
放养密度对寒区稻-扣蟹共作模式中河蟹生长及水质的影响
2023-05-01 13:46:58王世会赵志刚罗亮张瑞郭坤徐伟梁利群张旭彬
水产学杂志 2023年2期
王世会,赵志刚,罗亮,张瑞,郭坤,徐伟,梁利群,张旭彬
(1.中国水产科学研究院黑龙江水产研究所,黑龙江省冷水性鱼类种质资源及增养殖重点开放实验室,黑龙江 哈尔滨 150070;2.中国水产科学研究院盐碱水域渔业程技术研究中心(哈尔滨),黑龙江 哈尔滨 150070;3.黑龙江省水产技术推广总站,黑龙江 哈尔滨 150010)
作为我国目前主推的一项生态循环农业绿色种养模式,稻渔综合种养发展迅速。该种养模式可显著增加经济效益,使种植业与水产养殖业的生态环境服务功能更加明显,是发展“高效、优质、生态、健康、安全”环境友好型水产养殖的有效途径[1]。稻蟹综合种养是稻渔综合种养的重要模式之一,主要分布在我国北方地区,2019 年在全国的应用面积为1.37×105hm2[2]。与水稻单作模式比较,稻蟹综合种养的优势在于可以实现一田两出[3]。目前对稻蟹综合种养模式下的土壤理化性质[4]、底栖生物[5]、浮游动物[6]、稻田水质变化[7,8]、中华绒螯蟹(Eriocheir sinensis)肠道微生物和环境微生物[9]、经济和环境效应评估[10]、氯虫苯甲酰胺类农药的风险评估[11]、甲烷和氧化亚氮排放[12]和水稻品质[13]等均有研究,但针对我国高寒地区(黑龙江省)稻蟹综合种养中华绒螯蟹生长及水质状况研究较少,这不利于全面评估我国稻蟹综合种养模式经济及生态效益。黑龙江省作为我国水稻种植面积第一大省,发展稻蟹综合种养潜力巨大。尽管稻田资源丰富,但黑龙江省地处高寒地区,水产动物在稻田中的生长期短、有效积温低,适合稻渔综合种养的水产动物较少。稻蟹共作模式作为黑龙江省目前主要的稻渔综合种养模式,在生产过程中能够产生良好的综合效益。
中华绒螯蟹俗称河蟹或大闸蟹,主要分布于我国东部沿海通海的河流或湖泊中[14,15],长期地理隔离形成了不同的地理种群,包括辽河、黄河、长江、瓯江和闽江等水系[16]。辽河水系和长江水系中华绒螯蟹是目前已成功开发的稻蟹共作模式的主要养殖种类,辽河水系中华绒螯蟹生长成熟期较短,是目前寒冷地区稻蟹共作模式的主要养殖对象,但其商品规格相对较小,市场价格受限。长江水系中华绒螯蟹因个体较大,市场价格相对较高。已有诸多文献报道了湖泊和池塘养殖模式下不同水系中华绒螯蟹的生长性能[17-19],但在寒区稻蟹共作模式下长江水系中华绒螯蟹生长性能等研究较少。水质理化因子严重影响稻蟹共作的经济和生态效益,探明其在稻蟹综合种养过程中的变化规律,对于解析该模式在高寒地区的生态效益具有重要意义。为此,本研究以长江水系中华绒螯蟹为研究对象,采用放养密度,探究水稻单作及稻-扣蟹共作期间中华绒螯蟹与水稻的生长性能及水质变化状况,以期为我国寒冷地区稻蟹共作模式的科学构建提供技术参数,及为寒冷地区稻田大规格蟹种培育提供理论支撑。
