
外源色氨酸对低氮胁迫下高粱苗期叶片碳氮平衡和衰老特性的影响
2023-04-10 07:38:54谷闻东刘春娟李邦刘畅周宇飞
中国农业科学 2023年7期
谷闻东,刘春娟,李邦,刘畅,周宇飞
外源色氨酸对低氮胁迫下高粱苗期叶片碳氮平衡和衰老特性的影响
谷闻东,刘春娟,李邦,刘畅,周宇飞
沈阳农业大学农学院,沈阳 110866
【】研究外源色氨酸对低氮胁迫下高粱叶片衰老的影响,探讨低氮条件下高粱叶片碳氮平衡与衰老的关系,旨在挖掘高粱耐低氮胁迫的有效调控手段。【】选用耐低氮高粱自交系398B和氮敏感自交系CS3541为试验材料,采用水培试验,设置正常氮(5 mmol·L-1)和低氮(0.5 mmol·L-1)2个氮水平,并进行50 mg·L-1外源色氨酸喷施处理,喷施10 d后测定叶片形态、组织结构、光合特性、叶绿素荧光参数、叶片碳氮代谢相关物质含量和酶活性、C/N及衰老相关基因表达水平,并分析低氮胁迫下高粱幼苗叶片C/N与衰老相关基因的相关性。【】(1)与正常氮水平相比,低氮胁迫显著降低了398B和CS3541的叶面积,而外源色氨酸处理使398B和CS3541的叶面积显著增加了36.72%和52.06%;外源色氨酸显著增加了低氮胁迫下398B和CS3541的叶干重和叶鲜重。(2)与正常氮水平相比,低氮胁迫下,398B花环结构相对完整,而外源色氨酸处理使叶片细胞排列整齐,花环结构清晰;外源色氨酸显著增加了低氮胁迫下398B叶片叶绿素含量(36.85%),而未显著提高CS3541叶片各色素含量。(3)低氮胁迫下,与未喷施色氨酸相比,外源色氨酸处理使叶片光系统II(PSII)表现出更高的最大光化学效率(v/m)和热耗散(NPQ),增加了叶片光合速率,从而维持叶片更强的光合能力。(4)外源色氨酸提高了低氮胁迫下高粱叶片氮含量和碳氮代谢相关酶活性;降低了高粱叶片碳水平(可溶性糖、蔗糖、淀粉和总糖)和C/N,进而保证低氮胁迫下植株的碳氮生理平衡。(5)外源色氨酸正调控低氮胁迫下衰老相关基因和的表达,负调控、、和的表达;另外,C/N与和表达量呈正相关,与、、和表达量呈负相关。【】低氮胁迫下,外源色氨酸通过降低C/N和衰老基因的表达量,影响叶片形态和光合特性,同时通过调节叶片碳氮代谢,延缓了叶片衰老,增强高粱幼苗对低氮胁迫的耐性。因此,喷施外源色氨酸是缓解低氮胁迫的一种潜在策略。
高粱;衰老相关基因;C/N;光合特性;碳氮代谢
0 引言
【研究意义】高粱(L.)常种植于边际土地,容易受到土壤养分供应不足的影响,氮素缺乏是影响高粱生长发育的重要因素,因此,阐明低氮胁迫对高粱造成的生理损伤,揭示增强其耐低氮胁迫能力的有效途径,是促进高粱生产健康发展的重要目标。【前人研究进展】氮(N)是植物体内的大量元素之一,对植物的生长发育和生理代谢具有重要作用[1]。低氮条件下,植物叶片形态、光合能力、碳水化合物代谢、蛋白质合成与降解等生理过程都会受到不同程度的影响[2]。一方面,叶片中约70% N会被分配到叶绿体中,其中大部分用于光合作用,因此,低氮胁迫会直接抑制叶片的光合特性[3];另一方面,低氮诱导叶绿素降解的同时,可以引起氮代谢受阻,蛋白质合成能力下降,进而影响植物的生命活动[4]。