因家族成员有VvCCD1、VvCCD4a、VvCCD4b、VvCCD7和VvCCD8,其中VvCCD1、VvCCD4a和VvCCD4b能够催化环状和链状类胡萝卜素,生成降异戊二烯香气物质[11-12]。藤稔葡萄生长发育过程中总类胡萝卜素含量与VvCCD基因转录水平有一定的相关性[13]。基于降异戊二烯香气物质在葡萄和葡萄酒香气构成中的重要性,近二十年来,众多学者开始关注研究葡萄中降异戊二烯物质的构成、代谢和影响因素[14]。但是关于霞多丽葡萄果实生长期降异戊二烯香气物质积累及代谢酶活变化分析研究目前鲜见报道。
试验在2020和2021年两个连续生产季,以霞多丽葡萄为试材,于果实幼果(花后10 d)至成熟(花后80 d)期间每2周采集样品,测定样品理化指标、总类胡萝卜素含量、降异戊二烯香气物质含量和CCD活性,分析类胡萝卜素含量、降异戊二烯香气物质含量和CCD活性三者间的相关性,探讨降异戊二烯香气代谢前体和酶活性对其生成的影响,确定霞多丽葡萄果实生长期间降异戊二烯香气物质的积累规律。以期为霞多丽葡萄降异戊二烯香气代谢机理研究和香气品质调控提供科学数据参考。
1 材料与方法
1.1 材料与试剂
霞多丽(Chardonnay)葡萄:2010年定植,砧木5BB,单干单臂架形,南北行向,株行距0.6 m×2 m,每行20株,规范化管理,种植于甘肃省兰州市安宁区甘肃农业大学葡萄产业技术研发中心葡萄品种园(103°41′E,36°5′N;海拔1 530 m)。兰州市2020年年均温为11.2 ℃,年降水量为367 mm;2021年年均温为14.1 ℃,年降水量为341 mm。气象数据从中国气象数据共享服务系统(http://cdc.cma.gov.cn/home)查询获得。
标准品:6-甲基-5-庚烯-2-酮、α-紫罗兰酮、β-紫罗兰酮、β-环柠檬醛、β-大马士酮、香叶基丙酮和2-辛醇等标品购于美国Sigma Aldrich公司;类胡萝卜素裂解双加氧酶(CCD)ELISA试剂盒购于江苏科晶生物科技有限公司;糖苷酶AR2000购于Creative Enzymes公司;固相萃取柱(Cleanert PEP-SPE,150 mg/6 mL)购于天津博纳艾杰尔科技有限公司。
1.2 仪器与设备
Thermo Scientific 265079气相色谱-质谱联用仪(GC-MS),美国Thermo Scientific公司;色谱柱,DB-WAX(60 m×2.5 mm×0.25 μm),美国Thermo Scientific公司;固相微萃取头(SPME)Fiber,DVB/CAR/PDMS(50/30 μm),美国Surpelco公司。
1.3 方法
1.3.1 样品采集
2020和2021年6—8月,选择树势、负载量基本一致的3行葡萄植株,花后10 d(幼果)至花后80 d(成熟),每隔2周,上午8:00—10:00采样,逢雨天隔天采。采样排除边缘葡萄藤,兼顾阴阳面和葡萄叶幕内,随机摘取3~10穗葡萄,去除果梗,混匀,取100粒进行理化指标测定,其余浆果根据检测要求处理液氮速冻,-80 ℃贮藏待测。
1.3.2 理化品质分析
参照王福荣[15]的方法测定葡萄百粒重、可溶性固形物含量、可滴定酸含量(以酒石酸计)和pH值。