1 材料与方法
1.1 材料及实验地点
实验蟹为长江水系III 期幼蟹,平均体质量为(1.02±0.16)g;实验用水稻品种为“绥粳4”。实验在黑龙江省绥化市北林区稻渔综合种养基地(127.08°E,46.88°N)进行(图1)。灌溉用水源为呼兰河。

图1 实验地点示意图Fig.1 Schematic diagram of experiment location
1.2 实验设计
本研究共设置1 个对照组(水稻单作,RM)和3个处理组(稻蟹共作组,RCC):中华绒螯蟹设3 个密度,分别为22 500 只/hm2(稻蟹共作I,RCCI)、45 000只/hm2(稻蟹共作II,RCCII)和67 500 只/hm2(稻蟹共作III,RCCIII),每个实验单元300 m2,每组3 个重复,共计12 个实验单元。对照组不挖环沟,常规手插秧,插秧行距30 cm,穴距15 cm,每穴3~4 株;处理组在距稻田埂1 m 处开挖梯形环沟,环沟上部宽0.6 m、下部宽0.3 m、深0.4 m,环沟面积不超过稻田总面积的10%。处理组采用“大垄双行,边沟加密”的栽培模式,手插秧,插秧行距40 cm 与行距20 cm 间隔排列,穴距15 cm,每穴3~4 株。借助稻田边际通风透光的优势,在环沟周围两侧增加插秧密度,将环沟占用的水稻补上,保证每个稻田块插秧穴数一致。稻田四周设底端埋入土下20 cm、地面上40 cm 高的防逃网。6 月2 日水稻插秧,7 月6 日投放长江水系III 期幼蟹,每天17:00 给中华绒螯蟹投喂配合饲料(盘锦光合蟹业有限公司),按体质量2%~3%投喂,晴天正常投喂,小雨天少投喂,大雨天不投喂。稻蟹共作期间,每两周上午9:00~11:00采集一次水质样品。
1.3 数据采集、分析及指标测定
1.3.1 水稻和扣蟹生长性能
9 月下旬,统计水稻产量、养成扣蟹平均体质量。随机取100 只养成扣蟹,擦干体表水分后,用电子天平精确称重(精确度=0.01 g),计算增重率(weight gain rate,WGR,%)和特定生长率(specific growth rate,SGR,%/d)。计算公式如下:
式中:Wf为终末扣蟹的体质量(g),Wi为初始III 期幼蟹的体质量(g),D 为养殖时间(d)。
1.3.2 水体和土壤环境因子测定
水体温度(TEMP)、溶解氧(DO)、pH 和电导率(SPC)采用美国维赛多参数水质分析仪(YSI Professional Plus)现场测定。水体总氨氮(TAN)、亚硝酸态氮(NO2-N)、硝酸态氮(NO3-N)、总氮(TN)、正磷酸盐(PO4-P)、总磷(TP)、化学需氧量(COD)和叶绿素a(Chl.a)含量按照水和废水监测分析方法[20]测定,土壤TN、TP 采用相应方法[21,22]测定。