Liu等[5]研究表明,耐低氮糜子自交系通过维持较高的叶片叶绿素含量、光合能力和氮代谢相关酶活性,减弱了光抑制的损伤,因而具有更强的耐低氮胁迫能力。同时,氮素的缺乏也会引起植物叶片中糖分的积累,减少其转化分解,致使可溶性糖和淀粉含量升高,加速了叶片的衰老[6]。叶片是植物进行生命活动的主要器官,其衰老受植株内部因素(如发育阶段)和外部环境(如胁迫)共同调控,典型症状为形态上的黄化与生理活性的下降[7]。碳氮代谢作为维持叶片生理活性的关键过程,两者的生理平衡调控叶片衰老,而非碳或氮代谢单独发挥决定作用[8-9]。曹蓓蓓等[10]研究表明,碳氮平衡参与了低氮诱导的小麦叶片衰老,低氮条件下叶片光合能力下降,碳氮同化能力明显减弱。周琴等[11]指出,黑麦草叶片内氮素的匮乏和转移引起了叶绿素降解,导致碳同化能力降低,碳氮代谢平衡被打破,最终引起叶片衰老。拟南芥的全基因组表达分析也证实了碳氮信号在调控植物衰老中的重要性[12]。由此可知,维持碳氮平衡、延缓叶片衰老是植物适应低氮环境的一种重要策略。色氨酸(Tryptophan,Trp)由莽草酸途径的中间体分支酸经过一系列酶催化而成。色氨酸(C11H12N2O2)含有13%—14%的N,可通过转氨基作用、脱氨基作用及其他过程加以同化,被植物所吸收利用[13-14]。喷施外源色氨酸可使玉米和小麦氮吸收增加28%和31%[15-16],其代谢过程中N的释放可能是作物对色氨酸产生积极反应的关键原因。El-Awadi等[17]研究发现,叶片喷施外源色氨酸降低了菜豆对氮肥的需求,对植株生长发育具有积极的作用。此外,色氨酸通过维持植株体内离子与激素的平衡,增强细胞膜的稳定性,一定程度上提高了植株对非生物胁迫的耐受性[18]。【本研究切入点】近年来,随着中国农业供给侧结构性改革的推进,高粱以其优异的抗逆性和营养品质成为种植业结构调整中重要的替代作物[19]。长期以来,作物对低氮胁迫的响应及提高耐低氮能力的研究大多集中在水稻[20]、小麦[21]和玉米[22]等主要粮食作物上,有关低氮胁迫下高粱生理代谢适应能力,以及外源色氨酸提高耐低氮生理响应机制的研究却鲜见报道。【拟解决的关键问题】本研究以耐低氮和氮敏感高粱自交系为材料,探讨外源色氨酸作用下高粱叶片形态-微观结构-光合特性-生理生化水平的变化规律,从碳氮平衡角度揭示外源色氨酸缓解低氮胁迫下高粱叶片衰老的生理机制,以期为中国高粱生产的可持续发展提供理论依据。
1 材料与方法
1.1 试验材料和设计
选用耐低氮自交系(398B)与氮敏感自交系(CS3541)为试验材料,在人工气候箱内进行水培培养。挑选大小一致,籽粒饱满的高粱种子,用10% NaClO溶液消毒5—10 min,蒸馏水洗净后摆放于铺有湿润滤纸的培养皿中,置于培养箱中培养,培养箱条件设置为:昼夜温度为28℃/25℃,光照与黑暗时间均为12 h。3 d后挑选长势良好的幼苗移入减去底部的PCR板中,每板20株,置于水培盆中。待幼苗三叶期后分别进行正常氮(5 mmol·L-1,NN)和低氮(0.5 mmol·L-1,LN)处理,同时叶面喷施50 mg·L-1色氨酸(Trp),以喷施清水为对照(NTrp),并加入0.1%吐温(Tween)作为黏合剂,连续喷施3 d,每天上午9:00进行叶面喷施,喷施状态均以叶片表面形成水滴而不掉落为准。每个处理设置4个生物学重复,喷施外源色氨酸后10 d,选取长势一致的高粱幼苗,取上数第1片完全展开叶进行相关指标的测定。
