
‘阳光玫瑰’葡萄冬果采后叶片光合同化物转运能力研究
2023-04-01 10:48:12覃富强王博韩佳宇黄秋秘廖原张延晖岑佳慧陈志宁白先进曹雄军
中外葡萄与葡萄酒 2023年2期
覃富强,王博,韩佳宇,黄秋秘,廖原,张延晖,岑佳慧,陈志宁,白先进,曹雄军
(1. 广西农业科学院葡萄与葡萄酒研究所,广西南宁 530007;2. 广西大学农学院,广西南宁 530004;3. 广西生态工程职业技术学院,广西柳州 545004;4. 广西真诚农业有限公司,广西南宁 530007)
叶片衰老是植物为适应环境而进化的自然现象,具有重要的生物学意义[1]。叶片衰老是叶片由绿转黄的过程,叶绿素含量逐渐下降,光合能力逐渐降低,光合同化物合成能力逐渐降低。一般分为三个时期:(1)诱导期:在环境或激素等因子诱导下启动或加速叶片衰老;(2)重组期:细胞组分中大分子物质如蛋白质、核酸、酯类等物质开始降解;(3)终止期:细胞发生自溶反应,细胞器开始裂解和死亡[2]。叶片衰老时最明显的外观形态变化是颜色由绿转黄,因此人们常用形态指标来对叶片衰老进行判断。然而,从外观形态判别叶片衰老程度受主观意识的影响较大。研究发现,光合能力下降是叶片衰老的实质性特征,因此常根据叶片净光合速率、胞间CO2浓度、气孔导度、蒸腾速率、细胞超微结构、叶绿素和蛋白质含量等指标判断叶片的衰老程度[3-4]。随着植物衰老,叶片光合能力下降,光合同化物合成减缓[5]。
13C稳定同位素示踪技术在植物光合产物转运、分配、利用等研究中被广泛利用。在葡萄、苹果、油茶等植物中均有应用13C稳定同位素示踪来研究光合同化物的去向和分配情况的报道[6-8]。
一年两收栽培模式下,因冬果成熟采收后很快开始冬季修剪,进行下一年度的生产。采后修剪时期不同,叶片的衰老程度就不同,进而会影响树体叶片光合同化产物的回流,研究‘阳光玫瑰’葡萄冬果采后不同衰老程度叶片光合能力和光合同化物转运能力对冬果采后修剪时间的确定具有重要的生产指导意义。
1 材料与方法
1.1 试材及地点
试验在广西农业科学院明阳葡萄示范基地内进行。以3年生树体长势良好的一年两收栽培两代不同堂模式‘阳光玫瑰’葡萄为试材,株行距2 m×4 m,T形架式,钢架大棚避雨栽培,土壤为红黄壤,按照当地两收栽培模式常规管理。该地属于亚热带季风气候,示范区冬果生长季光照时数为785.6 h、平均温度为21.8 ℃、有效积温3486.1 ℃。
1.2 试验设计
试验于冬果采收后第7天(2021年12月27日)进行,选取衰老程度不同的叶片和枝条作为处理,衰老等级参考刘道宏[12]划分为全绿、半绿、全黄,作为3个处理,如图1所示。在园区内随机选取长势一致的枝梢,把第4节位以下的叶片摘除,并通过修剪使每条枝保留6片叶,将整个枝梢的叶片分为全绿、半绿和全黄,每处理3枝,测定枝梢第8节位叶片光合参数。为探究叶片无光环境下是否有光合同化物的合成与转运,设计一个完全遮光13C处理。遮光处理枝梢叶片为全绿状态,随后利用13C稳定同位素标记以上枝梢全部叶片,测定标记枝条和叶片的13C丰度。同时,另选叶片为全绿状态的3个枝梢,不进行13C稳定同位素标记,作为对照(CK)用于检测自然条件下枝条和叶片13C丰度。