所有数据均采用平均值±标准误(Mean±SE)表示,采用SPSS 22.0 软件统计分析。数据采用单因素方差(ANOVA)分析,Duncan 氏法多重比较,P<0.05 表示差异显著。
2 结果与分析
2.1 中华绒螯蟹生长和水稻产量
三种放养密度中华绒螯蟹的终末体质量、WGR和SGR 均无显著性差异(P>0.05)。稻蟹共作组水稻产量均高于水稻单作产量,且稻蟹共作III 产量显著高于其他处理组(P<0.05)(表1)。

表1 不同处理组中华绒螯蟹的生长参数及水稻产量Tab.1 The growth performance of Eriocheir sinensis and yield of Oryza sativa L.in each treatment
2.2 水环境因子
整个实验期间,水稻单作与稻蟹共作组的TEMP、DO、pH、TAN、NO2-N、NO3-N、TN 和COD 含量均无显著性差异(P>0.05)。稻蟹共作组SPC、TP、Chl.a 含量显著高于水稻单作(P<0.05),水稻单作PO4--P 含量则显著高于稻蟹共作组(P<0.05)。水稻单作组土壤中TN 和TP 含量均高于稻蟹共作组,且TN 含量存在显著性差异(P<0.05)(表2)。

表2 实验期间各处理组水质及土壤参数Tab.2 Water quality and soil parameters in each treatment during the experiment
整个实验期间,稻田水体TEMP 呈现波动性变化(图2),进入8 月份后,TEMP 逐渐下降,9 月2 日降至18.8℃左右。四组稻田TEMP 基本一致,无显著性差异(P>0.05)。pH 呈先下降后上升趋势,但各组略有不同(图3)。8 月5 日水稻单作pH 为5.68,而稻蟹共作I 为5.87,稻蟹共作II 为5.92,稻蟹共作III 为5.99,以上四组间均存在显著性差异(P<0.05)。8 月15 日水稻单作pH 为6.68,稻蟹共作I 为6.84,稻蟹共作II 为6.94,稻蟹共作III 为6.90,水稻单作与稻蟹共作组存在显著性差异(P<0.05),稻蟹共作组内无显著性差异(P>0.05)。9 月2 日稻蟹共作I和稻蟹共作II 无显著性差异(P>0.05),但以上两个处理组与稻蟹共作III 存在显著性差异(P<0.05)。
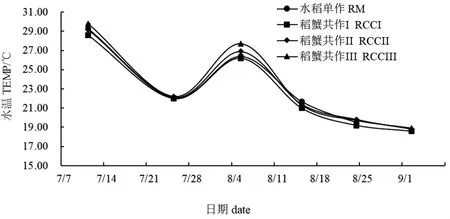
图2 各处理组稻田水体温度变化Fig.2 Changes in paddy field water temperature in each treatment
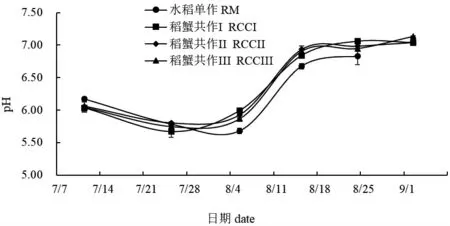
图3 各处理组稻田水体p H 变化Fig.3 Changes in paddy field water pH in each treatment