1.2 测定项目和方法
1.2.1 叶面积 每个重复中随机选取3株幼苗叶片,重复4次,每个处理共12株高粱幼苗叶片用于测量叶面积,采用长宽系数法测定叶面积。
1.2.2 叶片显微结构 每个处理选取4株高粱幼苗叶片,采用石蜡切片的方法对叶片显微结构进行观察。切取高粱幼苗叶片15—20 mm,用固定液固定。将固定好的叶片脱水、透明、包埋、切片、番红染色2 h,用不同梯度酒精脱色、固绿染色、二甲苯透明及中性树胶封片。使用扫描仪(Pannoramic DESK,P-MIDI,P250匈牙利)和扫描软件(Pannoramic Scanner)分别对切片进行扫描和处理。
1.2.3 叶片色素含量 每个重复中随机选取4株幼苗叶片,重复4次,每个处理共16株高粱幼苗叶片用于测量叶片色素含量,使用打孔器(直径1.2 mm)对高粱幼苗叶片进行取样,取样后将叶片放入装有10 ml 80%丙酮溶液的不透光试剂瓶内,黑暗保存24 h提取总叶绿素。采用分光光度计(UV-2550,岛津,日本)法分别测定叶绿素a、叶绿素b、总叶绿素及类胡萝卜素含量(mg·L-1)。
2.3.3 外源色氨酸对低氮胁迫下高粱苗期叶片总氮含量的影响 如图7所示,喷施外源色氨酸与未喷施色氨酸时,与正常氮条件下相比,低氮胁迫显著降低了叶片总氮含量;未喷施色氨酸时398B和CS3541叶片总氮含量分别下降67.09%和74.77%;低氮胁迫下,与未喷施色氨酸相比,喷施外源色氨酸398B和CS3541叶片总氮含量显著提高了24.81%和41.04%,不同自交系间、低氮处理和外源色氨酸三者交互作用显著,但两两之间不显著。
1.2.5 荧光参数 选取高粱幼苗上数第1片完全展开叶,每个处理4次重复。蒸馏水洗净后用滤纸吸干表面水分,于室温暗处理30 min后,采用FlourCam FC 800-O/2020叶绿素荧光仪(Brno, Czech Republic)对叶片进行拍照。测定的参数设置为:用小于0.1 μmol·m-2·s-1的测量光照射叶片,测得初始荧光(0),随后施加饱和脉冲光(10 000 μmol·m-2·s-1,0.7 s),测得最大荧光(m),光适应15 min(800 μmol·m-2·s-1)后获得最大荧光m′。获取的主要参数有:0(初始荧光)、m(最大荧光),v=m–0(可变荧光),v/m(PSⅡ最大光化学效率或原始光能转换效率),(m–m′)/m′(NPQ热耗散)。
软件工程专业是目前很多高校开设的计算机类专业,其主要以培养软件人才为目标,随着社会的发展,高素质软件人才的需求越来越大,而我国软件人才的培养,尤其是高校高素质软件人才的培养和社会需求有较大的差距,从而造成了高素质软件人才的匮乏。由此可见,建设好高校的软件工程专业是培养社会需要的高素质软件人才的重要保障。