1.3.3 总类胡萝卜素含量测定
参照Leng等[13]的方法,准确称取0.500 0 g葡萄果皮样品,加10 mL 95%乙醇匀浆,10 000 r·min-1(4 ℃)离心15 min。取0.5 mL上清液加4.5 mL 95%乙醇。分光光度计测定664 nm、649 nm和470 nm处吸光值,计算总类胡萝卜素含量。
1.3.4 CCD活性测定
葡萄果实加液氮研磨成粉,称取0.200 0 g粉末,置于1.5 mL离心管,加入pH 值7.4 磷酸盐缓冲液(1 g∶4 mL),摇匀,4 000 r·min-1(4 ℃)离心15 min,取上清液,ELISA试剂盒测定活性。
1.3.5 香气物质提取
游离态香气。样品50 g解冻、去籽、去果蒂,制备葡萄醪。
结合态香气。参照刘斌[3]的方法并修改。50 g葡萄果实,去籽,去果蒂,液氮速冻后,加入1 g PVPP和0.5 gD-葡萄糖酸内酯于组织捣碎机打粉,置于50 mL离心管4 ℃冰箱静置2 h,8 000 r·min-1(4 ℃)离心10 min。取2 mL澄清葡萄汁过活化后的固相萃取柱,依次用2 mL水和5 mL二氯甲烷洗脱去除低分子量的糖、酸和游离态香气等化合物,20 mL甲醇洗脱结合态香气物质收集至50 mL圆底烧瓶,30 ℃真空旋转蒸发至干。加10 mL柠檬酸/磷酸缓冲液(0.2 mol·L-1,pH值5)溶解,转移至含有0.1 g糖苷酶AR2000的密闭离心管,37 ℃恒温酶解16 h,制得结合态香气物质样品。
1.3.6 香气物质分析
参照王雨等[16]的方法并修改,准确称取5.00 g样品于20 mL样品瓶中,加入1.00 g NaCl,10 μL内标(2-辛醇,0.5 mg·kg-1)及磁力转子,40 ℃水浴磁力搅拌平衡30 min,插入活化后的SPME萃取头40 ℃顶空萃取30 min,气相色谱进样口230 ℃解析10 min进样。
色谱条件:进样口温度230 ℃;不分流进样;载气(高纯He),流速1 mL·min-1;升温程序:50 ℃(10 min),3 ℃·min-1升至180 ℃(6 min)。
质谱条件:电子轰击离子源(EI);电子能量70 eV;连接杆温度180 ℃;离子源温度250 ℃;质谱扫描范围m/z50~350。
1.3.7 定性与定量分析
定性:外标法结合NIST谱库检索和相对保留指数(RI)定性;
定量:外标定量。标准品样品制备加入内标2-辛醇,消除基质影响。标准曲线方程见表1。

表1 降异戊二烯香气物质标准曲线方程Table 1 Standard curve equation of norisoprenoids
1.4 数据处理
Microsoft Excel 2016统计,SPSS Statistics 25.0显著性分析(Duncan法,P<0.05),Origin 2018绘图,Metabo Analyst 5.0进行正交偏最小二乘法判别分析(OPLS-DA)和相关性分析(Pearson法),所有处理均做3个生物样本重复。
2 结果与分析
2.1 葡萄果实生长期理化指标的变化