图4 和图5 分别为各处理组稻田水体DO 和SPC 变化趋势。整体来看,7 月11 日到8 月24 日间,除稻蟹共作I 外,其余各组稻田中水体DO 值均降低,8 月24 日之后又逐渐升高。7 月11 日水稻单作与稻蟹共作组DO 值存在显著性差异(P<0.05)。8月5 日,水稻单作、稻蟹共作II 和稻蟹共作III DO值无显著性差异(P>0.05),但与稻蟹共作I 有显著性差异(P<0.05)。9 月2 日,稻蟹共作II 和稻蟹共作III DO 值无显著性差异(P>0.05),但与稻蟹共作I 均有显著性差异(P<0.05)。实验期间,水稻单作SPC 值较低,而稻蟹共作组SPC 值则基本稳定在0.3~0.4 ms/cm 左右。除8 月15 日水稻单作与稻蟹共作组SPC 值无显著性差异(P>0.05)外,其余各测量时间点水稻单作SPC 值均显著低于稻蟹共作组SPC 值(P<0.05)。7 月25 日和8 月24 日稻蟹共作组内SPC 值存在显著性差异(P<0.05)。
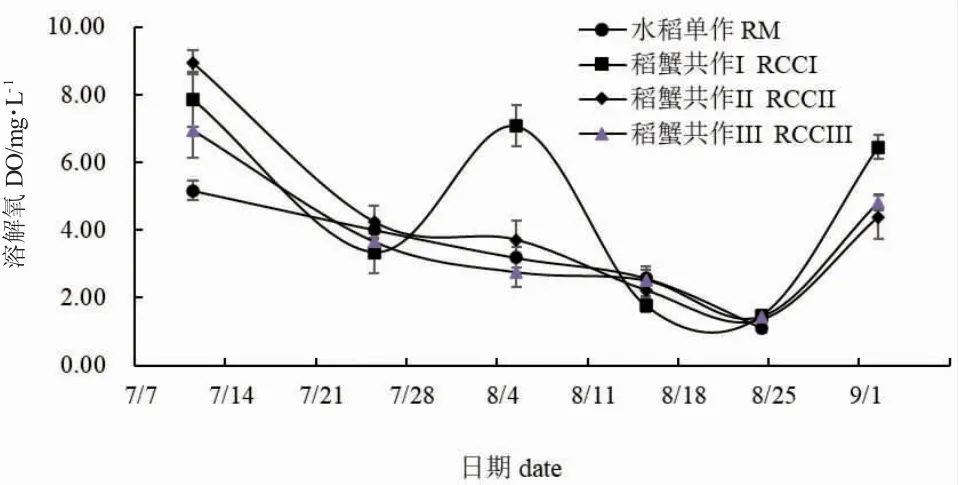
图4 各处理组稻田水体溶解氧量变化Fig.4 Changes in dissolved oxygen(DO)levels in paddy field water in each treatment

图5 各处理组稻田水体电导率变化Fig.5 Changes in paddy field water conductivity(SPC)in each treatment
实验期间,水稻单作和稻蟹共作组稻田水体TAN 数值均呈现先降低后升高再降低的趋势(图6),但出现的时间不同。7 月25 日,稻蟹共作II 与稻蟹共作III TAN 值无显著性差异(P>0.05),水稻单作与稻蟹共作I TAN 值无显著性差异(P>0.05),但水稻单作TAN 值显著高于稻蟹共作II(P<0.05)。8 月5 日,水稻单作TAN 值显著高于稻蟹共作组(P<0.05)。其余各采样时间,水稻单作与稻蟹共作组TAN 值间均无显著性差异(P>0.05)。图7 为水体NO2-N 数值变化情况。实验期间水稻单作水体NO2-N 值整体上呈下降趋势,而稻蟹共作组则波动较大。7 月11 日,水稻单作与稻蟹共作I NO2--N 值无显著性差异(P>0.05),但却显著高于稻蟹共作II、稻蟹共作III(P<0.05)。8 月5 日和8 月15 日,稻蟹共作III NO2--N 值显著高于其余各处理组(P<0.05)。
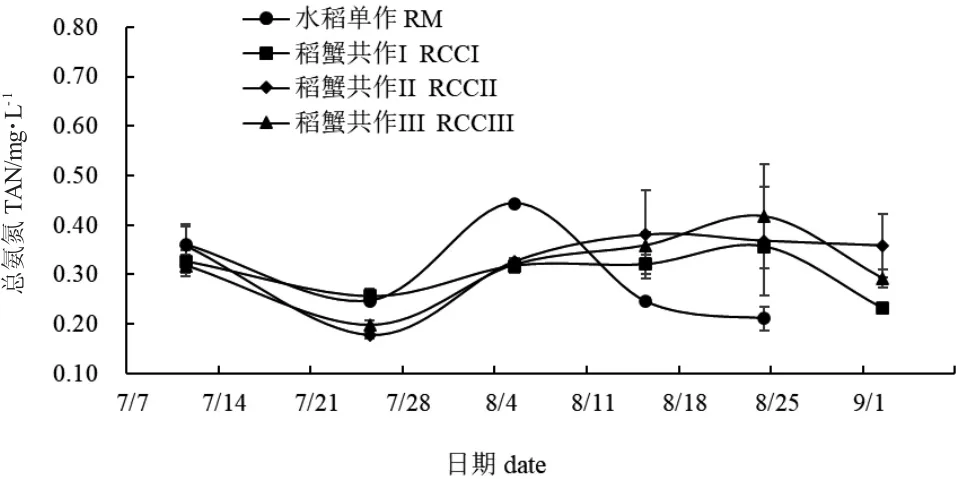
图6 各处理组稻田水体总氨氮变化Fig.6 Changes in total ammonia nitrogen(TAN)in paddy field water in each treatment