采用Excel 2016对数据进行整理,使用Graph Pad Prism 8作图,用SPSS21.0对各性状进行方差分析,以Duncan法检验处理间差异显著性。不同小写字母表示不同处理间在<0.05水平下差异显著。
1.2.7 氮代谢相关指标 将叶片烘干粉碎后,采用浓硫酸消煮,凯氏定氮法测定全氮(N)含量。碳氮比(C/N)=全碳含量/全氮含量。硝酸还原酶活性(nitrate reductase,NR)和亚硝酸还原酶(nitrite reductase,NiR)活性参照李合生[23]的方法进行测定。谷氨酰胺合成酶(glutamine synthetase,GS)和谷氨酸合成酶活性(glutamate synthase,GOGAT)分别参照Gong等[27]和Liu等[5]的方法进行测定。
1.2.8 定量PCR检测衰老相关基因表达量 称取叶片鲜样25—100 mg,选用多糖多酚植物总RNA提取试剂盒(TSP412)提取叶片总RNA。使用逆转录试剂盒Goldenstar RT6 cDNA Synthesis Mix进行反转录扩增,反转录得到的cDNA产物3倍稀释后作为qPCR模板,qPCR在杭州博日FQD-96A仪器中进行,反应体系(20 μL)为:cDNA 1 μL、SYBR Green 10 μL、Primer F 1 μL、Primer R 1 μL和ddH2O 7 μL。qPCR反应条件为:95℃ 2 min;95℃ 15 s,60℃ 15 s,72℃ 20 s,40个循环。根据扩增的循环数(CT值),采用2-ΔΔct方法对数据进行相对定量分析。用于qPCR的基因ID和引物信息如表1所示。
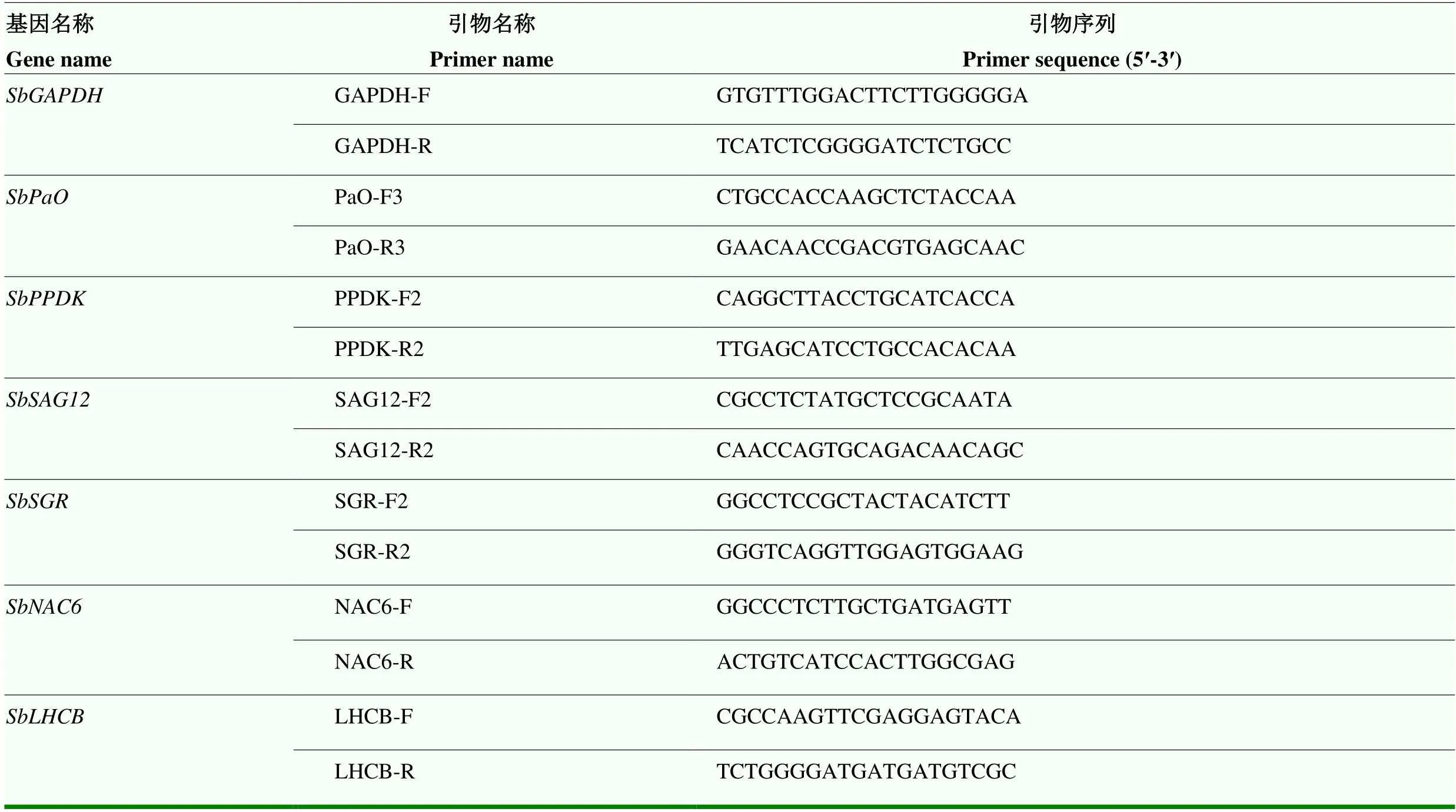
表1 衰老相关基因引物序列
1.3 数据分析
1.2.6 碳代谢相关指标 采用硫酸蒽酮比色法测定可溶性糖、蔗糖和淀粉含量[23]。称取叶片鲜样0.1 g,放入80%乙醇溶液中进行提取,收集上清液,用80%乙醇定容到20 mL,分别用于测定可溶性糖和蔗糖含量,沉淀用于淀粉含量的测定。全碳(C)含量=可溶性糖含量+淀粉含量。采用北京索莱宝科技有限公司的测试盒测定蔗糖合酶(sucrose synthase,SS)、蔗糖磷酸合酶(sucrose phosphate synthase,SPS)和转化酶(invertase,INV)活性[24-26]。
“你好,我能帮你点儿什么?”女孩抬起头,满脸的雪,看不清鼻子眼睛,很滑稽。最后只哆嗦着说出两个字:“好冷。”
2 结果
2.1 外源色氨酸对低氮胁迫下高粱苗期叶片形态特征与显微结构的影响
2.1.1 外源色氨酸对低氮胁迫下高粱苗期叶片形态特征的影响 由叶片表型可以看出(图1),低氮胁迫使CS3541第1片叶表现出早衰症状,该症状始于叶尖及中上部叶边缘,起初表现为黄褐色,而后叶片出现皱缩逐渐枯萎。与正常氮相比,低氮胁迫显著降低了398B和CS3541的叶面积,分别减少了30.74%和36.01%;同时,低氮胁迫下398B叶面积显著大于CS3541(36.30%)。与未喷施外源色氨酸相比,正常氮和低氮条件下外源色氨酸处理使398B和CS3541的叶面积显著增加了17.26%、33.29%和36.72%、52.06%。同时,低氮胁迫下外源色氨酸显著增加了398B和CS3541叶干重和叶鲜重,色氨酸与氮素互作对叶片干重和鲜重的影响均达显著水平。
