
银腺杨Pag WOX11/12a基因对茎生长发育的影响
2023-04-29 00:44文爽爽王留强卢孟柱
林业科学研究 2023年1期
文爽爽 王留强 卢孟柱
关键词:PagWOX11/12a;银腺杨;节间伸长;株高;功能分析
中图分类号:S718.46 文献标志码:A 文章编号:1001-1498(2023)01-0039-08
WUSCHEL-related homeobox(WOX)家族是真核生物中Homeobox(HB)转录因子超家族的成员,该家族成员均含有同源异型结构(Homeodomain, HD),且为植物特异性转录因子。拟南芥(Arabidopsis thaliana (L.) Heynh.)WOX家族共有15个成员,这些成员可分成3个分支:现代/WUS型分支(WUS、WOX1-7)、中间型分支(WOX8-9、WOX11-12)和古老型分支(WOX10、WOX13-14)。除WOX7外,现代/WUS型分支的成员中均含有1个特定的WUS-box结构域,该结构域在中间型分支及远古型分支成员中并不完整。
WOX基因家族的成员参与调控了植物体不同的生长发育过程,它们在调节细胞增殖和分化方面发挥着重要功能,同时也参与了植物顶端分生组织和干细胞的维持、侧生器官和花器官的形成、胚胎发育、激素信号转导和逆境响应等过程。拟南芥WUS蛋白可以通过和SHOOT MERISTEMLESS(STM)的相互作用,增强WUS对CLAVATA3(CLV3)基因启动子的结合能力,二者共同调控CLV3的表达,维持干细胞稳态。同时,AtWOX1基因也可以调控CLV3基因的表达,且能通过调节脱羧酶蛋白(Decarboxylase,SAMDC)活性以及多胺的稳态,参与拟南芥分生组织的发育。AtWOX4基因可以促进原形成层的发育,调节植物的侧向生长。毛白杨Pto WOX5a基因可以调控杨树不定根的发育,过表达该基因能够增加转基因杨树不定根的数量。过表达WOX14可以促进拟南芥体内赤霉素的积累,促进花序茎中的维管细胞分化及木质化。过表达毛白杨WOX11/12a基因可以促进形成层的发育,影响杨树木质部分化。
杨树(Populus)是重要的速生造林树种,同时也是木本植物分子生物学研究的模式植物。已有报道指出,WOX11/12基因可以调控植物的根系发育和茎部次生生长,并能参与植物对非生物胁迫的响应,这表明WOX11/12基因在植物生长发育中起着重要作用。为明确WOX11/12基因对杨树其它组织及器官生长发育的作用,本研究以非转基因银腺杨(Populus alba×P.glandulosa,即‘84K)和PagWOX11/12a显性抑制转基因杨树(35S::Pag WOX11/12a-SRDX)为材料,研究了显性抑制该基因后杨树高生长及茎中细胞长度的变化趋势,揭示了Pag WOX11/12a基因对杨树茎部生长发育的影响,为进一步探讨其作用机制提供参考。