葡萄的成熟度可通过可滴定酸含量、pH值、可溶性固形物含量等指标进行评估[17],霞多丽葡萄在花后38~52 d可滴定酸含量开始下降、可溶性固形物含量开始上升、百粒重缓慢增长(图1-A、C、D),符合葡萄转色期糖酸积累和果实生长规律[17],该时期为转色期,进而确定花后10~38 d试验果实处于幼果期,花后52~80 d试验果实处于成熟期。
随着葡萄成熟,样品果实可滴定酸含量呈先升后降趋势(图1-A),花后38 d达到峰值,2020和2021年分别为30.41 g·L-1和27.27 g·L-1,花后52 d样品果实进入成熟期可滴定酸急剧下降,成熟果实样品(花后80 d)可滴定酸下降至4.91 g·L-1(2020年)和4.98 g·L-1(2021年)。样品pH值呈先降后升趋势(图1-B),花后38 d达到最低值(2020年2.49;2021年2.60),花后52 d后成熟期急剧增加,成熟果实(花后80 d)上升至3.63(2020年)和3.64(2021年),变化趋势与可滴定酸含量变化相符。
可溶性固形物含量(图1-C)和百粒重(图1-D)在果实生长期间均呈上升趋势。可溶性固形物含量在成熟期(花后52~80 d)急剧增加,成熟果实(花后80 d)两个生产季的可溶性固形物含量分别为20.32oBrix(2020年)和22.30oBrix(2021年);百粒重在转色期(花后38~52 d)增加缓慢,花后10~24 d、52~80 d果实先后两次膨大,百粒重显著增加,2020年分别增加了82.47%、85.50%,2021年分别增加了174.00%、57.01%,成熟果实(花后80 d)两个生产季的百粒重分别为111.19 g(2020年)和81.66 g(2021年)。霞多丽葡萄果实在幼果期和成熟期均膨大,幼果期果肉细胞快速分裂,体积快速膨大,果实硬度大,以积累有机酸为主,果肉细胞开始积累淀粉,该时期结束时有机酸含量达到最大,pH值最小;成熟期果实体积再次迅速增大,果实逐渐变软,酸度下降,果实细胞液泡开始积累糖分,葡萄糖和果糖含量迅速增加,二者比例接近1[18],试验果实的可溶性固形物含量、可滴定酸含量、百粒重和pH值变化与李秀杰等[18]报道的葡萄基本理化指标变化规律相符。
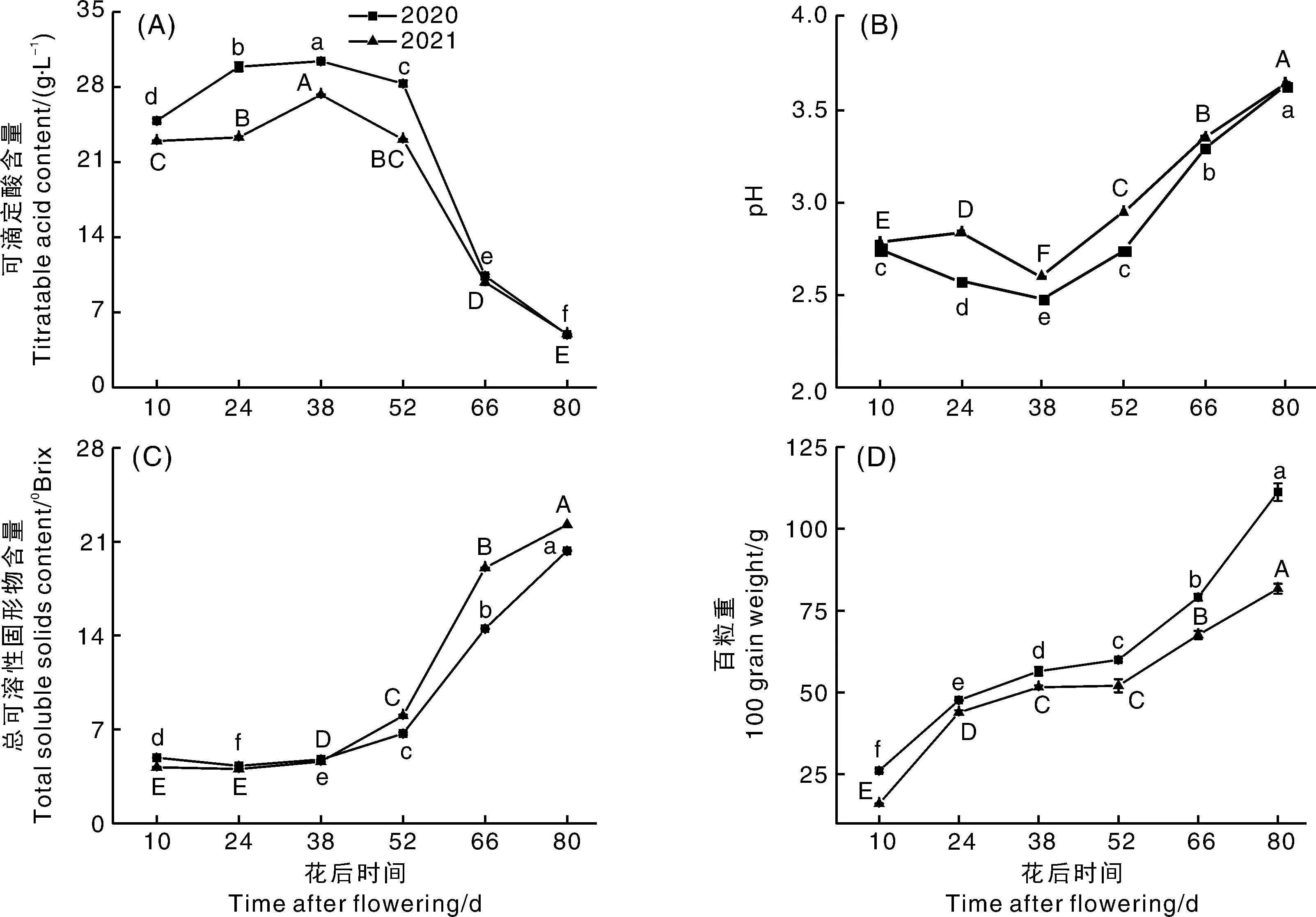
A,可滴定酸含量;B,pH值;C,可溶性固形物含量;D,百粒重。不同小写字母表示2020年不同时期之间差异显著(P<0.05);不同大写字母表示2021年不同时期之间差异显著(P<0.05),下同。A, Titratable acid content; B, pH; C, Soluble solids content; D, 100-grain weight. Different lowercase letters indicate significant differences between different periods in 2020 (P<0.05); different capital letters indicate significant differences between different periods in 2021 (P<0.05). The same as below.图1 霞多丽葡萄果实发育过程中理化指标变化Fig.1 Changes of physical and chemical indexes during development of Chardonnay grape berries
