
铝在茶树叶片亚细胞中的分布及其与细胞壁的结合研究
2023-04-05 04:21李春雷徐红梅张汝君马兴云
浙江农业学报 2023年3期
李春雷,徐红梅,刘 杰,张汝君,马兴云,张 华
(1.潍坊科技学院 农学院,山东省设施园艺生物工程研究中心,山东 寿光 262700; 2.山东农业大学 园艺科学与工程学院,山东 泰安 271018)
茶树(Camelliasinensis)是一种非常典型的聚铝植物。据渠心静等[1]调查发现,茶树嫩叶中铝含量能达到3 910 mg·kg-1,老叶中铝含量能达到11 124 mg·kg-1,有的甚至达到了30 000 mg·kg-1[2]。在生产茶园中,茶树叶片中富集如此高含量的铝,却未出现中毒症状,且能生长良好,说明茶树对铝有较强的调节和解毒功能。前人研究结果表明,植物超能力地富集一种或几种元素及其解毒功能与这些富集元素在亚细胞的分布有直接关系。Wu等[3]对大麦亚细胞中的镉分布进行了分析,发现细胞壁和可溶性组分是镉的主要聚集地,周小勇等[4]报道了铅在长柔毛委陵菜中的亚细胞聚集情况,发现细胞壁中占47%~77%,可溶性部分中占13%~45%。铝被植物吸收后主要在细胞壁和液泡中积累。有研究发现,小麦根系中77%的铝累积于细胞壁[5],两种基因型水稻品种的细胞壁中铝含量占总量的80%~96%[6],Zheng等[7]进一步的研究还表明,细胞壁中的铝是被磷固定,潘根生等[8]早年的研究就发现茶树根亚细胞中铝的含量分布情况为:根尖细胞壁>根尖细胞质>根尖细胞核>线粒体。已经探明铝主要聚集在茶树叶片中,而叶片中的铝在亚细胞中又是如何分布?叶片中细胞壁在茶树聚铝耐铝毒中起着什么作用?细胞壁与铝结合可能的机理是什么?为了探明以上问题,本文选取了两个不同聚铝能力的茶树品种,分析了铝在叶片亚细胞中的分布及铝与细胞壁的结合特点,以期能够明确茶树叶片聚铝机理。
1 材料与方法
1.1 试验材料
实验中所用材料为高铝品种农抗早和低铝品种平阳特早的成熟叶片,均采自山东省日照市巨峰镇平家村。
1.2 试验方法
1.2.1 茶树叶片亚细胞组分分离
采用差速离心法[3]。称取鲜叶2.0 g,加入预冷的缓冲液研磨,料液比1∶15,匀浆用100目尼龙纱布过滤,缓冲液淋洗滤渣,滤渣为细胞壁组分,所得的上层清液在1 500×g条件下离心15 min,所得沉淀物即为细胞核和叶绿体组分,收集所得上层清液在15 000×g下离心35 min,所得沉淀物即为线粒体组分,收集所得的上层清液即为可溶性组分(包含核糖体)。以上实验均在低温下进行,冷冻干燥,-80 ℃保存备用。
1.2.2 细胞壁组分分离