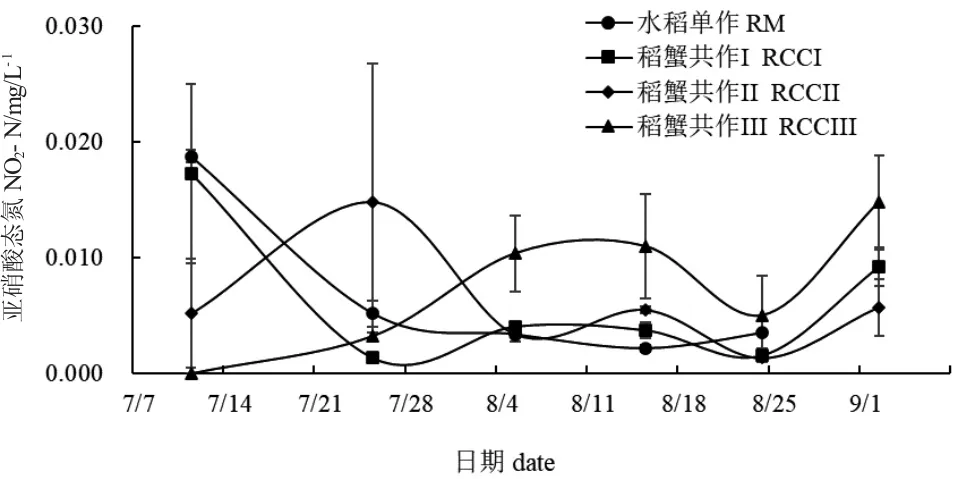
图7 各处理组稻田水体亚硝酸态氮变化Fig.7 Changes in nitrite nitrogen(NO2--N)in paddy field water in each treatment
实验期间各处理组稻田水体NO3--N 和TN 数值变化情况见图8 和图9。7 月11 日—7 月25 日,各组NO3-N 值均呈下降趋势,随后稳定在0~0.50 mg/L 之间。整个实验期间,稻蟹共作III NO3--N 值均显著高于其余各处理组(P<0.05)。整体来看,TN数值呈现波动变化。8 月5 日,水稻单作与稻蟹共作I TN 值无显著性变化(P>0.05),但与其他处理组差异显著(P<0.05)。8 月24 日,水稻单作与稻蟹共作组均存在显著性差异(P<0.05)。

图8 各处理组稻田水体硝酸态氮变化Fig.8 Changes in nitrate nitrogen(NO3--N)in paddy field water in each treatment
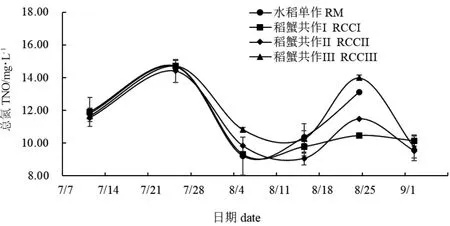
图9 各处理组稻田水体总氮变化Fig.9 Changes in total nitrogen(TN)in paddy field water in each treatment
图10 和图11 分别为各处理组稻田水体PO4-P和TP 数值变化趋势。整体来看,水稻单作PO4--P值呈现先升高后降低的趋势(图10),且7 月11 日到8 月24 日间采样点水稻单作与稻蟹共作组PO4--P 值均存在显著性差异(P<0.05)。9 月2 日,稻蟹共作组PO4--P 值均呈现下降趋势。整个养殖期间,除7 月11 日,水稻单作TP 值与稻蟹共作组数值无显著性差异(P>0.05)外,其余采样点水稻单作TP 值均显著低于稻蟹共作组(P<0.05)。9 月2 日,稻蟹共作组TP 值降低,稻蟹共作I 与稻蟹共作III无显著性差异(P>0.05),但与稻蟹共作II 均存在显著性差异(P<0.05)。
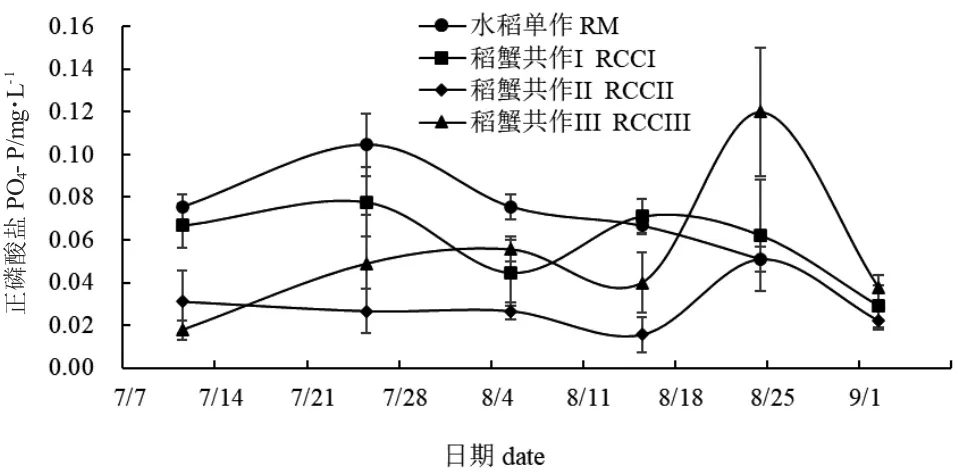
图10 各处理组稻田水体正磷酸盐变化Fig.10 Changes in orthophosphate(PO4-P)in paddy field water in each treatment

图11 各处理组稻田水体总磷变化Fig.11 Changes in total phosphorus(TP)in paddy field water in each treatment