图1 ‘阳光玫瑰’葡萄不同衰老程度叶片外观形态特征Figure 1 Leaf appearance morphology of 'Shine Muscat' grapevine with different senescence degrees
1.3 测定项目与方法
1.3.1 叶片光合参数的测定
采用人工光源测定叶片净光合速率以反映叶片净光合速率理论值[13]。采用CIRAS-3光合仪(PP systems,USA)在人工光源(红∶蓝∶白=90∶5∶5,1200 μmol·m-2·s-1)下,于上午9:00—11:00对所选叶片测定净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)等。
1.3.213C同位素标记方法
标记前在枝条基部第1至2节位节间处进行环割,宽度5 mm,深度达木质部,防止光合同化物往其他部位转运。标记枝条与叶片置于透明塑料袋(遮光处理使用不透光黑色塑料袋)和金属框架制作的反应室内,将50 mL的反应瓶和电风扇挂在两端,反应瓶中加入0.6 g Ba13CO3(98%13C,上海化工研究院)+0.74 g BaCO3。标记时向反应瓶中注射6 mL 40%的乳酸溶液产生13CO2,为了确保产生的13CO2被叶片充分吸收,同时添加0.74 g BaCO3到反应瓶中,开启电风扇以促进反应室内的气体流通。标记24 h后从枝条基部环割处剪下,分解为叶片、枝条,分装并带回实验室。样品按清水、洗涤剂、清水、1%盐酸溶液、去离子水的顺序冲洗干净,于105 ℃下杀青30 min,随后在80 ℃下烘至恒重,粉碎过100目筛,每个样品充分混匀后装袋备用。
来自泰国海洋局、港务局局长Pongwish Innmai 向记者表示,这次学习交流很有意义,受益匪浅,加深了我对中国海上交通安全监管工作的认识,对佛山海事的智慧海事管理工作感到印象深刻,尤其是近几年水上安全事故的大幅下降。在危险天气预警,船舶违章处理等方面可以向佛山海事学习。希望可以继续加强中国和东南亚国家船舶安全管理经验交流,以提升自己国家的船舶安全管理能力。
1.3.313C测定与数据计算方法
测定时称取2.0~3.0 mg干样置于锡囊中,将装好待测样品的锡囊导入元素分析同位素质谱联用仪(Elementar-Sercon Integra 2,UK)测定样品中13C的相对丰度(δ13C)和总碳含量(C)。
根据公式δ13C=[(RS-RPBD)/RPBD]×1000、RS=13C/12C计算样品中的13C丰度,其中RPBD=13C/12C=0.0112372(自然环境中的13C与12C的比值)。
总含碳量计算公式:C(%)=20/[(W样品/W标样) ×(BA样品/BA标样)]×100
其中,W样品(样品质量,μg);W标样(标准物质量,μg);BA样品(样品峰面积);BA标样(标准物峰面积)
各器官含碳量:Ci(g)=各器官干质量×C
标记样品中13C的占比:Atom(%)=(δ13C+1000)×RPBD/[(δ13C+1000)×RPBD+1000]×100
器官的13C量:13Ci(mg)=Ci×[Atom(%)13C标记-Atom(%)13C自然]×1000/100
13C在各器官分配率:13Ci(%)=13Ci/13Ci净吸收×100
1.4 数据处理