2.1.2 外源色氨酸对低氮胁迫下高粱苗期叶片显微结构的影响 如图2所示,在正常氮条件下,398B和CS3541细胞排列整齐,花环结构清晰,细胞染色较深,尤其398B表现更好。低氮胁迫下,398B花环结构完整,维管束鞘结构清晰可见,叶肉及泡状细胞数量较多;CS3541细胞部分解体,花环结构模糊,维管束缩小,染色较淡。外源色氨酸处理后,2个自交系叶片结构均得到恢复,398B泡状细胞数量增多,利于控制叶片的失水。
在不同浓度标气的情况下,标准CO2气体浓度与六次重复实验的电压比值fi(i为实验次数)测试数据如表1所示。
2.2 外源色氨酸对低氮胁迫下高粱苗期叶片光合特性的影响
2.3.5 外源色氨酸对低氮胁迫下高粱苗期叶片碳氮比值的影响 正常氮条件下,高粱幼苗叶片碳氮比(C/N)维持在0.35—0.50;低氮胁迫使叶片总糖含量增多,而总氮含量减少,从而使叶片C/N显著提高(图9)。同时,未喷施色氨酸时398B和CS3541叶片C/N分别增加了147.42%和188.40%,但喷施外源色氨酸后,398B和CS3541叶片C/N分别降低了35.50%和44.56%。其中,各处理及各处理互作对叶片C/N的影响均达显著水平。
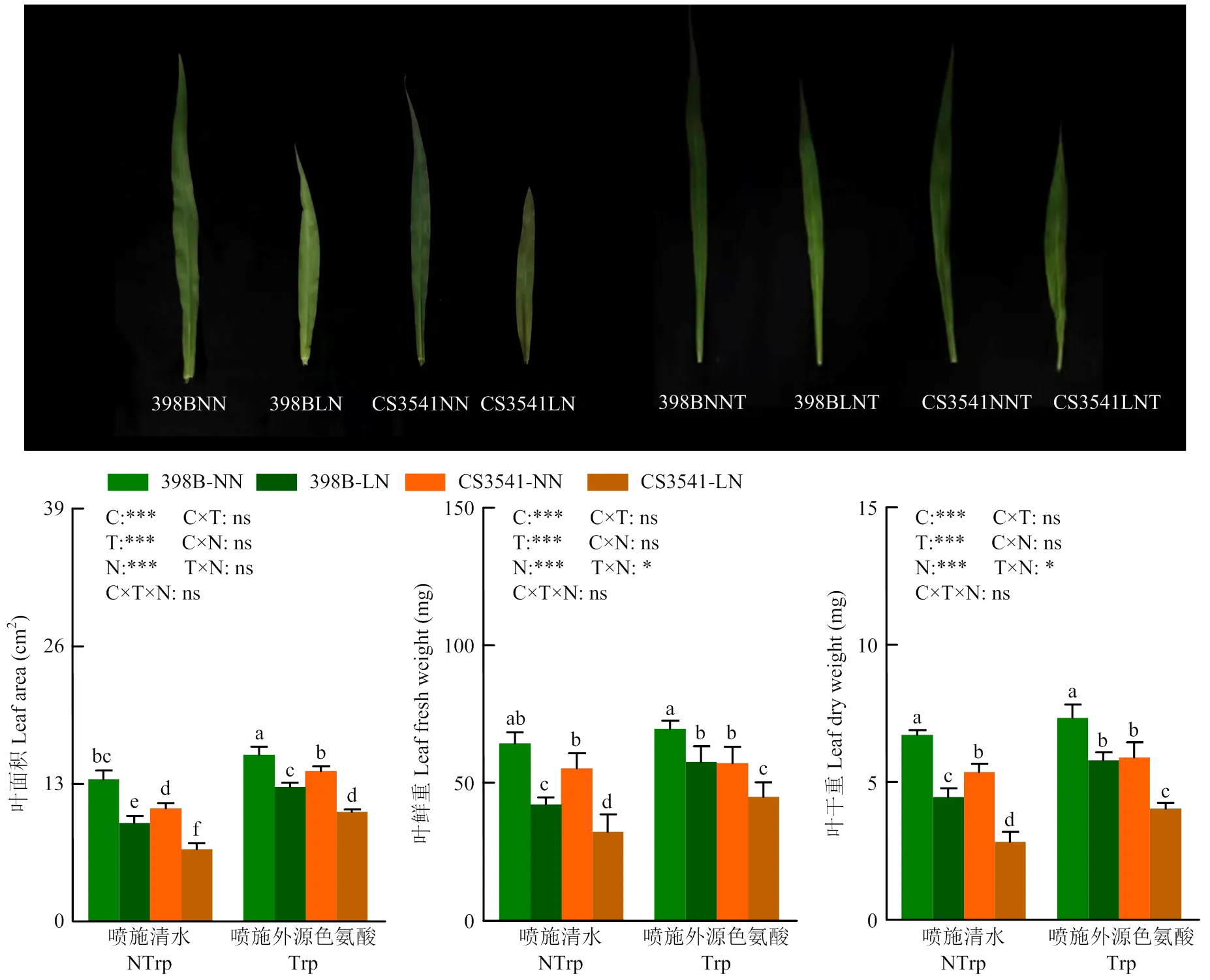
不同小写字母代表不同处理下在5%水平上差异显著性。C:自交系;T:色氨酸;N:氮。*、**和***分别在0.05、0.01和0.001概率水平上差异显著。ns:没有显著差异。NN:正常氮处理;LN:低氮处理;NNT:正常氮外源色氨酸处理;LNT:低氮外源色氨酸处理。下同

表2 外源色氨酸对低氮胁迫下高粱幼苗叶片色素含量的影响
NTrp:喷施清水;Trp:喷施50 mg·L-1色氨酸。NN:正常氮处理;LN:低氮处理;NNT:正常氮外源色氨酸处理;LNT:低氮外源色氨酸处理。表中数据为平均值±标准误;同列数据后不同小写字母表示处理间在<0.05水平差异显著水平
NTrp: spraying with water; Trp: spraying with 50 mg·L-1tryptophan. NN: Normal nitrogen; LN: Low nitrogen: NNT: Normal nitrogen+ tryptophan; LNT: Low nitrogen+tryptophan. The data in the table is the mean of 3 replicates, values followed by different lowercase letters are significantly different at<0.05