2020和2021年两个连续生产季霞多丽葡萄可滴定酸含量、pH值、可溶性固形物含量和百粒重的变化趋势一致,但含量存在差异。成熟果实(花后80 d)2021年百粒重较2020年降低了26.56%,可能与生产地2021年较2020年降水量少有关,张晓霞[19]研究发现,在葡萄膨大期进行调亏灌溉会导致果实粒径减小,百粒重降低;2020年可溶性固形物含量较2021年降低了8.88%,可能是由于2020年浆果体积较大而产生的稀释作用[20];可滴定酸含量、pH值差异不大,表明生产季气候条件对其积累的影响不大。
2.2 葡萄果实总类胡萝卜素含量的变化
总类胡萝卜素含量整体呈下降趋势,但转色期前后会有一个生成积累小高峰(图2)。幼果期果实膨大过程(花后10~24 d)总类胡萝卜素含量快速下降(2020年30.50%,2021年47.04%),这可能是稀释效应[19]所致;随后总类胡萝卜素含量增加(2020年花后24~38 d;2021年花后24~52 d),2020年峰值(71.96 μg·g-1)出现在转色期前(花后38 d),2021年峰值(46.68 μg·g-1)出现在转色期后(花后52 d)。有学者发现[21],类胡萝卜素异构酶基因(VvOR)在葡萄果实转色期有最低或最高的表达量,VvOR基因影响了类胡萝卜素合成,使果实总类胡萝卜素含量在转色前期或后期出现峰值,试验结果符合此规律。成熟期样品果实总类胡萝卜素含量呈下降趋势,至果实成熟总类胡萝卜素含量2020和2021年分别较峰值下降了48.22%和57.01%。可能是由于果实成熟期八氢番茄红素合成酶基因(VvPSY1)表达量降低,VvCCD1和9-顺式环氧类胡萝卜素双加氧酶基因(VvNCED2)表达量增高[13],类胡萝卜素合成速率降低,降解加快所致。
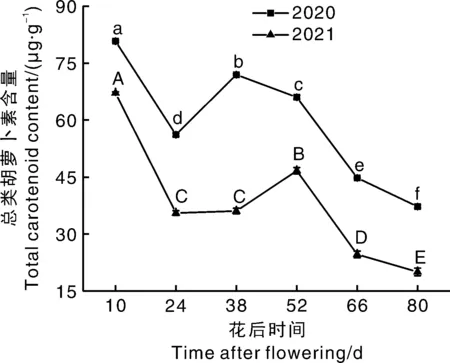
图2 霞多丽葡萄果实发育过程中总类胡萝卜素含量的变化Fig.2 Changes of total carotenoid content during development of Chardonnay grape berries
2020年葡萄生长过程中总类胡萝卜素含量始终高于2021年,说明生产季气候对类胡萝卜素积累影响较大。葡萄类胡萝卜素稳态水平取决于生物合成和降解之间的平衡[22],类胡萝卜素生物合成和降解代谢速率与代谢酶活性大小相关,而酶活性又受年份因素(光照、温度等)影响。有学者研究发现,强光照能够提高PSY、CCD等基因的表达量,从而影响果实中类胡萝卜素的合成或降解[7,23]。通过中国国家气象数据分享系统(http://cdc.cma.gov.cn)检索兰州两个生产季气温和光照发现,2021年生产季较2020年的温度高、光照强,这可能影响了果实的CCD活性,后续试验发现,2021年样品CCD活性显著高于2020年样品,2021年样品总类胡萝卜素含量较2020年低,可能是样品CCD活性较高,总类胡萝卜素降解量增加的结果。