整个实验期间,水稻单作COD 值呈先降低再升高最后下降的趋势(图12);随着扣蟹放养密度的增大,稻蟹共作组间COD 值差异较大。除8 月5日,各处理组COD 值无显著性差异(P>0.05)外,其余采样时间均存在显著性差异(P<0.05)。整体来看,各采样时间点水稻单作Chl.a 值均稳定在0.02 mg/L左右,低于稻蟹共作组。7 月25 日、8 月5 日和8 月24 日,水稻单作Chl.a 值显著低于稻蟹共作组(P<0.05)(图13)。
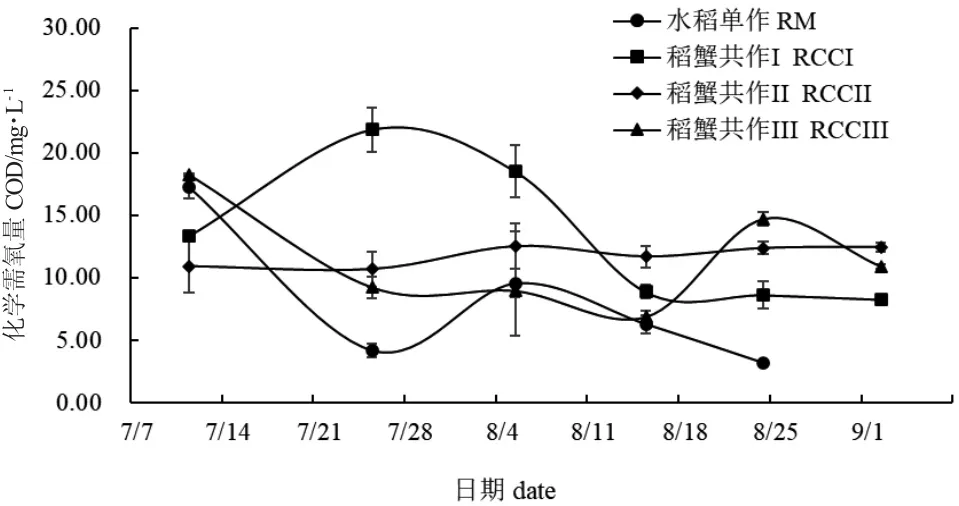
图12 各处理组稻田水体化学需氧量变化Fig.12 Changes in chemical oxygen demand(COD)in paddy field water in each treatment
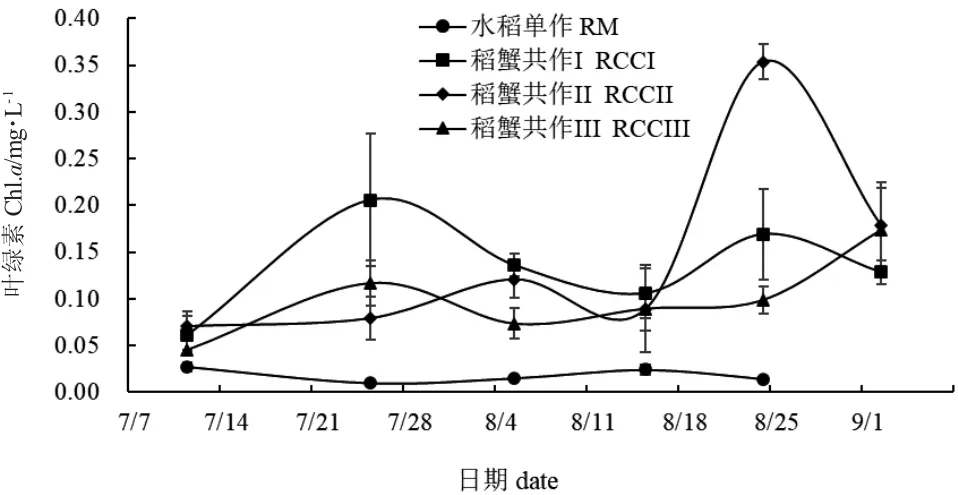
图13 各处理组稻田水体叶绿素a 变化Fig.13 Changes in Chl.a in paddy field water in each treatment
3 讨论
稻蟹共作是利用水稻与中华绒螯蟹互利共生使物质和能量向有利于水稻和中华绒螯蟹生长的方向流动,促进稻蟹共生生态系统内的物质和能量良性循环[23]。水稻田为中华绒螯蟹提供了优良的生存环境和食物来源及隐蔽场所;而中华绒螯蟹可以捕食杂草及害虫,排泄的粪便及未残余饲料可以为水稻施肥,促进水稻生长。本研究表明,稻蟹共作I中华绒螯蟹终末平均体质量为(16.02±0.58)g,明显高于辽宁盘锦地区稻蟹共作养成中华绒螯蟹平均体质量(11.97±0.15)g[7],这可能与养殖中华绒螯蟹的不同地理种群有关。本实验采用的是长江水系中华绒螯蟹,而盘锦地区所采用的是辽河水系中华绒螯蟹。辽宁盘锦地区稻田的养殖环境与黑龙江高寒地区稻田养殖环境也有所差异,以上两方面都可能造成中华绒螯蟹生长的差异。本实验扣蟹终末体质量、WGR 和SGR 均明显高于长江流域池塘养殖扣蟹[24],可能与初始苗种体质量、苗种质量以及养殖环境、幼蟹放养密度等有关。本研究表明:水稻单作产量低于稻蟹共作,这与张云杰等[7]和孙文通等[8]的研究结果一致。首先稻蟹共作组采用大垄双行的水稻栽培方式,具有边际效应,可以提高水稻产量[25],其次稻蟹共作组中华绒螯蟹的觅食活动可以翻动土壤,改良土壤结构[26],粪便中含有丰富的可降解有机碳,有利于微生物增殖,促进土壤活性改善,增加土壤养分,促进水稻生长[8]。稻蟹共作III的水稻产量最高,说明在一定放养密度条件下,水稻产量随着放养密度的提高而增加。