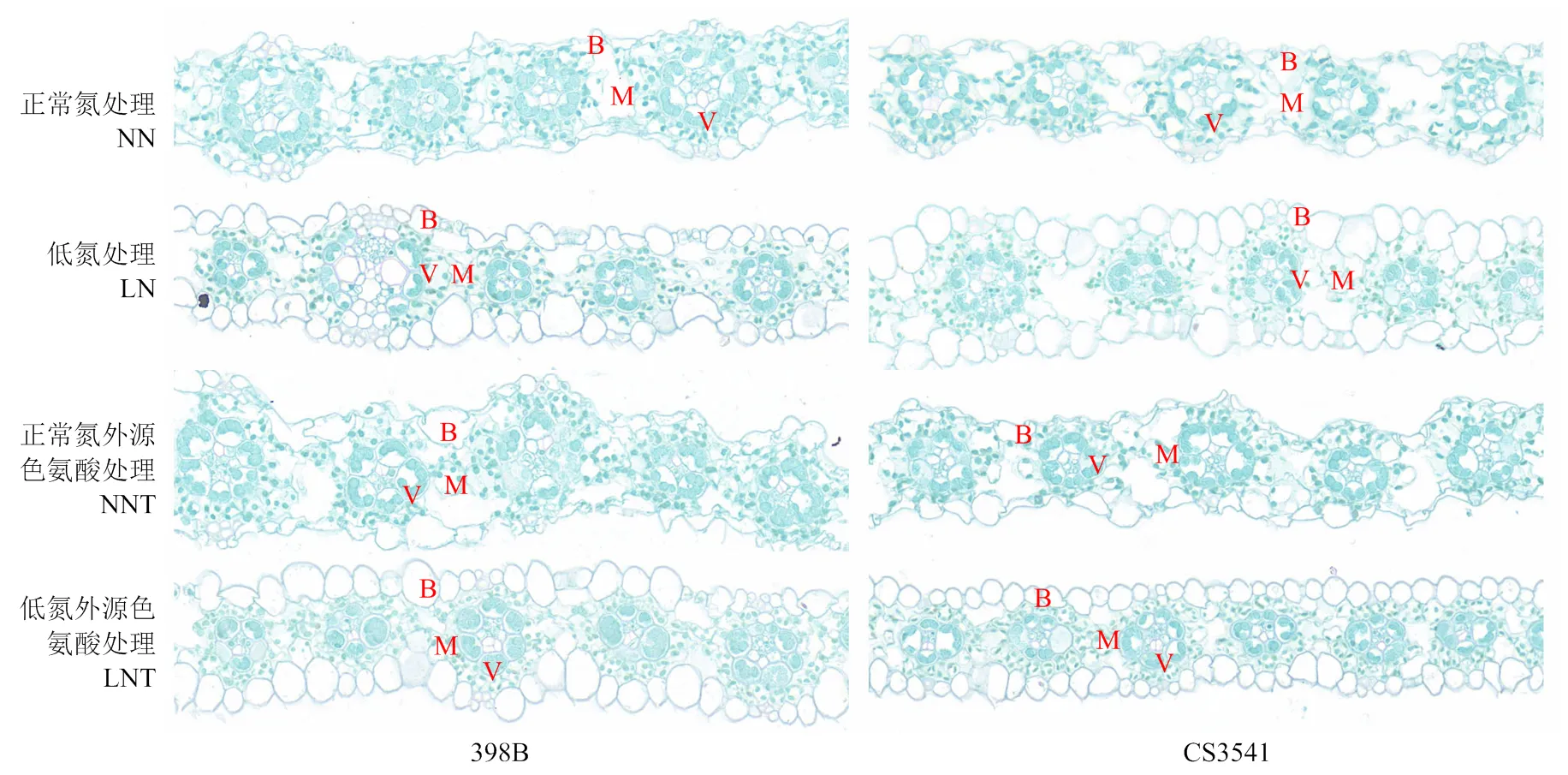
20倍放大,比例尺为50 μm。B:叶泡细胞,V:维管束鞘;M:叶肉组织
那么教师“支架”能起到哪些作用?什么样的教师“支架”才更有效?影响教师“支架”起作用的因素有哪些?本文试图从社会文化理论视角并基于先前实证研究结果对这些问题进行探讨。

图3 外源色氨酸对低氮胁迫下高粱幼苗叶片光合参数的影响
2.2.3 外源色氨酸对低氮胁迫下高粱苗期叶片荧光参数的影响 由图4可知,与正常氮相比,低氮胁迫显著降低高粱幼苗叶片v/m和NPQ,且降幅表现为398B<CS3541;未喷施色氨酸时,398B和CS3541叶片v/m和NPQ分别降低了18.04%、24.59%和9.94%、36.05%;喷施外源色氨酸后,398B和CS3541叶片v/m和NPQ分别提高了13.21%、26.24%和38.95%、63.09%,色氨酸降低了低氮胁迫诱导的高粱叶片的光氧化损伤,从而减缓了叶片衰老。
总之,神经肌电生物反馈在急性期重症脑梗死患者防止其骨骼肌萎缩是一个有效和可行应对策略,但是否会转化为长期好处,如提高生存率、减少住院治疗滞留时间、改善康复结果还需进一步研究。
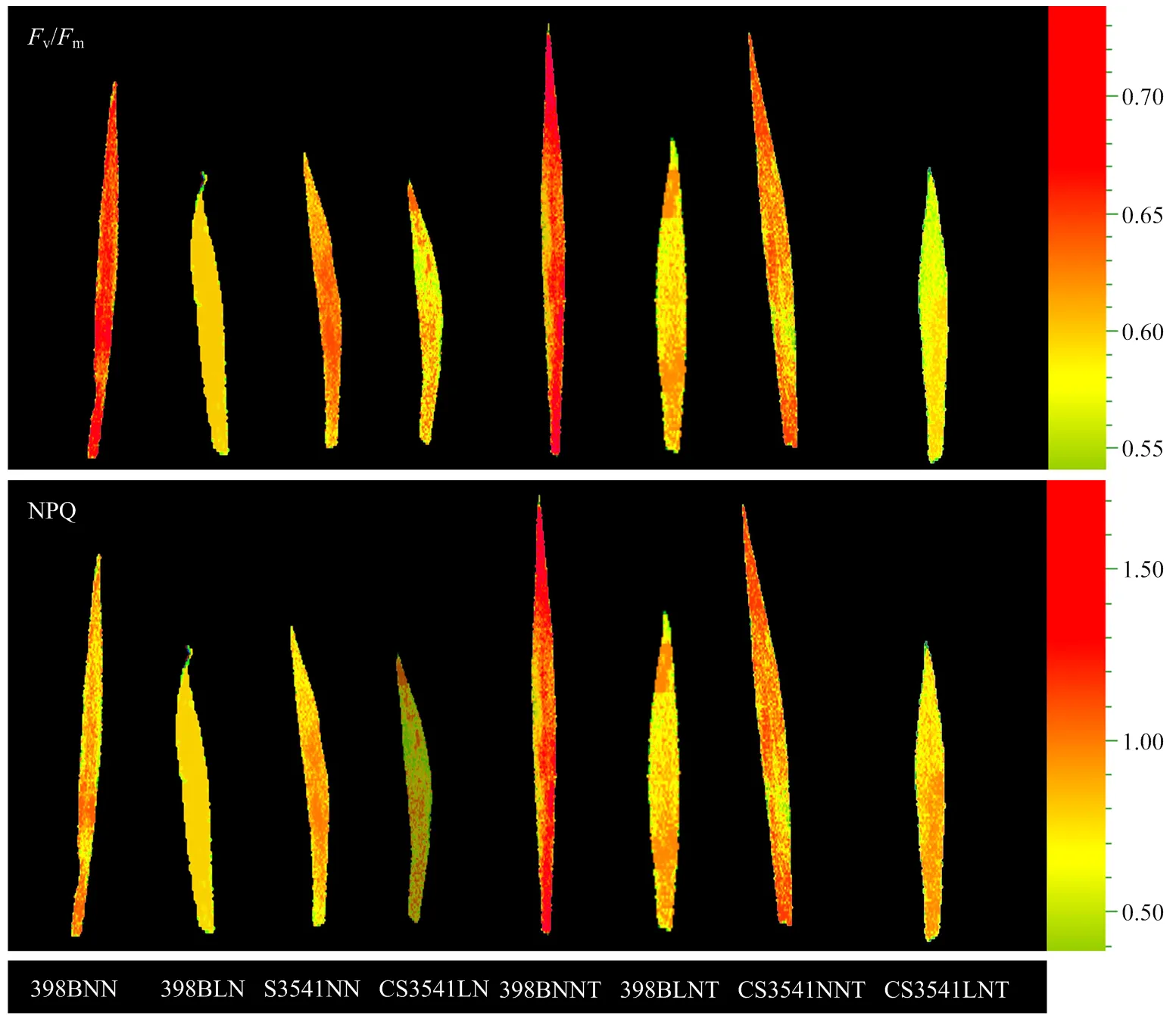
Fv/Fm:PSⅡ最大光化学效率,NPQ:热耗散
2.3 外源色氨酸对低氮胁迫下高粱苗期叶片碳氮代谢的影响
2.3.1 外源色氨酸对低氮胁迫下高粱苗期叶片糖含量的影响 低氮胁迫显著提高了高粱幼苗叶片总糖、可溶性糖、蔗糖和淀粉含量,增幅表现为398B<CS3541(图5)。未喷施外源色氨酸时,低氮胁迫使398B和CS3541叶片总糖、可溶性糖、蔗糖、淀粉含量分别提高了47.93%、14.33%、23.34%、83.65%和51.67%、63.97%、76.89%、66.32%;而外源色氨酸处理显著降低了低氮胁迫下398B和CS3541叶片总糖、可溶性糖、蔗糖、淀粉含量,分别降低24.13%、11.42%、18.76%、34.27%和27.87%、21.92%、47.79%、36.03%。各处理及各处理互作对叶片总糖和蔗糖含量的影响均达显著水平,叶片糖的积累是衡量叶片衰老的另一项重要指标,外源色氨酸降低了糖积累量,从而缓解了低氮胁迫对高粱叶片衰老的影响。
2.2.2 外源色氨酸对低氮胁迫下高粱苗期叶片光合参数的影响 低氮胁迫后,398B与CS3541叶片n、s、r和i均呈现下降趋势,下降幅度有所不同(图3)。与正常氮相比,398B叶片n、s、i和r分别显著降低了43.34%、43.64%、28.34%和43.54%,而CS3541各光合参数分别显著下降了57.12%、67.22%、28.85%和64.77%。无论正常氮与低氮胁迫下,喷施外源色氨酸较未喷施色氨酸均显著提高了各光合参数。其中,低氮胁迫下,喷施外源色氨酸较未喷施,398B与CS3541叶片n、s、i和r分别显著增加1—2倍。因此,外源色氨酸提高了低氮胁迫下高粱幼苗叶片的光合能力。自交系与色氨酸互作,色氨酸与氮素互作,三者之间互作对叶片n的影响均达显著水平;除三者互作,各处理及各处理互作对叶片i的影响均达显著水平;而各处理及处理互作对叶片r的影响均达显著水平。