2.3 葡萄果实CCD活性的变化分析
由图3可知,幼果膨大期试验果实CCD活性显著下降(2020年13.46%,2021年11.01%),植物中CCD定位于胞质或质体[7],幼果膨大期果实细胞处于快速分裂阶段,质体数目维持在低水平[24],CCD整体活性降低;幼果后期试验果实CCD活性上升。转色期样品CCD活性再次下降。果实进入成熟期,样品CCD活性开始上升,花后66 d开始快速升高,果实成熟时(花后80 d)上升至2020年430.99 U·g-1,2021年872.13 U·g-1。Leng等[13]发现,VvCCD1和VvCCD4a均在果实成熟期表达量增加,其中VvCCD1在成熟期达到最大值,与本研究结果一致。
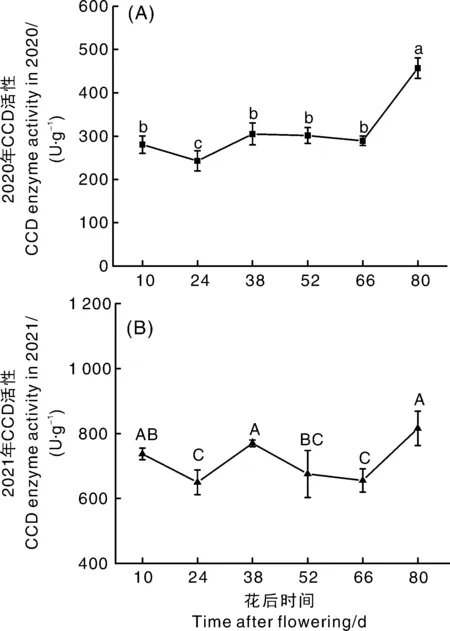
A,2020年CCD活性;B,2021年CCD活性。A, CCD activity in 2020; B, CCD activity in 2021.图3 霞多丽葡萄果实发育过程中CCD活性的变化Fig.3 Changes of CCD activity during development of Chardonnay grape berries
两年CCD活性变化趋势基本一致,但2021年活性明显高于2020年,可能是由于兰州2021年的气温和光照强度较2020年高,降雨量少(http://cdc.cma.gov.cn)导致。Wang等[25]研究发现,脱水胁迫可提高CCD1基因表达量;降雨量少,果实水分亏缺可提高CCD转录丰度[26]。
2.4 降异戊二烯香气物质总量的变化分析
霞多丽果实样品中定性定量6种游离态和结合态降异戊二烯香气物质,其中6-甲基-5-庚烯-2-酮是C8降异戊二烯香气物质,β-环柠檬醛是C10降异戊二烯香气物质,α-紫罗兰酮、β-紫罗兰酮、β-大马士酮和香叶基丙酮是C13降异戊二烯香气物质。两个生产季所检测出的游离态和结合态组分种类相同,但含量不同,表明生产季气候条件差异不会改变降异戊二烯香气物质的种类,但会影响降异戊二烯物质的含量。
样品果实降异戊二烯香气物质总量在幼果期和转色期两年变化趋势有差异(图4),但成熟期变化趋势一致(先降后升),果实成熟时(花后80 d)总量达到16.05 μg·kg-1(2020年)和39.04 μg·kg-1(2021年)。试验成熟期CCD活性也呈先降后升趋势,说明成熟期降异戊二烯香气物质总量变化与CCD活性有一定的相关性,与Meng等[27]研究结果一致。
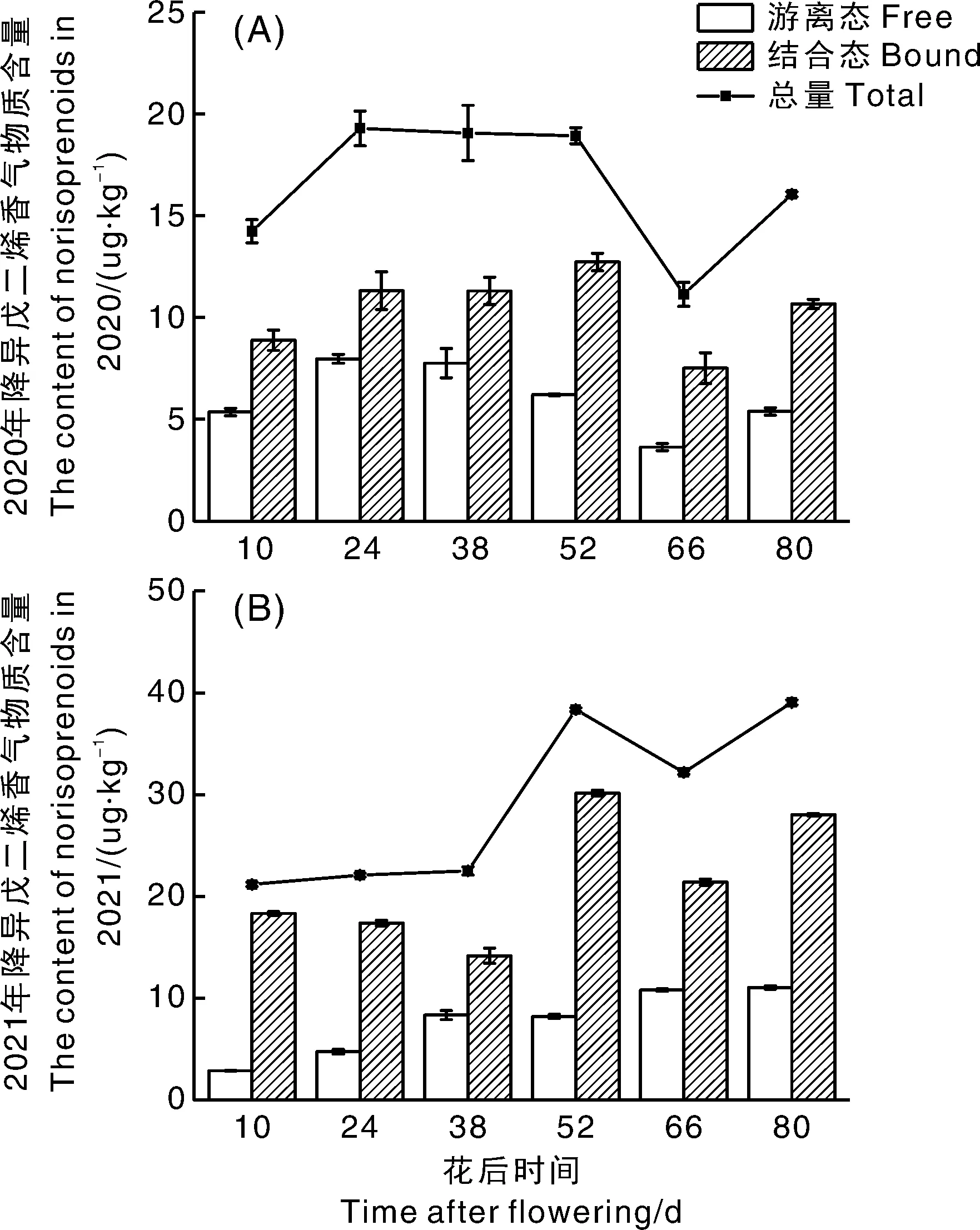
A,2020年降异戊二烯香气物质含量;B,2021年降异戊二烯香气物质含量。A, The content of norisoprenoids in 2020; B, The content of norisoprenoids in 2021.图4 霞多丽葡萄果实发育过程中降异戊二烯香气物质含量的变化Fig.4 Changes of norisoprenoids during development of Chardonnay grape berries