试验数据使用SPSS 18.0软件进行数据统计分析,Microsoft Excel 2016对数据整理并作图。
2 结果分析
2.1 ‘阳光玫瑰’葡萄不同衰老程度叶片的光合能力
由图2A看出,全绿、半绿、全黄3种叶片净光合速率存在显著差异,全绿叶片的净光合速率最高。为2.33 μmol·m-2·s-1;全黄叶片的净光合速率最低为 ﹣2.51 μmol·m-2·s-1。
叶片蒸腾作用是指水分由细胞蒸发到细胞间隙,然后扩散至植物体外的过程,产生的拉力能为植物营养物质、水分运输提供动力保障[14]。如图2B所示,不同衰老程度叶片之间的蒸腾速率存在显著差异,随着叶片黄化程度的加深,其蒸腾速率逐渐降低。全绿叶片的蒸腾速率为1.49 mmol·m-2·s-1,全黄叶片蒸腾速率仅为0.34 mmol·m-2·s-1。
气孔导度能反映叶片与外界气体交换的能力,气孔导度数值越大叶片光合能力越强[9]。如图2C所示,随着叶片黄化程度的加深,气孔导度不断降低。全绿叶片的气孔导度为64.9 mmol·m-2·s-1,是全黄叶片的2.3倍。
Ci能够反映植物叶片CO2的滞留量,该值越高,代表叶片对CO2的利用率越低[15]。由图2D可知,随着叶片黄化程度的加深,胞间CO2浓度不断升高,全绿叶片胞间CO2浓度为260.70 μmol·m-2·s-1,半绿叶片的为366.92 μmol·m-2·s-1,全黄叶片的为565.2 μmol·m-2·s-1。
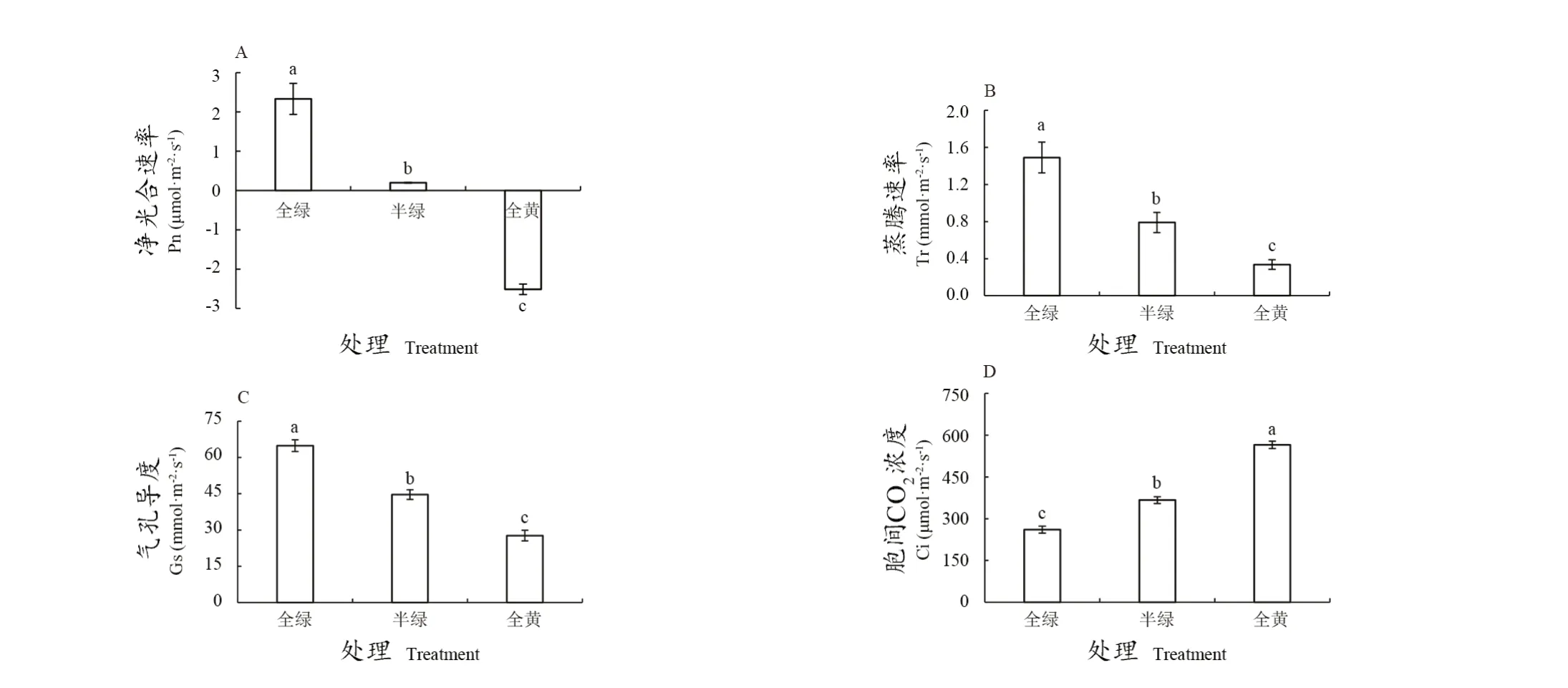
图2 ‘阳光玫瑰’葡萄不同衰老程度叶片光合参数Figure 2 Photosynthetic parameter of 'Shine Muscat' grape leaf with different senescence degrees
2.2 不同叶片各器官中13C含量与分配率
表1为各器官中13C标记新合成的同化物与器官中原有同化物的比例。叶片和枝条中13C的比例均为全绿>半绿>全黄。另外,对于叶片中的13C比例,全黄叶片显著高于遮光13C处理和对照,而遮光13C处理和对照无显著性差异;对于枝条中的13C比例,全黄叶片、遮光处理以及对照中的13C比例均无显著差异。

表1 不同衰老程度叶片和枝条中13C比例Table 1 Proportion of 13C in leaves and fruiting branches with different senescence degrees %
如表2所示,全绿叶片中13C含量为14.534 mg,枝条中的13C含量为2.762 mg,均显著高于其他处理。半绿叶片中13C含量是全黄叶片的12.25倍,半绿叶片的枝条中13C含量是全黄叶片下枝条的20.80倍;全黄状态下叶片中13C含量显著高于遮光处理及对照。这表明‘阳光玫瑰’葡萄采后叶片处于半绿状态时,叶片仍有合成光合同化物并向枝条转运的能力,而全黄叶片仍具有微弱的积累光合同化物的能力。

表2 不同衰老程度叶片和枝条中13C含量Table 2 13C content in leaves and branches with different senescence degreesmg