2.3.2 外源色氨酸对低氮胁迫下高粱苗期叶片碳代谢关键酶活性的影响 低氮胁迫显著提高了高粱幼苗叶片蔗糖合酶(SS)和蔗糖磷酸合酶(SPS)活性,但显著降低了叶片蔗糖转化酶(INV)活性(图 6)。无论正常氮还是低氮处理,外源色氨酸均显著增加了叶片SS、SPS和INV活性。低氮胁迫下,外源色氨酸处理分别显著增加了398B和CS3541叶片SS(6.87%和6.56%)、SPS(6.87%和6.56%)及INV(8.33%和7.14%)活性。各处理及各处理互作对叶片INV活性的影响均达显著水平。由此推测,低氮胁迫下外源色氨酸促进了叶片蔗糖的分解,降低了糖的合成。

图5 外源色氨酸对低氮胁迫下高粱幼苗叶片糖含量的影响
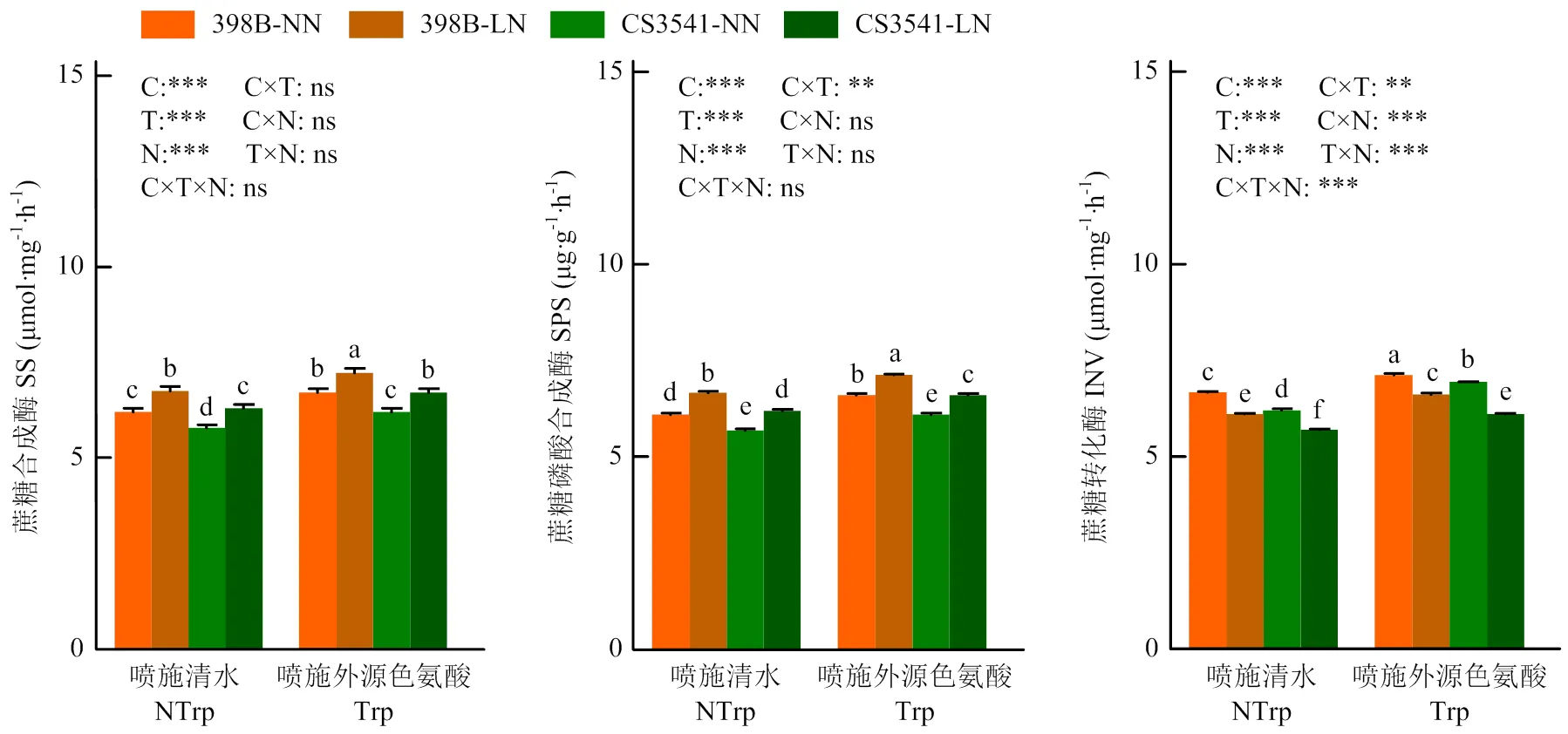
图6 外源色氨酸对低氮胁迫下高粱幼苗叶片碳代谢酶活性的影响
1.2.4 光合相关指标 利用Li-6400光合仪(美国,LI-COR公司)测定高粱上数第1片完全展开叶n(净光合速率)、s(气孔导度)、r(蒸腾速率)和i(胞间CO2浓度),每个处理4次重复。设定的测定参数为光照强度1000 μmol·m-2·s-1,CO2浓度为(385±5)μmol·mol-1,叶面积为2 cm2,温度为28℃。
本文的被解释变量是企业劳动生产率。参考以往涉及劳动生产率的研究文献,特别是实证文献,以人均产出来定义企业的劳动生产率,采用人均工业增加值来度量。
2.3.4 外源色氨酸对低氮胁迫下高粱苗期叶片氮代谢关键酶活性的影响 正常氮和低氮条件下,外源色氨酸处理显著提高了硝酸还原酶(NR)、(谷氨酰胺合成酶)GS和谷氨酸合成酶(GOGAT)的活性,而显著降低了亚硝酸还原酶(NiR)活性(图8)。在低氮胁迫下,外源色氨酸分别显著增加398B和CS3541叶片的NR(7.39%和11.16%)、GS(16.01%和20.50%)及GOGAT(17.04%和23.22%)活性;低氮胁迫下外源色氨酸处理使398B和CS3541叶片NiR活力分别降低了83.60%和87.23%。自交系、色氨酸、氮素对叶片氮代谢关键酶活性的影响均达显著水平;除叶片GS活性外,三者互作对叶片NR、NiR和GOGAT活性的影响均达显著水平。
为适应未来城市化加速发展、工业化深化发展、土地、耕地和水等资源日益短缺、国内人口流动规模不断加大、国际游客不断增多等的态势,新世纪我国养猪业将朝着优质、高效、安全的目标发展,养猪生产将以适度规模化、规范化、标准化、生态循环经济模式为主体,多项技术的综合配套利用是提高养猪业整体生产水平和效益的关键。下面将从育种、营养、猪肉安全生产等方面探讨我国新世纪养猪业的发展。
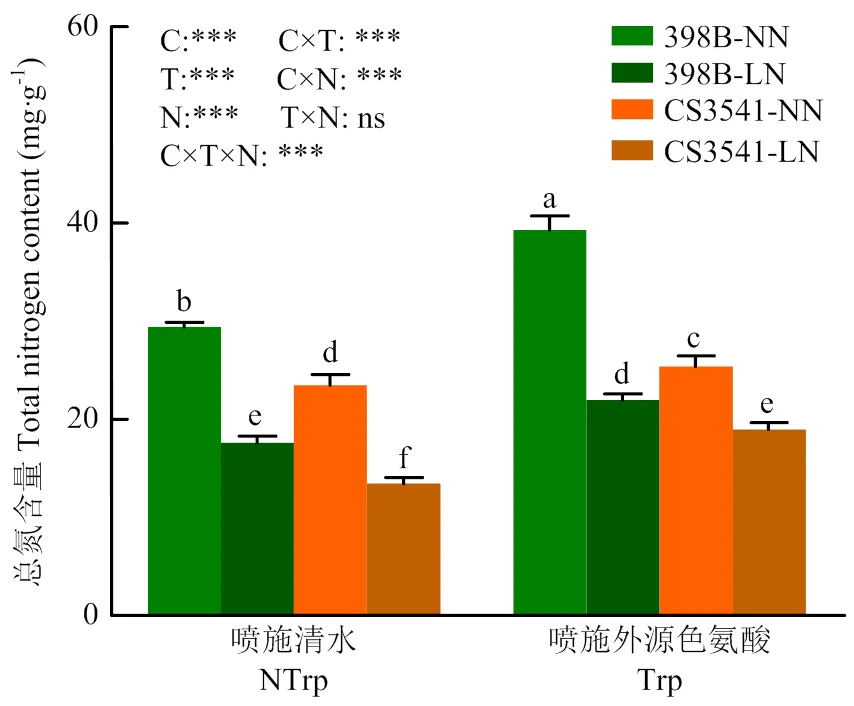
图7 外源色氨酸对低氮胁迫下高粱幼苗叶片氮含量的影响