1材料与方法
1.1研究材料
本研究以84K杨和35S::Pag WOX11/12a-SRDX(DR)转基因杨树为研究材料,35S::Pag WOX11/12a-SRDX是由编码SRDX肽的核酸序列与Pag WOX11/12a的cDNA融合构建的载体转化84K杨所得到的转基因苗,该载体可以特异性抑制目标基因的表达,该转基因苗由前期研究获得,组培苗保存于中国林业科学研究院林木遗传育种国家重点实验室。上述组培苗在生根培养基中培养,培养基组成为:1/2 MS(Murashige& Skoog Basal Medium w/Vitamins)基础培养基、0.05 mg·L-13-吲哚丁酸(3-indole butyric acid,IBA)、0.02 mg·L-1萘乙酸(Naphthylacetic acid,NAA)及5 mg·L-1琼脂粉(pH=5.8~6.0)。组培苗培养于人工气候室(温度:24±1℃;光周期:16 h/8 h光照,黑暗;相对湿度:50%~60%:光照强度:55~65μmol·m-2·s-1)。待生长21 d后,将组培苗转入营养土(土壤:珍珠岩= 3:1)中,并置于温室培养。
本研究中定量试剂盒SYBR PremixExTaqTMKit及反转录试剂盒PrimeScriptTMRT reagent Kit购自TaKaRa,RNA提取试剂盒(RNAprep pure Plant Kit)购自QIAGEN。
1.2研究方法
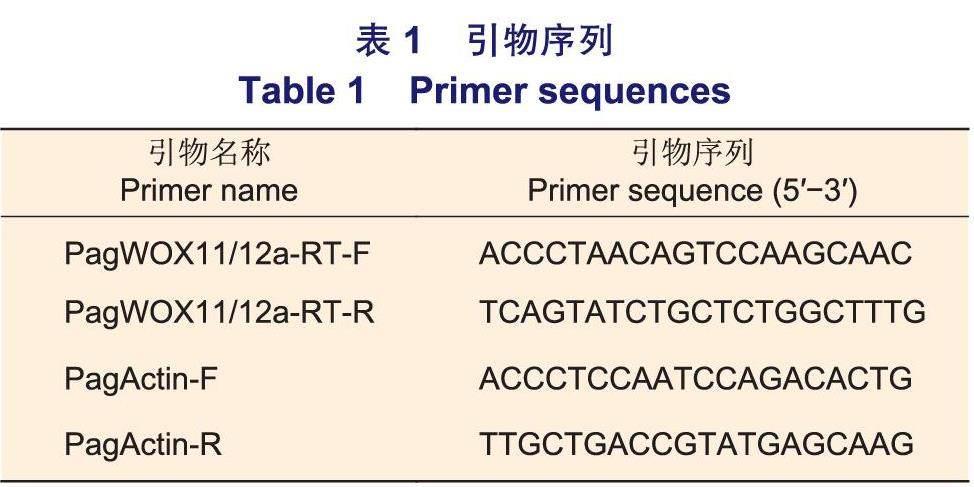
1.2.1生物信息学分析 使用在线网站ExPASy(https://web.expasy.orglprotparaml)分析PagWOX11/12a蛋白的理化性质。利用植物基因组网站(https://phytozome.jgi.doe.gov/pz/portal.html)获得毛果杨(Populus trichocarpa Torr.&Gray) WOX基因家族的氨基酸序列。84K杨基因组获取于Figshare database( https://doi.org/10.6084/m9.figsh are.12369209),利用本地BLAST获得84K杨中WOX基因家族的蛋白序列。使用MEGA 11软件,根据邻接法(Neighbor-joiningmethod)构建系统发育进化树,节点旁的数字表示基于1000次重复次数的自展值(Bootstrap)。