两个生产季结合态降异戊二烯香气物质含量变化趋势在果实幼果期相反,转色期和成熟期一致,但都在花后52 d转色期结束时达到峰值(2020年,12.73 μg·kg-1;2021年,30.13 μg·kg-1)。2021年结合态降异戊二烯香气物质含量高于2020年,2021年生产季光照较2020年高,Bureau等[28]研究发现,光照能刺激果实类胡萝卜素分解,促进结合态降异戊二烯香气物质合成,这可能是导致两个生产季结合态降异戊二烯物质含量差异的原因。
幼果期游离态降异戊二烯香气物质含量在两个生产季均呈上升趋势;转色期2020年下降迅速,2021年下降幅度小;成熟期2020年先降后升,成熟果实含量为5.38 μg·kg-1,2021年持续上升至11.04 μg·kg-1。就含量而言,2021年转色期和成熟期游离态降异戊二烯香气物质含量比2020年高,可能是由于香气物质主要分布于果皮[29],2021年该时期气温较高、降雨量少,百粒重小于2020年样品,果皮果肉比增大,这可能是2021年样品降异戊二烯香气物质含量较高的原因之一。
2.5 降异戊二烯香气物质组分的变化分析
为了直观反映不同生产季和不同成熟度对葡萄果实降异戊二烯香气组分的影响,以游离态和结合态降异戊二烯香气组分含量为变量,进行了正交偏最小二乘判别分析(OPLS-DA)(图5)。由图5-A可知,两个主成分可以反映总体66.6%的原始变量信息,其中RX2、RY2、Q2分别为0.905、0.574和0.462,由图5-C可知,两个主成分可以反映总体77.5%的原始变量信息,其中RX2、RY2、Q2分别为0.855、0.982和0.975,说明这两个模型可以反映游离态和结合态降异戊二烯香气组分的总体情况,且模型稳定。图5-A、C可以将不同生产季样品清楚地分开,表明不同生产季果实中降异戊二烯香气组分含量有较大差别,尤其是结合态香气组分(图5-C),2020年样品分布于X轴的负半轴,2021年样品分布于正半轴,虽然样品点分布随果实成熟未呈现规律性分布,但大部分样品点可以较好地分离。从图5-B、D样品得分图可以看出,生产季气候对游离态和结合态降异戊二烯香气组分(得分>1.0)的影响有所不同,生产季气候对游离态β-紫罗兰酮和结合态β-大马士酮的积累影响最大。
对2个生长季游离态β-紫罗兰酮和结合态β-大马士酮的积累动态进一步进行分析,结果见图5-E、F。2个生产季游离态β-紫罗兰酮在幼果期含量呈下降趋势,转色期(花后38~52 d)含量上升,2020年花后52 d较38 d含量上升了86.67%,2021年花后52 d含量是花后38 d的6.08倍。成熟期2个生产季的游离态β-紫罗兰酮含量变化不一致。2020年游离态β-紫罗兰酮含量持续显著下降至0.08 μg·kg-1;2021年游离态β-紫罗兰酮含量在花后52~66 d显著上升并达到最大值(2.80 μg·kg-1),随后含量显著下降至1.50 μg·kg-1。2021年转色期和成熟期β-紫罗兰酮含量高于2020年,可能与2021年气温较2020年高,降雨量少,果实体积脱水缩小,香气浓度上升[29]有关。