取适量细胞壁于离心管中,按料液比1∶20的比例加入pH值7.0、0.5 mol·L-1咪唑溶液提取螯合态果胶,25 ℃、200 g·min-1的条件下振荡提取24 h,6 000×g、20 min条件下离心取上清液,所得到的沉淀物用pH值7.0、0.5 mol·L-1咪唑溶液离心洗涤2~3次,将上清液收集并合并,冷冻干燥;所得到的沉淀物按料液比1∶20的比例,用含20 mmol·L-1CDTA的50 mmol·L-1Na2CO3继续提取碱溶性果胶,在25 ℃条件下振荡提取24 h,离心取上层清液,将所得到的沉淀物用Na2CO3提取液离心洗涤3次,收集上清液并合并,冷冻干燥;沉淀物用4 mol·L-1KOH(料液比1∶20)提半纤维素,于25 ℃恒温环境中振荡提取24 h,离心,取其上清液,加入4 mol·L-1HCl中和至pH值5.0,冷冻干燥;最后将残渣主要为纤维素,冷冻干燥[9]。
1.2.3 细胞壁改性
参考文献[10]的方法。新鲜茶树叶片(去掉中间主脉)在冰浴中用预冷的研钵磨至细胞全部破碎,用-20 ℃、10倍体积的80%乙醇淋洗残渣3次,然后用-20 ℃、10倍体积的甲醇与氯仿等体积混合液淋洗一次,200目纱布过滤,除去水溶性物质,再用-20 ℃、10倍体积的丙酮淋洗除去脂溶性物质,最后12 000×g、4 ℃条件下离心10 min,弃上清液,再加适量蒸馏水,离心,弃上清液,残余物即为细胞壁,经冷冻干燥后备用。
细胞壁甲基化改性:准确称量2.0000 g茶树叶片的细胞壁,放入离心管中,再加入120 mL甲酸和甲醛(体积比2∶1)的混合液。离心管在125 g·min-1条件下振荡反应6 h,然后离心,所得沉淀物经冲洗后冷冻干燥,即可得到甲基化改性成功的细胞壁,4 ℃保存备用。
细胞壁酯化改性:准确称量2.000 0 g茶树叶片细胞壁,放入离心管中,依次加入无水甲醇和浓HCl,体积分别为140 mL和1.2 mL,调整HCl浓度为0.1 mmol·L-1。将离心管在125 g·min-1下振荡12 h,然后离心,将上清液弃去,所得到的沉淀物经冲洗后冷冻干燥,即为酯化改性成功的细胞壁,4 ℃条件储存备用。
1.2.4 细胞壁酶解
果胶酶酶解:准确称量2.000 0 g茶树叶片细胞壁,放于离心管中,加入含0.1% BSA的1%果胶酶120 mL,离心管置于30 ℃的恒温水浴中加热30 min,所得沉淀先经离心,弃去上清液,沉淀用去离子水洗涤干净,再冷冻干燥,于4 ℃条件下储存备用。
纤维素酶解:称取2.000 0 g茶树叶片细胞壁,放置于离心管中,加入1%纤维素酶(含0.1% BSA)120 mL,离心管置于30 ℃恒温水浴中加热30 min,所得沉淀先经离心,弃去上清液,沉淀用去离子水洗涤干净,再冷冻干燥,于4 ℃条件下储存备用。
1.2.5 铝含量分析
采用 ICP-OES 法测定铝元素的含量[11]。
1.3 数据分析
采用 SAS软件进行 LSD显著性检验(LSD法,P<0.05)。
2 结果与分析
2.1 铝在茶树叶片亚细胞中的分布
由图1、图2、图3可以发现,铝在平阳特早和农抗早亚细胞组分中的分布均差异显著(P<0.05),两个品种亚细胞组分铝含量的变化趋势均为细胞壁>细胞核和叶绿体>线粒体>可溶性组分。平阳特早和农抗早均是细胞壁组分铝含量最高,分别为3.63和4.57 mg·g-1,占叶片总铝的46.2%和54.9%;其次是细胞核和叶绿体,分别为2.45和2.02 mg·g-1,占叶片总铝的31.2%和24.2%;再次为线粒体组分,分别为1.45和1.54 mg·g-1,占叶片总铝均为18.5%;铝含量最低的是可溶性组分,分别为0.32和0.20 mg·g-1,占叶片总铝的4.1%和2.4%。