2.2.1 外源色氨酸对低氮胁迫下高粱苗期叶片色素含量的影响 由表2可以看出,未喷施外源色氨酸时,低氮胁迫分别使398B和CS3541叶片叶绿素a、叶绿素b、类胡萝卜素含量和叶绿素含量显著降低了48.93%、53.41%、58.02%、49.70%和66.10%、133.46%、51.15%、75.34%,且降幅表现为398B<CS3541。低氮胁迫下,外源色氨酸显著提高了398B叶片叶绿素a(36.92%)、叶绿素b(36.50%)、类胡萝卜素含量(53.15%)和叶绿素含量(36.85%),而未显著提高CS3541叶片各色素含量。
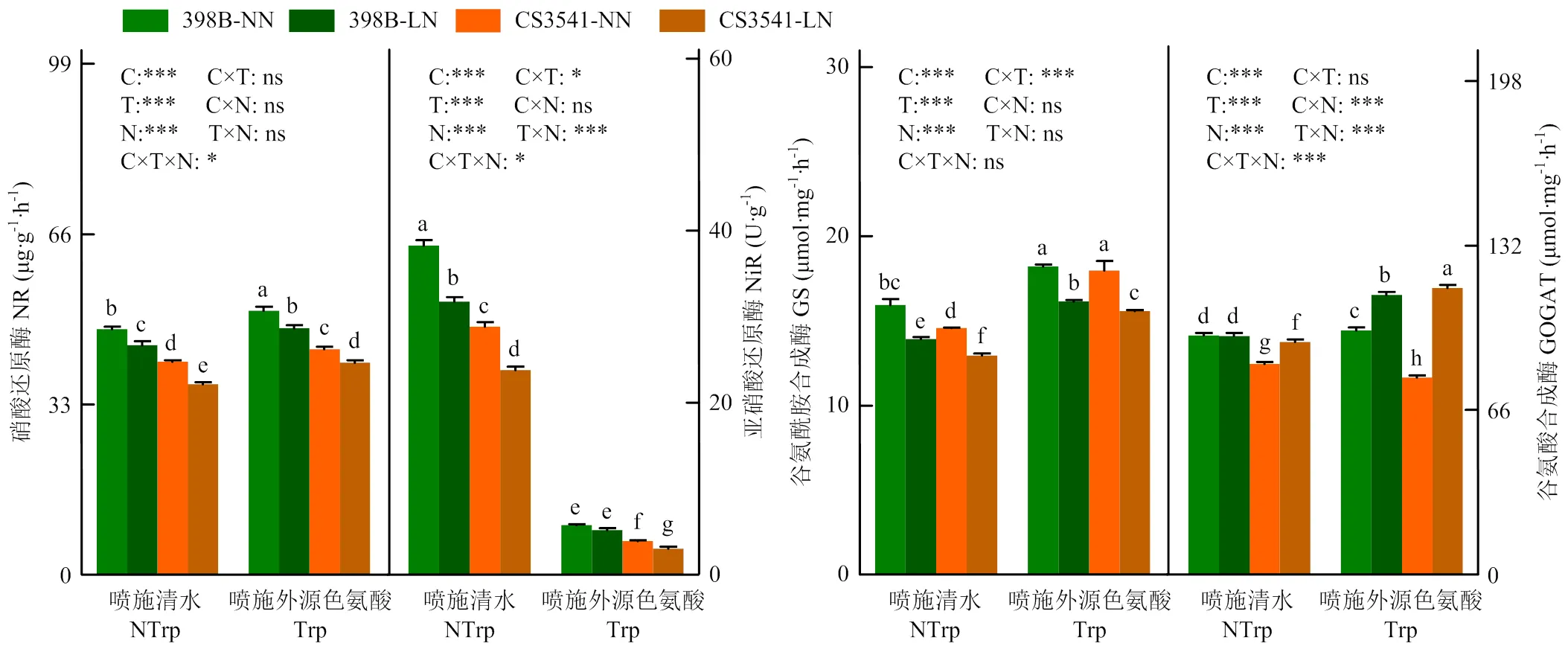
图8 外源色氨酸对低氮胁迫下高粱幼苗叶片氮代谢关键酶活性的影响
2.4 外源色氨酸对低氮胁迫下高粱苗期叶片衰老相关基因表达量的影响
由衰老相关基因表达情况可知(图10),与正常氮条件下398B叶片和表达量相比(均以正常氮条件下398B为对照,作为起点 1,其他同),低氮胁迫使398B叶片两者基因表达量分别下降了36.18%和18.02%,而、、和分别增加了109.53%、13.75%、289.76%和102.32%;相同情况下,低氮胁迫使CS3541叶片和表达量分别下降了54.58%和29.97%,但、、和分别增加了247.84%、79.49%、336.08%和482.09%。低氮胁迫下,与未喷施色氨酸相比,喷施外源色氨酸398B叶片和表达量分别增加了136.70%和176.22%;喷施外源色氨酸CS3541叶片各基因表达量分别增加了90.35%和168.70%。除叶片和外,各处理及各处理互作对叶片、、和表达量的影响均达显著水平。
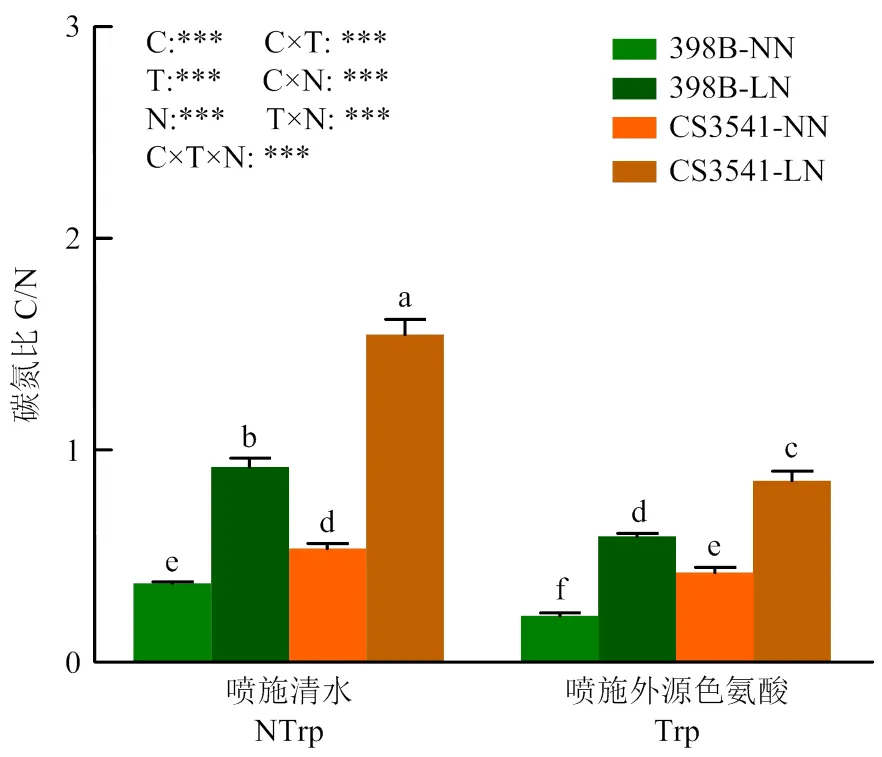
图9 外源色氨酸对低氮胁迫下高粱幼苗叶片碳氮比的影响
2.5 碳氮比值与衰老相关基因之间相关性分析
衰老相关的基因(、、、、和)与叶片C/N之间的关系见图11。叶片C/N与和呈极显著负相关,与、、和呈极显著正相关。另外,与、、和之间呈极显著负相关;与呈显著负相关,与、和之间呈极显著负相关;、和三者之间均呈极显著负相关。
3 讨论
3.1 外源色氨酸对低氮胁迫下高粱幼苗叶片形态的影响
氮素是调控作物叶片形态和衰老进程的重要因素,叶片结构能够反映植物生命活动对低氮条件的响应[28]。在不利环境条件下,叶面积的增加对
猜你喜欢
快乐作文(1.2年级)(2023年9期)2023-05-12 11:31:34
北方牧业(2022年9期)2022-11-22 05:11:22
古今农业(2022年2期)2022-08-15 01:39:52
舰船科学技术(2022年11期)2022-07-15 07:51:56
青年文学家(2022年1期)2022-03-11 12:27:39
西藏农业科技(2019年3期)2019-11-04 00:35:10
动物营养学报(2018年12期)2018-12-13 06:01:02
现代园艺(2018年3期)2018-02-10 05:18:12
上海农业学报(2017年3期)2017-04-10 12:39:12
饲料博览(2014年11期)2014-05-04 10:00:12