β-大马士酮是C13-降异戊二烯香气物质中贡献最大的组分[30],在果实生长发育过程中,2020年结合态β-大马士酮含量无显著变化(P>0.05);2021年β-大马士酮幼果期含量显著上升,转色期缓慢下降,成熟期含量上升并达到最大值(2.03 μg·kg-1),成熟后期又开始显著下降(P<0.05)。β-大马士酮的积累受温度、日照时长等因素的强烈影响[31],较高气温有利于β-大马士酮的生成[32],2021年兰州气温高于2020年,因此导致2021年幼果期至成熟期的β-大马士酮含量远高于2020年。
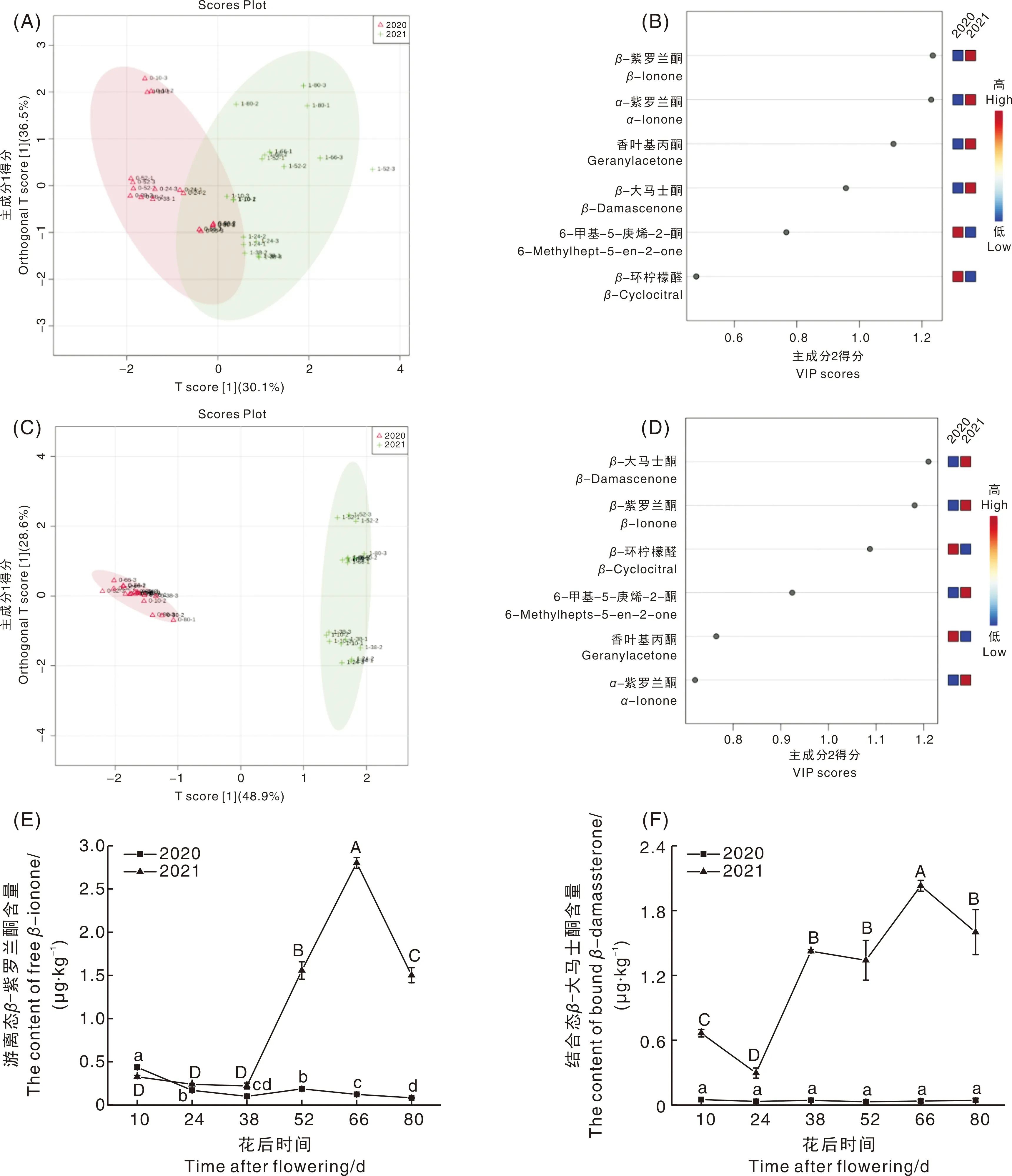
A、B,游离态降异戊二烯香气组分OPLS-DA分析;C、D,结合态降异戊二烯香气组分OPLS-DA分析;E,果实发育过程中游离态β-紫罗兰酮含量变化;F,果实发育过程中游离态结合态β-大马士酮含量变化。A and B, OPLS-DA analysis of free norisoprenoid components; C and D, OPLS-DA analysis of bound norisoprenoid components; E, The changes of free β-ionone content during fruit development; F, The changes of bound β-damascone content during fruit development.图5 霞多丽葡萄果实发育过程中降异戊二烯香气组分OPLS-DA和主要组分的分析Fig.5 Analysis of OPLS-DA and main components of norisoprenoids during development of Chardonnay grape berries
2.6 葡萄果实CCD活性、总类胡萝卜素含量与降异戊二烯香气组分含量相关性分析
CCD催化下,霞多丽果实中定性定量的6-甲基-5-庚烯-2酮主要来自类胡萝卜素中的番茄红素,α-紫罗兰酮主要来自ε-胡萝卜素,β-紫罗兰酮和β-环柠檬醛主要来自β-胡萝卜素生成,香叶基丙酮主要来自ξ-胡萝卜素[12,33]。
CCD活性、总类胡萝卜素含量与降异戊二烯香气组分含量相关性分析显示(图6),总类胡萝卜素含量与CCD活性呈极显著负相关(P<0.01),与Leng等[13]在藤稔葡萄中的研究结果一致。游离态和结合态α-紫罗兰酮含量与CCD活性均呈极显著正相关(P<0.01),与总类胡萝卜素含量分别呈极显著负相关(P<0.01)和显著负相关(P<0.05),与总类胡萝卜素含量相比,结合态α-紫罗兰酮含量与CCD活性相关性更强,说明结合态α-紫罗兰酮生成受CCD活性影响更大。游离态和结合态β-紫罗兰酮含量均与CCD活性呈极显著正相关(P<0.01),与总类胡萝卜素含量呈极显著负相关(P<0.01),说明总类胡萝卜素含量和CCD活性对β-紫罗兰酮的生成具有同等影响。游离态和结合态β-大马士酮含量与CCD活性分别呈显著正相关(P<0.05)和极显著正相关(P<0.01),与总类胡萝卜素含量均呈极显著负相关(P<0.01)。