图中同一品种没有相同小写字母表示差异显著(P<0.05),下同。

图2 平阳特早茶树叶片中铝亚细胞分布的相对比例

图3 农抗早茶树叶片中铝亚细胞分布的相对比例
2.2 铝在细胞壁各组分中的含量及分布
进一步分离了平阳特早和农抗早两个品种的细胞壁组分,分析了铝在细胞壁组分中的分布情况(图4、图5、图6)。结果表明,细胞壁的不同成分对铝的聚集能力差异显著。平阳特早和农抗早均是果胶组分中的铝含量最高,分别占细胞壁总铝的78.7%和60.1%,其中螯合态果胶组分中铝分别占细胞壁总铝的65.6%和51.5%,碱溶性果胶中铝分别占细胞壁总铝的13.1%和8.6%,纤维素组分中铝分别占细胞壁总铝的18.1%和31.3%,半纤维素中铝分别占细胞壁总铝的3.2%和8.6%。

图4 铝在茶树叶片细胞壁各组分的分布

图5 平阳特早茶树叶片细胞壁各组分中铝分布的相对比例
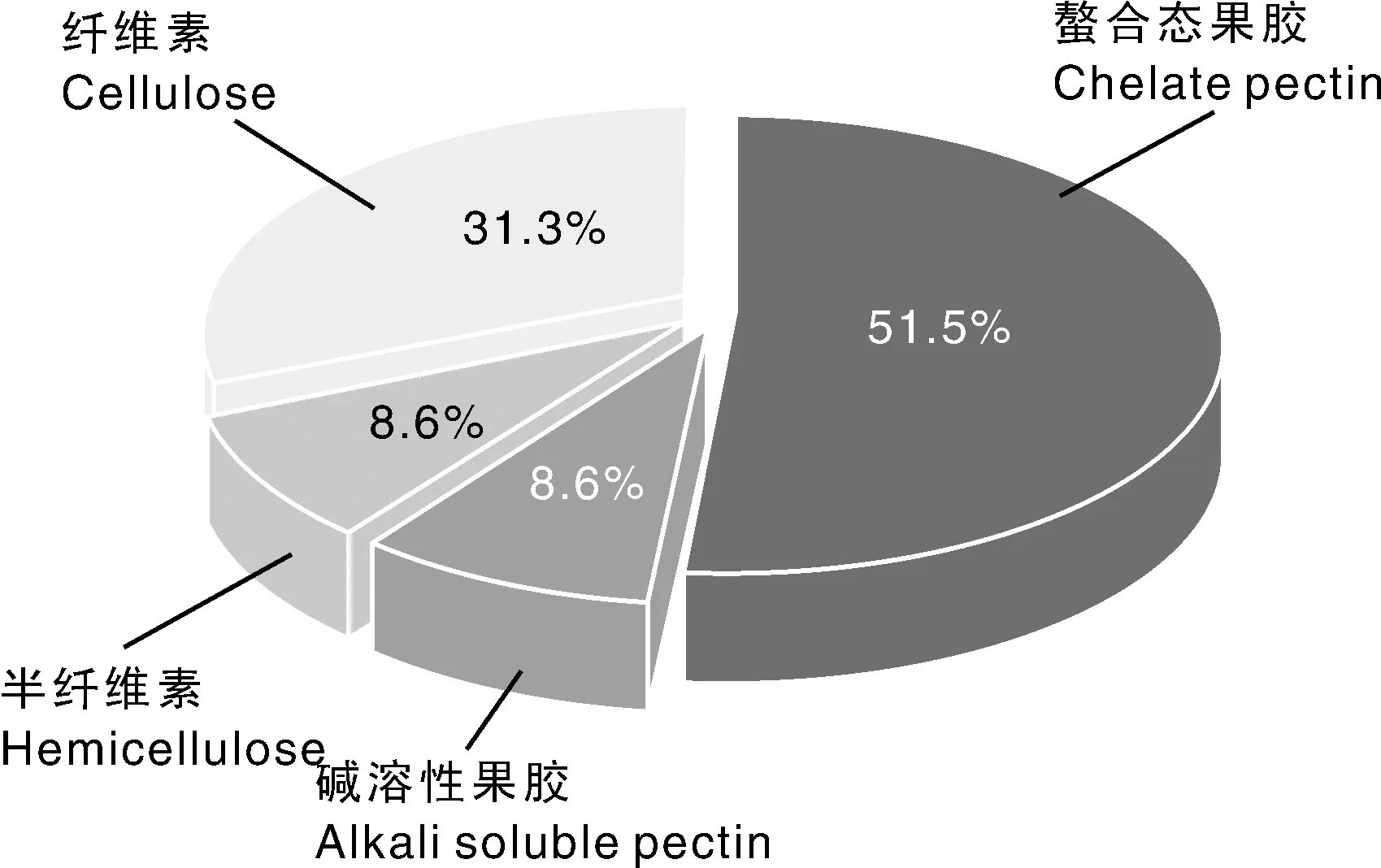
图6 农抗早茶树叶片细胞壁各组分中铝分布的相对比例
2.3 改变细胞壁结构对结合铝的影响
细胞壁结构的改变,对富集铝也有一定影响。因此,对茶树叶片提取出来的细胞壁进行了甲基化、酯化和酶解等改性处理。由结果(图7)可以看出,平阳特早和农抗早叶片细胞壁经甲基化改性处理和酯化改性处理后,两个品种细胞壁中的铝均比对照显著降低,而果胶酶解和纤维素酶解后,两品种细胞壁中的铝含量与未酶解处理相比差异不显著。其中,甲基化处理后,平阳特早和农抗早细胞壁中的铝比对照分别下降了65.5%和67.1%,酯化处理后,两品种细胞壁中的铝比对照分别下降了36.9%和48.2%。这说明,甲基化和酯化处理的细胞壁结构发生的变化对铝和细胞壁的结合有较大影响,而果胶组分和纤维素组分分子链的大小及长短对细胞壁与铝的结合影响不大。

图中同一品种同一处理不同小写字母表示差异显著(P<0.05)。
3 结论与讨论
茶树中聚集着高含量的铝,未出现中毒的症状,这些现象表明,茶树有着自身的耐铝机制和解毒机制。本文研究了茶树叶片中铝在亚细胞中的分布,结果表明:农抗早、平阳特早这两个茶树品种中均表现为茶树叶片细胞壁铝含量最高,占细胞总铝的46.2%~54.9%,这与前人的研究结果一致。Gao等[12]发现,在茶树根中,70%的铝聚集在根细胞壁中,在茶树叶片中,75%的铝聚集在叶片细胞壁中。Hajiboland等[13]采用共焦激光扫描显微镜发现,细胞壁中的荧光信号要高于其他细胞组分。Tolrà等[14]采用LEXRF技术对铝在茶树叶片亚细胞中分布的研究表明了铝优先储存在上表皮细胞壁,其次是液泡。Carr等[15]也得到了同样的发现,在细胞壁中发现了高浓度的铝。