1.2.2基因组织表达特异性分析 利用RNA提取试剂盒提取生长60 d的84K杨不同组织的总RNA,包括茎部木质部、韧皮部及形成层、成熟叶、1/2株高处茎部、主根和侧根,提取方法参照说明书。分别用Nanodrop8000和1.5%琼脂糖凝胶电泳检测RNA的浓度及质量。利用反转录试剂盒合成cDNA,反应体系及条件参照说明书。通过在线网站(https://sg.idtdna.com/Scitools/Applications/RealTimePCR/)设计PagWOX71/72a的定量引物PagWOX11/12a-RT-F和PagWOX11/12a-RT-R(表1)。利用SYBR Premix Ex TaqTMKit(TaKaRa)定量试剂及实时荧光PCR仪器Light Cycler 480(Roche Applied Science, Penzberg,Germany)进行qRT-PCR分析。选用PagActin作为内参基因,且进行3次生物学重复和3次技术重复,相对表达量根据2-△△CT方法进行计算。
1.2.3转基因植株表型鉴定 分别选取84K、DR1和DR9各9株长势一致的组培苗移入营养土中,并将其置于温室培养。拍照记录其表型变化,测量植株在培养第7、14、21、28天后的株高及节间数,统计第28天时前9节间的节间长度。上述实验均进行3次生物学重复。
1.2.4茎段解剖结构分析 选取生长28 d的植株第9节间茎段进行解剖结构分析。于第9节间茎段中部取约0.5~1 cm的材料,使用5%琼脂糖包埋后,将其水平放置固定于样品架上,通过震荡切片机(LEICA VT1000)进行纵切,切片厚度设置为40 pm。将切片置于去离子水中等待后续观察。观察茎段解剖结构时使用0.05%甲苯胺蓝O(Toluidine blue O,TBO)进行染色,待染色结束后用去离子水对样品进行冲洗;随后,用OlympusBX51显微镜进行观察并及时拍照,利用ImageJ软件测量照片中髓心及韧皮部细胞的长度。
1.2.5纤维离析 将生长28 d的植株相同节间茎段切成1 cm长度,去除树皮,并将茎段置于离析液(30%过氧化氢:去离子水:冰乙酸=1:4:5)中,样品至少在56℃金属浴中放置72 h;随后用去离子水将样品洗涤3~5次,并浸泡过夜;最后将样品捣碎,使用TBO进行染色并吸取适量混悬液至载玻片中,置于Olympus BX51显微镜下观察、拍照,通过Image J软件测量木质部纤维细胞的长度。
1.2.6数据统计 使用软件IBM SPSS Statistics23对统计的数据进行分析,使用f检验法分析差异显著水平。
2结果与分析
2.1 84K杨WOX基因家族的鉴定及系统发育分析
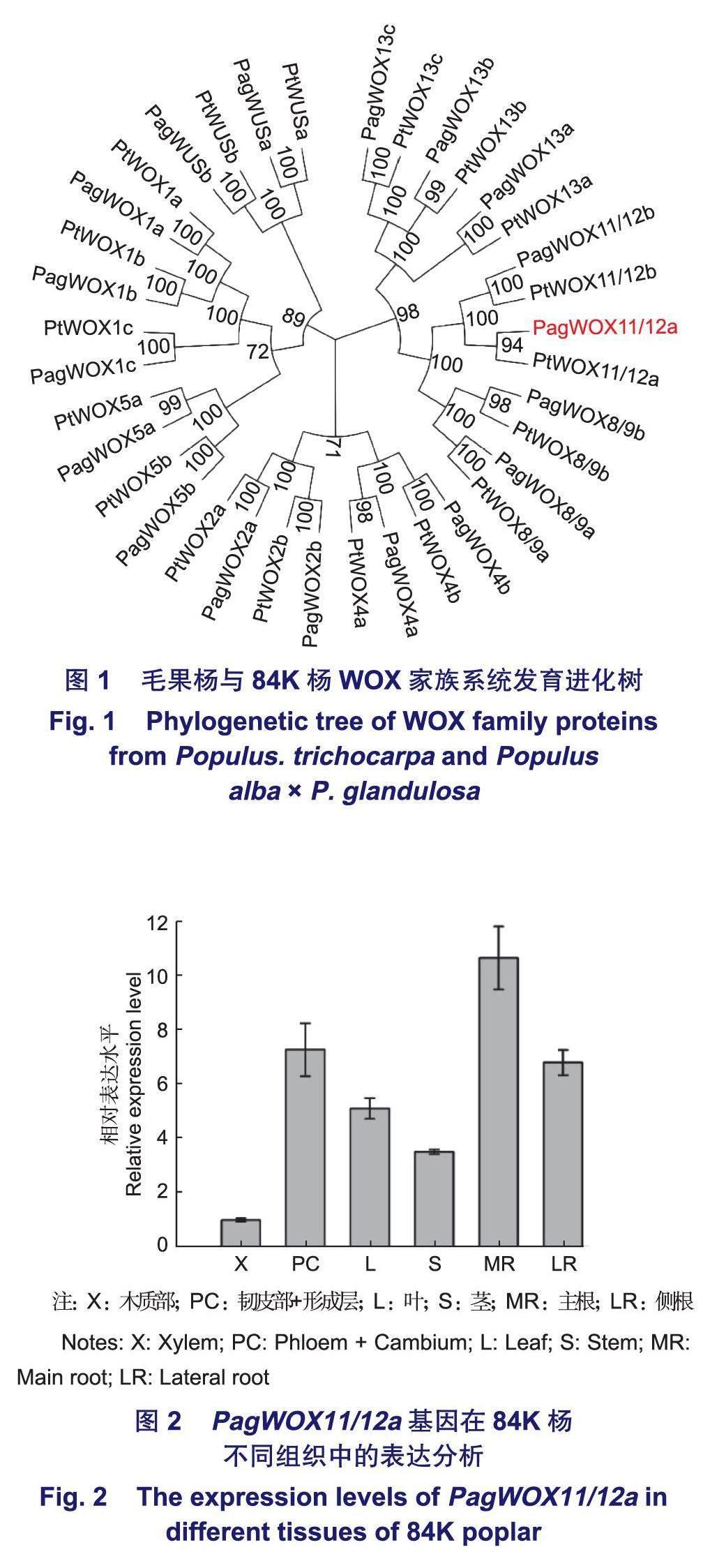