如表3所示,3个不同衰老程度的叶片将大多数13C同化物分配到叶片当中。其中,全绿叶片13C分配率达到84.03%。遮光处理及对照的13C同化物大多分配到枝条中,且两者之间无显著差异。这些数据表明,虽然半绿叶片仍具有将光合同化物向枝条转运的能力,但由于光合能力降低,其枝条13C分配率高于全绿叶片。而对比同为光合能力缺失或抑制的全黄叶片和遮光处理的13C分配率,进一步验证了全黄状态叶片已基本丧失将光合同化物向枝条转运的能力。

表3 不同衰老程度叶片和枝条中13C分配率Table 3 Distribution of 13C in leaves and branches with different senescence degrees%
3 讨论与结论
叶绿素含量和净光合速率下降是叶片衰老的显著特征。唐桓伟[16]的研究表明,‘阳光玫瑰’冬果叶片净光合速率随叶龄的增加先升高后降低,净光合速率在叶龄53 d时达到最高,叶龄66 d后急剧下降。本研究中冬果采后第7天全绿叶片净光合速率为2.33 μmol·m-2·s-1,该状态下叶片光合强度大于呼吸强度。
葡萄生长前期光合同化物主要集中转运分配到地上部的新生器官,中期光合同化物在地上部与地下部分配比较均衡,而后期则稳定回流至主蔓和根系[17-18]。本试验表明,冬果采后第7天叶片为全绿状态的枝条中13C含量为2.762 mg;叶片为半绿状态的枝条中13C含量为2.33 mg,显著高于全黄状态和遮光处理枝条13C含量,说明半绿叶片仍有合成光合同化物并向枝条转运的能力;而从13C分配率得知,全黄叶片将光合同化物向枝条转运的能力低下。由于构成大气CO2的碳有12C、13C、14C,自然界中12C的CO2约占98.89%,13C的CO2占1.11%,14C的CO2则相对较低,其丰度仅占碳总量的10-12[19],因此自然状态下在植株中仍能检测到13C。试验中全黄叶片的枝条中13C含量显著低于自然状态下的枝条,进一步说明全黄叶片已基本丧失将光合同化物向枝条转运的能力。
一年两收中夏果、冬果不重叠模式第二茬果(冬果)一般12月中下旬采收,翌年1月进行修剪[20]。生产过程中,有时会在叶片还未失绿黄化时进行修剪,或者在叶片已失绿黄化仍然未修剪。试验结果表明,从光合同化物转运角度考虑,在一年两收栽培过程中,理论上可延长到枝条叶片失绿状态时再进行修剪,这样更有利于营养回流和树体树势的恢复,而对于已经失绿黄化的叶片,可及时摘除。
猜你喜欢
中学生数理化·高一版(2020年6期)2020-07-25 01:47:24
童话世界(2019年31期)2019-11-25 09:51:12
小学生必读(低年级版)(2019年5期)2019-08-30 03:22:32
中学生数理化·八年级物理人教版(2017年3期)2017-11-09 03:05:33
新课程·下旬(2017年7期)2017-08-14 11:12:50
诗林(2016年5期)2016-10-25 05:19:23
西北林学院学报(2015年5期)2015-01-03 05:12:20
上海金属(2014年5期)2014-12-20 07:58:39
小学生作文·小学低年级适用(2014年2期)2014-09-18 15:45:48
食品工业科技(2014年23期)2014-03-11 18:19:01