稻田水质理化因子是稻蟹共作生态系统的重要环境因素,环境中水质的质量显著影响水稻、中华绒螯蟹以及水体中浮游生物的生长[7]。稻田水体浅,水环境因子不稳定,探究稻蟹共作期间水环境因子的变化,可以为水稻、中华绒螯蟹生长提供重要的科学依据。温度是影响中华绒螯蟹生长发育的重要因子,温度过高(≥30℃)对中华绒螯蟹生长不利,中华绒螯蟹生长的最适温度为20~26℃。温度影响中华绒螯蟹生长发育主要体现在摄食、1 龄性早熟、生长蜕壳和性腺发育等方面[8]。整体来看,水稻单作TEMP 略高于稻蟹共作,但无显著性差异(P>0.05)。7~8 月间,稻田TEMP 呈现先下降再上升的趋势,主要与2019 年黑龙江省7~8 月间持续降水有关。DO 是中华绒螯蟹养殖中另一重要水环境因子[27],其来源主要是空气中氧气溶解和水体中植物光合作用。影响稻田水体溶解氧的因素主要有:光照强度、水温、空气中氧分压和水体中浮游植物数量等。本研究表明,稻蟹共作组水体DO 含量高于水稻单作,可能有以下三个原因。首先是稻蟹共作组“大垄双行,边沟加密”的栽培模式有利于水体接收更多的光照;其次,稻蟹共作组水体中Chl.a 含量显著高于水稻单作组,说明稻蟹共作组水体中浮游植物数量显著高于水稻单作,故其光合作用总量更大,释放出氧气量更高,因此水体中DO 含量更高。第三,中华绒螯蟹在稻田中觅食扰动水体,也可以增加一部分空气中氧气溶解在水体中。在稻蟹共作组,随着中华绒螯蟹放养密度的增加,水体中DO含量略下降,这可能与中华绒螯蟹密度高,呼吸作用总耗氧量高有关。本研究中7~8 月,稻蟹共作模式的pH 均呈弱酸性,且数值逐渐降低,可能与此期间持续降水有关,导致水体光照不足,水体中浮游植物和中华绒螯蟹呼吸作用占主导,产生CO2溶解于水体中,导致水体pH 降低。SPC 是表征水体溶解性固体物质或盐度的重要参数,可以直接反应水体中可溶性离子总量[28],并与水体营养物质浓度和悬浮物含量呈一定的正相关性[29]。本研究表明,稻蟹共作组SPC 数值显著高于水稻单作(P<0.05),说明中华绒螯蟹对水稻田泥土的扰动,加速了泥土中可溶性离子溶解到水体中。这与在稻鸭共作[30]和稻虾共作[31]的研究结果一致。
水体中总氮包括有机态氮和无机态氮两大类,有机态氮主要是氨基酸、蛋白质、核酸和腐殖酸等物质中所含的氮;无机态氮主要有AN、NO2--N 和NO3--N[32]。稻田水体AN 多来自含氮有机物分解还原以及水生动物自身代谢[33]。NO2--N 是水体中含氮有机物分解的中间产物,氧气充足时可以转化为NO3--N,而氧气不足时则可以转化为毒性较强的AN[8]。水体中NO3-N 主要由施肥和NO2--N转化而来,可以被水体中的浮游植物和水稻直接吸收。其中AN 和NO2--N含量是影响甲壳动物生长、免疫以及生理机能的主要限制因素之一[34-36]。实验期间,稻蟹共作组水体TAN、NO2--N、NO3--N和TN 值与水稻单作无显著性差异(P>0.05),说明稻蟹共作并未显著影响稻田水体含氮物质的指标。稻田水体NO3--N含量可以间接反映水稻田的氮肥供应能力。7 月11 日—7 月25 日期间,各组NO3--N值均呈现下降趋势,主要是由于此期间水稻对氮素的需求量大,因此NO3-N 呈较快的下降趋势,这与已往研究文献结果一致[22,37]。整体来看,稻蟹共作组随着中华绒螯蟹养殖密度的增加,NO3--N含量呈现递增的趋势,说明中华绒螯蟹放养密度增大,硝化细菌数量增多,因此将NO2--N转化为NO3--N的能力增强,故NO3--N含量逐渐增多。稻蟹共作组中土壤TN 含量显著高于水稻单作组(P<0.05),可能与中华绒螯蟹活动能有效促进水稻吸收土壤氮有关。