毛果杨PtWOX基因家族共有18个成员。利用从Phytozome网站获取的Pt WOX基因序列在84K杨基因组数据库中进行BLAST分析,最后在84K杨中鉴定出18个PagWOX基因家族成员。基于PtWOX和PagWOX的蛋白序列,利用邻接法构建了毛果杨和84K杨WOX家族的系统发育进化树(图1)。根据构建好的系统发育进化树,对84K杨PagWOX进行命名。
2.2 Pag WOX11/12a序列分析
对Pag WOX11/12a基因的核酸及蛋白序列进行分析。结果表明:84K杨与毛果杨WOX11/12a基因的核酸序列相似度为96.88%,蛋白序列相似度为95.29%。Pag WOX11/12a基因编码区长度为768 bp,编码的蛋白序列包含255个氨基酸,该蛋白的分子量约为28.18 kDa,等电点为5.30,脂肪系数为67.22,不稳定系数为62.31,蛋白平均亲水性(GRAVY)为-0.461。
2.3 Pag WOX11/12a基因组织表达特异性分析
为探究Pag WOX11/12a基因在84K杨不同组织中的表达情况,分别提取生长60 d的84K杨的木质部、韧皮部及形成层、成熟叶、1/2株高处茎部节间、主根和侧根的总RNA,反转录后对各部位进行qRT-PCR检测。结果表明:PagWOX11/12a基因在各个部位中均有不同程度的表达,在主根、侧根及韧皮部和形成层中表达量较高,在叶片及茎段中也有一定的表达量,在木质部中表达量最低(图2)。
2.4转基因植株表型分析
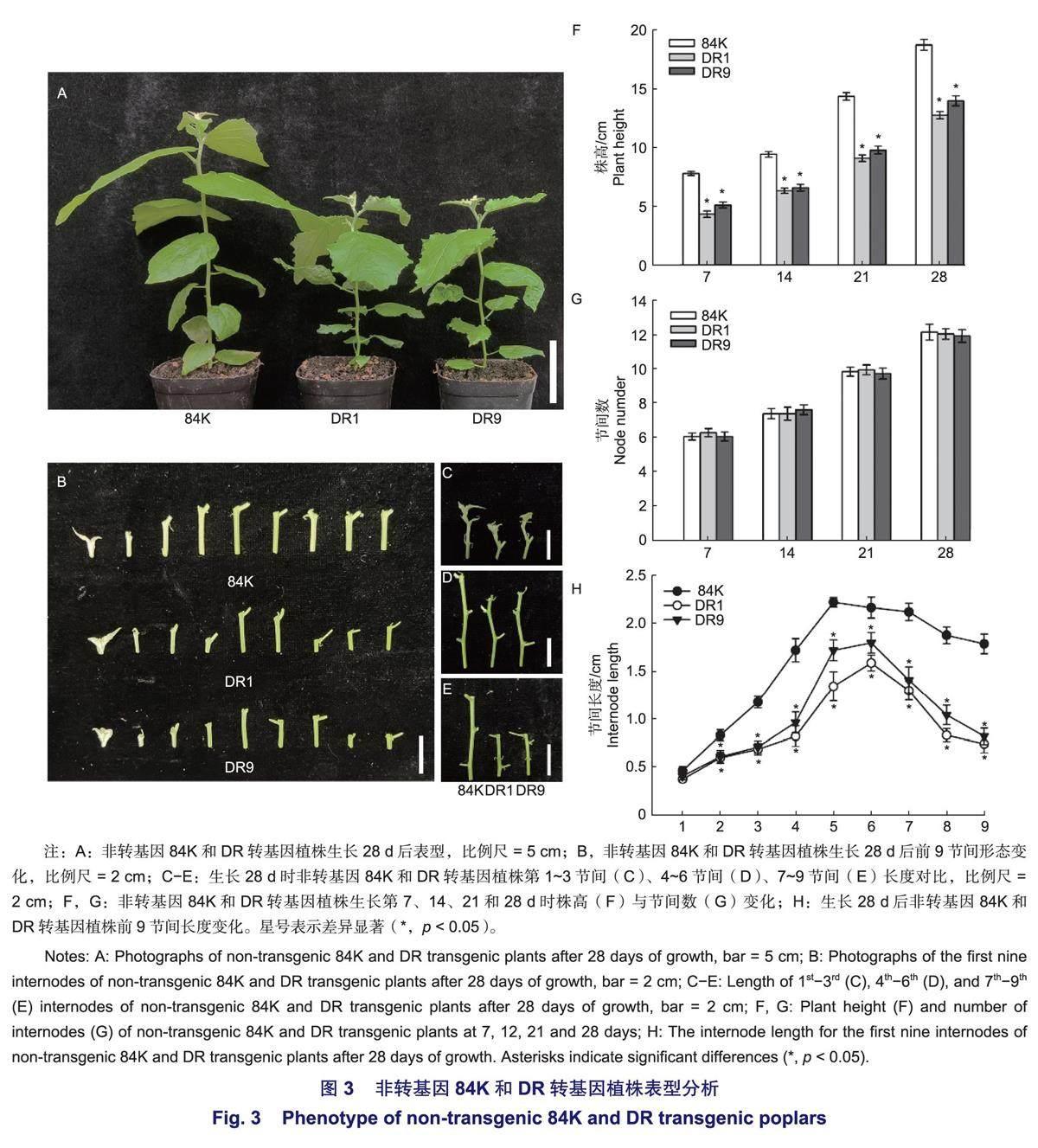
为了明确Pag WOX11/12a基因对杨树生长发育的影响,分别统计了非转基因84K和DR转基因植株在温室生长7、14、21和28 d后的表型(图3)。通过统计不同株系的株高与节间数发现,DR转基因植株的株高在第7、14、21、28天时均显著小于非转基因84K,而二者的节间数则无明显差异(图3A,F~G)。同时比较了非转基因84K和DR转基因植株在第28天时前9节间的表型(图3B)以及第1~3、4~6、7~9节间的变化(图3 C~E),可见DR转基因植株的节间长度小于对应84K的节间长度。为进一步明确各株系节间长度的变化,测量了第28天时非转基因84K和DR转基因植株前9节间的长度(图3H)。结果表明:从第2节间开始,DR转基因植株的节间长度均显著小于非转基因84K。上述结果表明,显性抑制Pag WOX11/12a基因抑制了杨树节间的伸长。