这些研究成果均证实了铝主要分布在细胞壁中,细胞壁作为保护内部细胞代谢的第一道屏障,阻挡了铝进入细胞内部,有效地减小了铝对细胞内部细胞器的伤害,这正是茶园中茶树未出现铝中毒症状的重要原因,当铝浓度过高,超出了细胞壁的固定能力,进入细胞的铝就会增多,造成对细胞结构的伤害,这在我们前期实验中已经被证实,Li等[16]通过透射电镜观察不同铝浓度处理水培茶苗叶片的亚细胞结构发现,在铝浓度小于0.32 mmol·L-1时,茶苗细胞超微结构保持完整,未受到铝的破坏,当铝浓度达到0.53 mmol·L-1时,细胞膜和叶绿体膜被完全破坏,泪囊体扭曲甚至溶解。铝对细胞内部结构的影响也与相关的生理指标相对应,丙二醛(MDA)是膜脂过氧化物反应程度的指标,在我们前期实验中,其含量在小于0.32 mmol·L-1时,呈下降趋势,说明细胞内部的膜系统未受到伤害,而当铝浓度达到0.53 mmol·L-1时,MDA含量显著升高,说明细胞内部的膜系统受到了破坏,这印证了铝处理下通过透射电镜观察的细胞超微结构的结果。另外,叶绿体、膜系统及泪囊体正常的排列方式是光合作用和叶绿素形成的保障,当铝浓度较高时,细胞结构遭到破坏,也将会影响光合作用和叶绿素的合成,这也得到了证实[17]。Li等[18]研究发现,铝浓度达到1.0 mmol·L-1处理茶苗1 h,茶苗就会出现铝毒害症状,我们前期的实验发现,铝浓度在0.32 mmol·L-1以下对茶苗是安全的,超过这个浓度就会对细胞结构造成伤害,因此,铝被固定在细胞壁上在一定程度上保护了细胞内部结构不受铝的伤害,这也是茶树耐铝的一个重要原因。
植物细胞壁是一个由多糖、蛋白质和木质素等组成的复合体,这些多糖和蛋白质等生物大分子含有大量羧基、氨基等配位基团,可以为离子提供大量的结合位点。本文结果表明,细胞壁组分中果胶组分的铝含量最高,占到细胞壁总铝的60.1%~78.7%,其中大部分在螯合态果胶,占细胞壁总铝的51.5%~65.6%,其次是纤维素中铝含量也较高。通过改变细胞壁结构,发现甲基化改性处理后铝含量显著降低,酯化改性处理后,铝含量也显著降低,而果胶酶解和纤维素酶解处理后,铝含量变化不明显,说明了铝主要结合在细胞壁中的-COOH和-NH2上,而细胞壁中多糖分子链大小对结合铝的作用不大。
综上试验结果,铝主要存在于茶树叶片的细胞壁中,推测主要以氢键的形式结合在细胞壁果胶中的-COOH和-NH2上,大部分的铝被固定于细胞壁中,减少了铝进一步进入细胞内部,从而降低了铝对细胞内部结构的伤害,这正是茶树解铝毒的一个重要机制。
猜你喜欢
酿酒科技(2023年10期)2023-11-23
建材发展导向(2022年20期)2022-11-03
建材发展导向(2022年12期)2022-08-19
建材发展导向(2021年20期)2021-11-20
矿产勘查(2020年1期)2020-12-28
食品安全导刊(2019年27期)2019-12-09
名城绘(2019年4期)2019-10-21
浙江工业大学学报(2017年5期)2018-01-22
广东农业科学(2017年5期)2017-08-29
饲料博览(2015年12期)2015-04-04