可见,游离态β-大马士酮生成受总类胡萝卜素含量影响更大。游离态香叶基丙酮含量与CCD活性呈极显著正相关(P<0.01),与总类胡萝卜素含量呈极显著负相关(P<0.01),说明CCD活性和总类胡萝卜素含量均会影响游离态香叶基丙酮的生成。结合态6-甲基-5-庚烯-2-酮含量与CCD活性呈极显著正相关(P<0.01),与总类胡萝卜素含量相关性不显著(P>0.05),说明CCD活性是影响结合态6-甲基-5-庚烯-2-酮生成的主要因素,总类胡萝卜素含量对其没有影响。
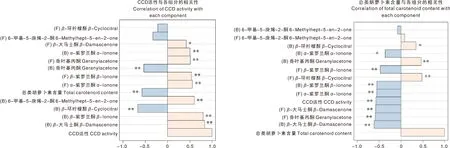
*,显著相关(P<0.05);**,极显著相关(P<0.01); F,游离态;B,结合态。*, Significant correlation (P<0.05); **, Very significant correlation (P<0.01); F, Free state; B, Bound state.图6 CCD活性、总类胡萝卜素含量与降异戊二烯香气组分含量相关性分析Fig.6 Analysis of correlation between CCD activity, total carotenoid content and norisoprenoids content
3 结论
2020—2021年连续两年,通过对不同生长期霞多丽葡萄果实的基础理化指标、总类胡萝卜素含量、CCD活性和降异戊二烯香气物质含量进行测定分析,结果表明,在霞多丽葡萄果实生长发育过程中,可滴定酸含量在转色期达到峰值后开始下降,pH值、可溶性固形物含量和百粒重均呈上升趋势;总类胡萝卜素含量总体呈下降趋势,转色期前后会有一个生成积累小高峰。CCD活性幼果期呈上升趋势,转色期下降,成熟期呈再次上升;果实样品共定性定量6种降异戊二烯香气物质,随着果实成熟,降异戊二烯香气物质总含量呈上升趋势;相关性分析显示,CCD活性与总类胡萝卜素含量呈负相关,游离态和结合态α-紫罗兰酮、β-紫罗兰酮、β-大马士酮以及游离态香叶基丙酮、结合态6-甲基-5-庚烯-2-酮与CCD活性呈正相关,与总类胡萝卜素含量呈负相关。不同生产季样品比较发现,生产季气候对果实理化指标、总类胡萝卜素含量、CCD活性和降异戊二烯香气物质含量均有影响。
猜你喜欢
中学化学(2024年5期)2024-07-08 09:24:57
作物学报(2022年1期)2022-11-05 08:06:34
河北科技师范学院学报(2022年2期)2022-08-26 08:55:32
农业工程学报(2022年7期)2022-07-09 06:16:32
锦州医科大学报(2021年6期)2021-09-10 04:14:43
今日农业(2019年11期)2019-08-15 00:56:32
中学教学参考·文综版(2018年9期)2018-10-23 11:36:12
Atmospheric and Oceanic Science Letters(2016年4期)2016-11-23 03:30:18
橡塑技术与装备(2016年21期)2016-02-25 06:33:12
石油化工(2015年9期)2015-08-15 00:43:05