为进一步明确Pag WOX11/12a基因对杨树茎部生长发育的影响,取生长28 d的非转基因84K和DR转基因植株第9节间茎段,通过纵切切片观察髓心和韧皮部细胞的结构特征,并统计其长度(图4)。结果表明:DR转基因植株的韧皮部细胞及髓心细胞与非转基因84K相比,长度均有不同程度的减小(图4A~C)。DR1和DR9的韧皮部细胞及髓心细胞长度显著小于非转基因84K。与84K相比,DR转基因株系DR1和DR9的韧皮部细胞分别减小了30%与21%,髓心细胞长度分别缩短了25%及15%(图4D、E)。
除了韧皮部及髓心外,木质部也是杨树茎段的重要组成,利用木质部纤维离析对非转基因84K和DR转基因植株的木质部纤维细胞长度进行了分析(图5)。通过对木质部纤维细胞的长度统计(图5D)可知:DR1和DR9的木纤维细胞与非转基因84K相比,分别减小了26%与20%。上述结果表明,DR转基因植株节间缩短是由茎中细胞长度减小导致。
3讨论
杨树是我国重要的速生造林树种,具有繁殖容易、生长快等特性,是生产建筑、造纸工业及生物燃料生产的重要材料。同时其转基因体系相对成熟,也是最早完成全基因组测定的树种,是木本植物进行分子生物学研究的模式树种。研究参与调控杨树生长发育相关基因,对于杨树的分子育种具有重要意义。本研究选取了与分生组织相关的WOX家族基因,以期明确Pag WOX11/12a基因对杨树高生长发育的作用,进而提高木材产量。
已有研究表明,毛果杨PtWOX基因家族共有18个成员,本研究利用84K杨的基因组信息,通过序列比对在84K中找到了18个同源序列,并构建了系统发育进化树(图1)。通过序列分析发现,PagWOX11/12a基因与毛果杨PtWOX11/12a基因序列相似度较高。
Pag WOX11/12a基因的组织表达特异性分析结果(图2)表明,该基因在主根中的表达量最高,在侧根中表达量也相对较高,这表明PagWOX11/12a基因可能影响杨树根系发育。已有研究表明,在杨树中过表达WOX11/12a可以促进植物根系发育。同时该基因在韧皮部及形成层中也有较高的表达量,暗示该基因很有可能参与韧皮部及分生组织发育的调控。
植株的高度通常由节间数量及节间长度决定。本研究对非转基因84K及DR转基因植株进行表型分析(图3),发现在不同时期内,二者的节间数量均无明显差异,而DR转基因植株的株高则显著低于非转基因84K,表明DR转基因植株的节间长度小于非转基因84K。为明确PagWOX11/72a基因对节间长度的影响,观察并统计了在温室生长28d后非转基因84K及DR转基因植株的节间长度(图3B~E,H),结果显示从第2节间开始,DR转基因植株的节间长度均低于非转基因84K,且差异显著。由此可知,PagWOX11/12a基因可以通过改变节间伸长来影响植株高度,节间长度的降低是DR转基因植株矮化的主要原因。