磷肥是植物生长发育过程中不可或缺的肥料,其重要性不亚于氮。磷是植物中核酸和蛋白质的主要组成部分,在物质的形成、代谢及运转等方面都起着非常重要的作用[38]。PO4--P 含量可以间接反映水稻田磷肥供应能力,而TP 则可以反映水体P 元素物质的含量。本研究表明,水稻单作PO4--P 含量显著高于稻蟹共作组(P<0.05),这与水稻单作Chl.a含量显著低于稻蟹共作组一致。说明稻蟹共作组浮游植物量增加,消耗PO4--P 量增加,故而稻蟹共作组PO4--P 含量低于水稻单作。稻蟹共作组TP 含量显著高于水稻单作(P<0.05),说明稻蟹共作可以提高水体TP 的含量,这与稻鸭共作的研究结果一致[30],稻蟹共作组TP 含量高与中华绒螯蟹在稻田中的觅食扰动、粪便排泄以及饲料残饵分解等有关。COD 是表征环境水样中有机物污染的特定综合指标,反映水环境中有机物的含量,且其主要来源为养殖过程中的代谢产物和残饵累积[39]。本研究表明,稻蟹共作组COD 含量高于水稻单作,说明中华绒螯蟹的扰动以及饲料残饵的分解等使水体中有机物的含量升高。
综上所述,本文系统研究了北方寒区稻-扣蟹共作模式下,中华绒螯蟹和水稻的生长性能,以及在整个稻蟹综合种养周期内稻田水质理化因子的变化规律。结果表明,在高纬度寒冷地区稻蟹综合种养过程中,设置适宜的蟹苗放养密度可培育出大规格扣蟹,可显著增加水稻的产量,与水稻单作相比,该模式除稻田水体SPC、TP 和叶绿素a 含量较高之外,其他水质参数并未表现出显著变化。这将为后续稻蟹综合种养过程中大规格商品蟹的养殖奠定基础。本研究结果也为稻-扣蟹共作模式在高寒地区的推广提供依据。
猜你喜欢
新农民(2023年18期)2023-07-06 08:27:20
广西林业科学(2022年2期)2022-05-09 07:49:54
天津农林科技(2022年2期)2022-04-19 10:48:16
今日农业(2021年10期)2021-11-27 09:45:24
今日农业(2021年17期)2021-11-26 23:38:44
科教创新与实践(2021年38期)2021-11-14 07:09:39
今日农业(2020年16期)2020-12-14 15:04:59
江西畜牧兽医杂志(2020年4期)2020-09-12 06:36:26
山东农业科学(2019年11期)2019-12-24 01:11:27
江苏农业科学(2019年11期)2019-07-22 01:32:45