为更精准的分析Pag WOX11/12a基因对杨树高生长的影响,对杨树茎段进行了解剖学分析,通过观察对比非转基因84K与DR转基因植株茎段纵切切片(图4),发现DR转基因杨树茎中韧皮部细胞及髓心细胞长度小于非转基因84K,同时纤维离析结果(图5)表明,与非转基因84K相比,DR转基因植株的纤维细胞的长度也显著变短。上述结果进一步说明,DR转基因植株节间长度的降低是由茎中细胞长度缩短所致,PagWOX11/12a基因可以通过影响杨树茎中细胞长度来控制节间伸长,进而改变植株高度。已有研究表明,毛白杨PtoWOX11/12a基因可以影响杨树茎的发育,改变转基因植株株高及地径,过表达毛白杨PtoWOX11/12a基因后可以促进形成层细胞层数的增加,但会抑制木质部的发育,表明Pto WOX11/12a基因可以影响形成层干细胞的活动及木质部的分化;而84K杨PagWOX11/12a基因可以通过调控韧皮部、髓心细胞及木质部纤维细胞的伸长,影响植株的节间长度,说明了WOX11/12a基因对茎生长发育的调控途径。前期已有研究表明,PagWOX11/12a基因可以促进根系生长,增强植物的抗旱性及耐盐性,而本研究指出,该基因也可以调控杨树茎的生长发育,进一步明确了PagWOX11/12a基因对杨树生长发育新的调控途径。
通过了解Pag WOX11/12a基因对节间伸长的调控作用有助于提高木材产量。已有研究表明,茎段节间伸长主要由生长素(Auxin)、赤霉素(Gibberellins,GAs)、油菜素内酯(Brassinosteroids,BRs)和独角金内酯(Strigolactones,SLs)等激素控制。虽然上述激素在茎段节间伸长中发挥的作用不同,但它们的生物合成或信号转导过程均可能影响植株高度。后续可以通过测定激素含量进一步明确PagWOX11/12a基因影响节间伸长的途径。
4结论
株高是林木遗传育种中的重要性状之一,本研究明确了PagWOX11/12a基因可以通过影响节间伸长改变植株高度,通过统计韧皮部细胞、髓心细胞及木质部纤维细胞的长度可以得知,该基因对节间伸长的抑制主要是通过控制茎中细胞长度来实现的,上述结果对揭示PagWOX11/12a基因对杨树的生长调控机制具有重要意义。
猜你喜欢
中国果业信息(2023年12期)2024-01-09
湖北农业科学(2021年13期)2021-08-04
植物研究(2021年2期)2021-02-26
World Journal of Clinical Cases(2020年9期)2020-05-13
种子(2019年4期)2019-05-28
天津农业科学(2015年11期)2015-12-03
江苏农业科学(2015年4期)2015-06-15
热带作物学报(2014年3期)2014-04-29
安徽农学通报(2014年7期)2014-04-29
中国果业信息(2014年11期)2014-01-23
